
Abstract
Background:
Glaucoma, the number one cause of irreversible blindness, is characterized by the loss of retinal ganglion cells (RGCs), which do not regenerate in humans or mammals after cell death. Cell transplantation provides an opportunity to restore vision in glaucoma, or other optic neuropathies. Since transplanting primary RGCs from deceased donor tissues may not be feasible, stem cell-derived RGCs could provide a plausible alternative source of donor cells for transplant.
Objective:
We define a robust chemically defined protocol to differentiate human embryonic stem cells (hESCs) into RGC-like neurons.
Methods:
Human embryonic stem cell lines (H7-A81 and H9) and induced pluripotent stem cell (iPSC) were used for RGC differentiation. RGC immaturity was measured by calcium imaging against muscimol. Cell markers were detected by immunofluorescence staining and qRT-PCR. RGC-like cells were intravitreally injected to rat eye, and co-stained with RBPMS and human nuclei markers. All experiments were conducted at least three times independently. Data were analyzed by ANOVA with Tukey’s test with P value of <0.05 considered statistically significant.
Results:
We detected retinal progenitor markers Rx and Pax6 after 15 days of differentiation, and the expression of markers for RGC-specific differentiation (Brn3a and Brn3b), maturation (synaptophysin) and neurite growth (β-III-Tubulin) after an additional 15 days. We further examined the physiologic differentiation of these hESC-derived RGC-like progeny to those differentiated in vitro from primary rodent retinal progenitor cells (RPCs) with calcium imaging, and found that both populations demonstrate the immature RGC-like response to muscimol, a GABAA receptor agonist. By one week after transplant to the adult rat eye by intravitreal injection, the human RGC-like cells successfully migrated into the ganglion cell layer.
Conclusions:
Our protocol provides a novel, short, and cost-effective approach for RGC differentiation from hESCs, and may broaden the scope for cell replacement therapy in RGC-related optic neuropathies such as glaucoma.
Introduction
Glaucoma is one of the leading causes of blindness and is estimated to afflict over 76 million people worldwide by 2020 (Tham et al., 2014). Loss or damage of retinal ganglion cells (RGCs) is a major hallmark of glaucoma, which is defined as a group of chronic neurodegenerative diseases resulting in progressive degeneration of the optic nerve (Agostinone & Di Polo, 2015). Although current treatment such as lowering the intraocular pressure may slow the progression of glaucoma, RGC death and the associated vision loss remains irreversible (Much et al., 2008; Shields, 2008). Thus, there is an unmet need to develop cell-based replacement therapy to restore advanced vision loss associated with RGC death in glaucoma and other opticneuropathies.
While primary donor RGCs may be limited in supply, RGC-like cells differentiated from human embryonic stem cells (hESCs) or human induced pluripotent stem cells (hiPSCs) may provide a source for cell transplantation. In the past decade, many stem cell (SC)-to-RGC differentiation protocols have been published but most of them require growth factors to promote RGC fate specification (Gill et al., 2014). However, cost and batch-to-batch variability of growth factors is a barrier to SC-to-RGC differentiation. Moreover, as large-scale SC-to-RGC differentiation will be required for cell replacement therapies and disease modeling, use of chemically defined medium that minimizes dependence on protein growth factors may offer advantages. Here we studied a chemically defined medium for hESC differentiation into RGC-like progeny, and examined differentiation by marker expression and physiology, as well as integration into the retina after transplant in mouse eyes in vivo.
Material and methods
hESC culture
Human embryonic stem cell lines H9 and H7-A81 (Sluch et al., 2015), and hiPSCs were cultured in mTeSR1 (StemCell Technologies) on 6-well tissue culture plates coated with Matrigel (Corning) or Synthemax (Corning), incubated in 37°C at 5% CO2, fed daily, and manually passaged approximately every 5 days or until about 80% confluence with ethylenediaminetetraacetic acid solution (EDTA, Sigma-Aldrich). For differentiation, hESCs were treated with Dulbecco’s phosphate-buffered saline (DPBS, Thermo Fisher Scientific) without Ca2+ and Mg2+ for 4 minutes and manually passaged and filtered using a 0.22 μm Steriflip Filter for detachment and embryonic body (EB) formation.
