
Abstract
Background:
To study motion perception, a stimulus consisting of a field of small, moving dots is often used. Generally, some of the dots coherently move in the same direction (signal) while the rest move randomly (noise). A percept of global coherent motion (CM) results when many different local motion signals are combined. CM computation is a complex process that requires the integrity of the middle-temporal area (MT/V5) and there is evidence that increasing the number of dots presented in the stimulus makes such computation more efficient.
Objective:
In this study, we explored whether anodal direct current stimulation (tDCS) over MT/V5 would increase individual performance in a CM task at a low signal-to-noise ratio (SNR, i.e. low percentage of coherent dots) and with a target consisting of a large number of moving dots (high dot numerosity, e.g. >250 dots) with respect to low dot numerosity (<60 dots), indicating that tDCS favour the integration of local motion signal into a single global percept (global motion).
Method:
Participants were asked to perform a CM detection task (two-interval forced-choice, 2IFC) while they received anodal, cathodal, or sham stimulation on three different days.
Results:
Our findings showed no effect of cathodal tDCS with respect to the sham condition. Instead, anodal tDCS improves performance, but mostly when dot numerosity is high (>400 dots) to promote efficient global motion processing.
Conclusions:
The present study suggests that tDCS may be used under appropriate stimulus conditions (low SNR and high dot numerosity) to boost the global motion processing efficiency, and may be useful to empower clinical protocols to treat visual deficits.
Introduction
When looking at a cloud of dots in correlated motion, we tend to perceive coherent rigid motion rather than a multitude of point motions. This percept reflects a fundamental aspect of visual processing, the ability to combine coherent local motion signals into a global directional motion percept (Smith, Snowden, & Milne, 1994).
In literature, the ability to combine local elements in motion is measured using the coherent motion paradigm (CM). This paradigm consists of the presentation of two moving dot populations within the stimulus area, one with a certain number of dots moving in the same direction (signal) and the other made up of dots that move in random directions (noise). The observer is asked to identify the direction of the signal dots, a task that requires spatial integration of coherently moving signals (Smith et al., 1994). An important characteristic of this specific stimulus is that the signal-to-noise ratio (SNR), i.e. the ratio between the number of signals (coherent) and noise (random direction) dots, of each presentation is adjustable from trial to trial. Thus, it is possible to psychophysically measure a motion coherence threshold for a motion detection task or a motion discrimination task (Newsome & Pare, 1988).
Physiological studies generally acknowledge that neurons in the middle-temporal area (MT) mediate the cortical representation of CM. These neurons signal global coherent motion by integrating a population of motion directions (Britten, Shadlen, Newsome, & Movshon, 1993). The importance of the MT area in computing coherent motion was confirmed in recent decades using several experimental approaches: psychophysics, lesions, brain-stimulation, and neuroimaging (Alais & Blake, 1999; Heeger, Huk, Geisler, & Albrecht, 2000; Marcar, Zihl, & Cowey, 1997). For instance, studies have shown that micro-stimulation on monkeys or transcranial electrical stimulation on humans, a procedure used to modulate cortical excitability, allows us to discern between different functional hypotheses by looking at behavioral changes in subjects’ performance (Antal, Nitsche, et al., 2004; Battaglini et al., 2019; Battaglini, Contemori, Penzo, & Maniglia, 2020; Battaglini, Ghiani, Casco, & Ronconi, 2020; Battaglini, Mena, et al., 2020; Camilleri, Pavan, Ghin, Battaglini, & Campana, 2014; Pavan et al., 2019; van der Groen, Tang, Wenderoth, & Mattingley, 2018; Zito et al., 2015). For example, in a previous study we used anodal and cathodal direct current stimulation (tDCS) to induce changes in cortical activity over MT with the aim to investigate the effect of electrical stimulation in a CM direction discrimination task (Battaglini et al., 2017). We found that both anodal (a-tDCS) and cathodal stimulation (c-tDCS) can improve performance, though this was achieved through different mechanisms. Specifically, a-tDCS enhances the strength of the signal dots (improving selectivity for target motion direction), whereas c-tDCS reduced the influences of the noisy dots (direction far from the target). In both cases, at a neural level, the SNR increases (Battaglini et al., 2017). Both polarities (anodal and cathodal tDCS) may operate by increasing the precision of the neural representations of each element direction (Barlow & Tripathy, 1997; Dakin, Mareschal, & Bex, 2005; Eagle & Rogers, 1997) and both mechanisms contribute to the final goal of reducing unreliable local processing, thereby increasing the capability of selecting a restricted number of directions for integration (Barlow & Tripathy, 1997; Dakin et al., 2005; Eagle & Rogers, 1997