hESC differentiation
This chemically defined protocol for neural differentiation was modified from a previously described protocol (Lamba et al., 2006). Once stem cell cultures in 6-well plates were 80% confluent, media was changed to DMEM/F-12 medium supplemented with 1% N2 (both Thermo Fisher Scientific) on day 0. On day 1, medium was supplemented with ALK 2/3 inhibitor dorsomorphin (1 μM, Tocris) and ALK 5 inhibitor SB-431542 (10 μM, Sigma-Aldrich) to block BMP signaling, and Wnt/β-catenin signaling inhibitor IWP-2 (2 μM, Sigma-Aldrich), which we named N2+Dor+SB+IWP2 (N2DSI). On Day 2, to create embryonic bodies (EBs), we washed cells with PBS (without Ca2+/Mg2+) for 5 min at 37°C. Cells were then mechanically detached with cell scraper (BD Falcon) into N2DSI medium and transferred to ultra-low binding plates (Corning). From day 3, N2DSI medium was changed every other day, using a strainer to avoid the loss of EBs, through day 9. On day 10, EBs were dissociated with Accutase (Innovative Cell Technologies) and cultured on Matrigel-coated plates using DMEM/F-12 supplemented with 1% PS, 0.5% N2, 1% B27 and bFGF, 20 ng ml–1 (NBF medium). RPCs formed from EBs over the next 6 days were then dissociated with Accutase and cultured on PDL/laminin-coated plates in NBF medium. Subsequent passaging of RPCs was noted to enrich these cultures; Rock inhibitor Y-27632 (10 μM; EMD Millipore, Billerica, MA) was added to prevent cell death after single cell dissociation.
For RGC differentiation, we unitized previously published Full Sato (FS) medium (Apara et al., 2017; Galvao et al., 2018) containing Notch signaling γ-Secretase inhibitor DAPT (10 μM; EMD Millipore, Billerica, MA). Half volumes of FS+DAPT medium were exchanged every other day until cell harvest.
Animals
All animal studies followed the Association for Research in Vision and Ophthalmology (ARVO) statement for the use of animals in research, and were approved by the Institutional Animal Care and Use Committee (IACUC) and the Institutional Biosafety Committee of the University of California, San Diego. Three-month-old Sprague-Dawley rats, both male and female, and 14-day-timed pregnant C57BL/6 mice were obtained from Charles River (Portage, MI).
Retinal progenitor cell isolation and differentiation
Retinal progenitor cells (RPCs) were isolated from embryonic day 14 (E14) mice, as previously described (Chang et al., 2017). Briefly, retinas were dissected, dissociated with papain (Worthington) in Dulbecco’s PBS (Life Technologies) incubated at 37°C for 30 min, and then gently triturated into single-cell (RPC) suspensions with ovomucoid inhibitor (Roche). RPCs were cultured in DMEM/F12 medium containing glucose (0.6%, Sigma), GS21 supplement (1:100, GlobalStem), Sato supplement (1:100), insulin (5 μg ml–1, Sigma), epidermal growth factor (EGF; 20 ng ml–1, Peprotech), fibroblast growth factor (FGF; 20 ng ml–1, Peprotech), and penicillin/streptomycin (1%, ThermoFisher Scientific), in a humidified atmosphere at 5% CO2, 37°C. RPCs were cultured 7 days before calcium imaging.
Quantitative real-time PCR
Total RNA was isolated from hESC-derived RGC-like cells (RNeasy; Qiagen, Valencia, CA) and treated with DNase before reverse transcription (RT). Then equal amount of RNA was reverse transcribed to cDNA (iScript cDNA Synthesis Kit, Bio-Rad, Hercules, CA). Quantitative real time PCR (qRT-PCR) was performed using primers against GAPDH, Ki67, Nanog, Rx, Pax6, Brn3a, Thy1 and Recoverin (Taqman Gene expression Mix Kit; ThermoFisher Scientific) in the iQ7 Multicolor Real-Time PCR Detection System (Bio-Rad). Gene expression levels were normalized to reference gene GAPDH.
Calcium imaging
For intracellular calcium recordings, E14 RPCs-derived or hESC-derived RGC-like cells were cultured on PDL/laminin-coated glass coverslips as previously described (Trakhtenberg et al., 2017). Coverslips with cultured cells were incubated for 30 min at room temperature with 5 μM fura 2-AM (F-1221; Life Technologies) in Tyrode’s solution: (in mM) NaCl 129, KCl 5, CaCl2 2, MgCl2 1, glucose 30, and HEPES 25, pH 7.4, containing 1 mg/ml bovine serum albumin. The cells were washed with Tyrode’s solution and placed in the imaging chamber (RC-26GLP, Warner Instruments). Calcium imaging was performed on an inverted fluorescence microscope (AxioObserver, Zeiss) with a fast switching light source (DG-4, Sutter Instruments) using MetaFluor software (Molecular Devices) working within the linear range of the camera. Regions of interest (ROIs) were drawn around brightly labeled cell bodies corresponding to cells with neuronal morphology observed under DIC illumination. Calcium responses within these regions of interest were observed as changes in fluorescence ratio (340/380) upon sequential stimulation with muscimol (100 μM, Worthington) and KCl (30 mM) in Tyrode’s solution. Test solutions were washed out with Tyrode’s solution following each stimulus. A response was considered positive if the change in fluorescence ratio exceeded six times the standard deviation in baseline values. With this criterion, cells were scored as responsive or non-responsive to a specific stimulus. More than 25 cells were tested and analyzed in each experiment.
Stem cell derived Retinal ganglion-like cell transplantation
hESC-differentiated RGC-like cells were intravitreally transplanted into adult rats, following our previous protocol (Hertz et al., 2014; Venugopalan et al., 2016). Briefly, cells were centrifuged at 80 rcf for 10 minutes and resuspended in 100 μl of FS medium. 4 μl of cell suspension (~50,000 cells) or vehicle (medium) was injected into the vitreous of anaesthetized rats using a 33-gauge Hamilton syringe. The rats were sacrificed 7 days after transplantation.
Immunostaining
In vitro-differentiated cells were fixed in 2% paraformaldehyde (PFA) for 20 minutes and then permeabilized with 0.1% Triton X-100 (Sigma) for 10 min, blocked with 10% normal goat serum (NGS; Sigma) in PBS for 1 h, and incubated with mouse monoclonal primary antibody anti-Brn3b (1:300; Santa Cruz) or rabbit polyclonal antibodies anti-Nestin (1:600; Cell signaling technology), anti-Ki67 (1:500; Abcam), anti-Pax6 (1:400; Covance), anti-Rx (1:200; Abcam), anti-synaptophysin (1:500; Sysy), anti-Nanog (1:500; Cell signaling technology), anti-Tra-1-60 (1:500; Cell signaling technology), anti-β-III-Tubulin (1:200; Abcam) overnight at 4°C. The cells were rinsed with PBS and incubated with fluorophore-conjugated secondary antibodies overnight, and then analyzed by inverted microscopy imaging.
After in vivo transplantation, rats were sacrificed and perfused with PBS followed by 4% PFA. Eyes were dissected and fixed in 4% PFA for 3 hours, then dissected and flat-mounted on glass slides. The flat-mounted samples were permeabilized with 0.3% Triton X-100 (Sigma) for 20 min, blocked with 10% normal goat serum (NGS; Sigma) in PBS for 1 h, and then incubated with a mouse monoclonal anti-human nucleus primary antibody (1:200; Millipore), RBPMS (1:500; PhosphoSolution) and β-III-Tubulin (1:200; Abcam) overnight at 4°C. Retina samples were rinsed three times with PBS and incubated with Alexa-Fluor 488-conjugated secondary antibody (1:500; Life technologies) overnight. The explants were then rinsed twice, stained with DAPI (1:5000 in PBS) for 15 min, rinsed twice again, and sealed under 0.17 mm coverslips with anti-fade mounting medium (ProLong Gold, Life Technologies) before confocal microscopy imaging.
Statistical analyses
For qRT-PCR, one-way ANOVA and post-hoc t-test with Tukey correction were used to analyze gene expression differences between stem cells and differentiated progeny. P < 0.05 was considered statistically significant. Means and standard errors of the mean (SEM) are shown in all graphs with at least 3 biological replicates per data point.
Results
Differentiation from hESCs to RPCs
Following our chemically defined protocol (Fig. 1A), we first asked if RPCs could be differentiated from hESCs (H7-A81) that expressed stem cell markers including Tra-1-60 and Nanog (Fig. 1B). After differentiation for 15 days, the cells changed their morphology from growth in tight clusters to spread out single cells, and expressed markers Rx, Nestin and Pax6, and also a proliferation maker, Ki67 (Fig. 1C, D), consistent with RPC differentiation [7]. We quantified this differentiation with single- and double-label markers, and found at 15 days in retinal progenitor induction media, 61% of progeny were Ki67+/Rx+, and 83% were Nestin+/Pax6+ in H7-A81 (Fig. 1E); 52% of progeny were Ki67+/Rx+, and 81% were Nestin+/Pax6+ in H9 (Fig. 1F) and 62% of progeny were Ki67+/Rx+, and 90% were Nestin+/Pax6+ in iPSC (Fig. 1G). Thus this chemically defined approach promotes differentiation of hESCs into RPCs.
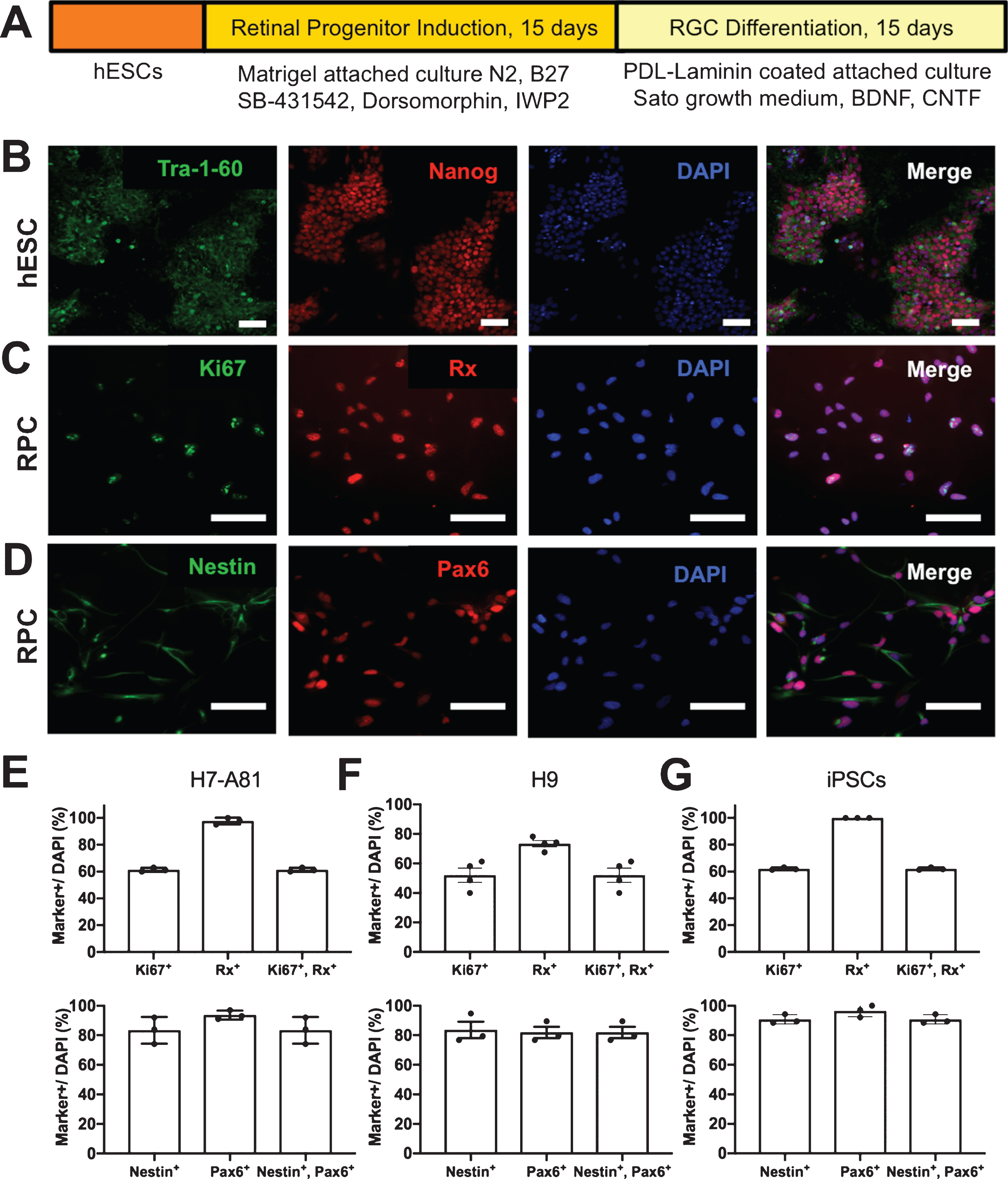
Chemically defined protocol for hESC-to-RPC-to-RGC differentiation. (A) hESCs were differentiated into RGCs using a retinal progenitor induction protocol for 15 days and then an RGC differentiation protocol for another 15 days. (B) hESCs express stem cell markers, Tra-1-60 and Nanog. (C, D) hESC-derived progeny express RPC markers such as Rx, Nestin and Pax6 as well as cell proliferation marker, Ki67. All cells were counterstained with DAPI for nucleus identification. (E-G) Quantification of the fraction of cell progeny expressing RPC genes at 15 days from H7-A81 (E), H9 (F) and iPSCs (G), as marked. Scale bars = 200 μm.
Next, we asked whether hESC-derived RPCs were able to differentiate into functional RGC-like cells. After culturing RPCs for 15 days with RGC culture medium supplemented with BDNF, CNTF, and DAPT, the cells became morphologically differentiated with extensive neurite growth, and expressed neuronal markers Brn3b, and a neurite marker, β-III-Tubulin (Fig. 2A), as well as a synapse marker, synaptophysin (Fig. 2B). After 15 days in this differentiation medium, 9% of progeny were Brn3b+/β-III-Tubulin+. Thus withdrawal of RPC-promoting media and additional of media that supports differentiation and survival of RGC progeny promotes RGC differentiation as assessed by multiple markers.

Differentiation of hESCs to RGC-like cells. (A-C), hESC-derived RPCs were further differentiated to RGC-like cells and express neuron markers such as β-III-Tubulin and Brn3b (A), or β-III-Tubulin and synapse marker, synaptophysin (B). (C) qRT-PCR was performed to detect markers of proliferation (Ki67), stem cells (Nanog), RPCs (Rx and Pax6), RGC-like cells (Brn3a and Thy1) and photoreceptor precursor cells (Recoverin). N = 3, *P < 0.05, One-way ANOVA post-hoc t- test with Tukey correction. Scale bars = 50 μm (A), 100 μm (B).
We further investigated the expression patterns of genes associated with changing differentiation stages by qRT-PCR (Fig. 2C). First, the cell proliferation marker, Ki67, was significantly reduced in the transition from RPC to RGC. Second, the stem cell marker, Nanog, was undetectable after the hESC stage in either RPCs or RGCs. The RPC marker, Rx, was expressed in RPCs but was not detected in hESCs and was significantly downregulated upon RGC differentiation. Finally, increasing gene expression of Pax6, Brn3a and Thy1 was observed as hESCs progressed through RPC and particularly into RGC differentiation stages, consistent with the importance of Pax6 in particular on RGC differentiation in vivo (Kayama et al., 2010). In addition, Ki67 as a marker of proliferating cells went down by half, and we did not detect any Recoverin expression at the RGC differentiation stage, suggesting that photoreceptors were not differentiated in this protocol, or at least not at this time point. For the purposes of this paper we did not undertake further exploration of what cell progeny other than progenitor cells and RGCs might be generated in these cultures. Thus, qRT-PCR confirms quantitatively the coordinated transitions from hESC to RPC to RGC differentiation in this culture model.
Going beyond molecular markers, we also studied the electrophysiological differentiation of these stem cell-derived RGCs, compared with primary RGCs. Newly differentiated neurons undergo several morphological and physiological changes as they mature functionally through development. One such developmental process involves a shift from excitatory depolarization to inhibitory hyperpolarization in response to the neurotransmitter gamma-aminobutyric acid (GABA). For example, early postnatal RGCs depolarize whereas mature RGCs hyperpolarize in response to GABAA receptor activation; only the immature RGCs thus demonstrate a calcium influx in response to GABAA-specific agonists like muscimol. To characterize this measure of electrophysiologic maturity, Ca2+ levels were monitored using ratiometric Fura-2 imaging in response to muscimol in RGCs differentiated from E14 mouse RPCs, in which we observed a muscimol-evoked increase in [Ca2+]i similar to that seen in KCl-induced depolarization (12 of 16 cells; Fig. 3A). Similarly, muscimol stimulated a depolarization-induced increase in [Ca2+]i in hESC-derived RGCs (10 of 12 cells; Fig. 3B). Thus after differentiation in vitro at this time point, hESC-derived RGC-like progeny are physiologically immature. For our hESC RGCs, we could specifically image Brn3-mCherry+ cells. However for the E14 mouse retinal progenitor-derived RGCs, we did not have that option. It is possible that some of the “GABA immature cells” are other non-RGC neurons, but given that the majority of the progeny were thought to be RGC-like, this likely does not affect themain conclusion.

Calcium imaging of rodent RPC-derived and hESC-derived RGCs. (A-B) Cultures of E14 mouse RPC-derived RGCs (A) or hESC-derived RGC (B) were loaded in culture with calcium indicator Fura-2, and then treated with muscimol and KCl as indicated in the example traces. Calcium-evoked response was monitored in real time selecting large nuclei and mCherry-positive RGC progeny in the RPC-derived and hESC-derived cultures, respectively. Scale bars = 20 μm (A), 40 μm (B).
Finally, we asked if the hESC-derived RGC-like cells survive in vivo and migrate into the ganglion cell layer (GCL) after intravitreal transplant in adult rat eyes. Seven days after injection, a significant number of transplanted hESC-derived RGCs were localized in the GCL, as identified by expression of a human nuclear marker that was not detectable in control medium-injected eyes (Fig. 4A-C). Although in these experiments the RGC marker RNA-binding protein with multiple splicing (RBPMS) was not detectable in donor human cells, a subset of donor cells that remained on the retinal surface or in the vitreous clearly expressed β-III-Tubulin (Fig. 4C). In terms of quantifying efficacy, we attempted the transplants into 5 eyes and found cells were detectable in all 5 attempts. Although some areas of the retina naturally had no cells detectable, in retinal sections where donor cells were found, the flatmount density ranged from 19 to 25 cells/mm2 (Fig. 4D), and the cross section donor cells ranged from 3.3 to 25 donor cells per linear mm of retinal section. Thus at least in these initial studies, hESC-derived RGC-like cells survive and migrate into the GCL after intravitreal transplant into the normal adult rodent eye.

Differentiated RGC-like cells survive in the adult retina after transplant. Vehicle (O.S. Control) or hESC-derived progeny (O.D. Transplant) were intravitreally injected into 6-week-old rat eyes. One week after transplant, rats were sacrificed and cryosections (A-C) and flatmount retinas (D-E) were probed with human nuclei, RBPMS and β-III-Tubulin, as marked. White arrows indicate ganglion cell layer (GCL). Scale bars = 200 μm (A-C), 50 μm (D-E).
In this study, we present data on a rapid, chemically defined approach to differentiate hESCs into RGC-like cells. Many stem cell-RGC differentiation protocols have been established (Kobayashi et al., 2018; Langer et al., 2018; Sluch et al., 2017), but the one presented here may offer advantages when considered against these prior pioneering studies. This protocol requires only 4 weeks, and the substitution in the hESC-to-RPC stage of chemical BMP and WNT pathway inhibitors instead of additional peptide growth factors may attenuate costs and reduce batch-to-batch variability. In another prior publication (Sluch et al., 2017), RGC differentiation was promoted with Dorsomorphin (a combined BMP, ALK2, ALK3, and ALK6 inhibitor), IDE2 (to activate Smad2 phosphorylation), nicotinamide, forskolin and DAPT (Notch inhibitor). Our protocol differed through the use of SB431542 (a Smad2 inhibitor) and IWP2 (a Wnt inhibitor), and that forskolin and DAPT were added only in the second differentiation step. Interestingly, the prior paper found that SB431542 reduced RGC differentiation; the pro-differentiation effect of SB431542 in our data may be due to the above differences between the protocols (media additives, time, and/or our use of two steps). Thus this fast, consistent and low-cost protocol provides a potential strategy to generate RGC-like cells for cell replacement in RGC and optic nerve degenerative diseases. In this protocol, we tried three cell lines in the lab, H7-A81, H9 and hiPSCs. All showed similar efficacy and PRC biomarker expression. Since H7-A81 was engineered with CRISPR to express mCherry under BRN3B promoter, we first utilized H7-A81 to further confirm RPC-to-RGC-like cell differentiation. Validation of this protocol in the other human stem cell lines for RGC-like cell differentiation would be an important futuredirection.
Since the RGC morphology closely resembles that of other central nervous system (CNS) neurons, it is difficult to distinguish them from other neurons in vitro. Although immunofluorescence staining and qRT-PCR confirmed the cell marker expression at different stages, many forebrain neurons share with RGCs similar expression of cell markers including Brn3a (Latchman, 1998) and β-III-Tubulin (von Bohlen Und Halbach, 2007). Consequently, a combination of morphological, electrophysiological and other specific markers may be used to characterize hESC-derived RGC-like cells. Using our protocol, differentiated RGC-like cells expressed appropriate morphological features, extending neurites and expressing synaptic proteins in vitro. Interestingly, however, calcium imaging of GABAA receptor responses revealed electrophysiologic immaturity of these RGC progeny. Future experiments exploring the signaling needed to mature these cells’ physiologic properties will help identify the pathways important in this developmental process, as well as allow testing of whether the stage of electrophysiologic differentiation impacts integration in vivo.
Additionally, we intravitreally injected RGC-like cells into adult rat eyes but only few RGC-like cells were observed in the GCL. This limitation may be attributable to the low yield of RGC-like cells differentiated from stem cells, as we did not attempt to purify RGCs from among the hESC-derived progeny before transplant. Previous studies successfully increased the yield of stem cell-derived RGC-like cells by Thy1 immunopanning (Gill et al., 2016; Teotia et al., 2017). We tried to purify RGC-like cells using mouse Thy-1 immunopanning but couldn’t get enough cells to do transplantation. Therefore, the whole, differentiated cell population was transplanted. Injection of more purified RGC-like cells will be a future direction to strengthen our results. RBPMS marker expresses later than β-III-Tubulin during RGC development. In transplant experiments, we did not see RBPMS expression in human donor cell, suggesting that transplanted cells may not reach the threshold of differentiation as fully mature RGCs. We previously found that co-injection of stem cells with RGCs promoted RGC survival (Wu et al., 2018), but a question remaining for the future is whether co-injection of non-RGC progeny affects the survival and integration of the RGCs after transplant. Incomplete differentiation, maturation or integration after transplant could also be attributable to incomplete expression of transcription factors normally required for RGC fate specification. For example, SoxC family genes play important roles in RGC fate specification and promote stem cell-to-RGC differentiation (Chang et al., 2017; Kuwajima et al., 2017). Exogenous expression of SoxC genes may be a strategy to generate more RGC-like cells and could in parallel benefit integration (Chang & Hertz, 2017). Extrinsically, growth and differentiation factors (GDFs) regulate RGC differentiation (Chang et al., 2019). Here we used human ESCs for RGC differentiation, but in our prior publications we found when differentiating mouse or human ESCs to RGCs, the signaling pathway cascades for differentiation are similar. Studies examining migration into the adult retina following transplant suggest RGCs integrate into the GCL and extend their dendrites into the inner plexiform layer and their axons along the nerve fiber layer (Hertz et al., 2014; Venugopalan et al., 2016). Nevertheless, longer-term experiments will be necessary to assay axon growth, synapse formation and functional aspects of transplanted RGC-like cells. In addition, this study was done in the normal eye to demonstrate that human ESC-differentiated RGC-like cells can integrate to healthy host retina. Transplantation into injured retina is an important question for future study. Given the importance of RGC transplantation as a potential treatment for glaucoma and related optic neuropathies, improved cell survival and successful integration in retinal circuitry are still critical challenges that need further research.
Conflict of interest
The authors declare no conflict of interest.
Footnotes
Acknowledgments
This study was supported by the BrightFocus Foundation, the National Eye Institute (P30-EY026877 and F32-EY029137) and Research to Prevent Blindness, Inc.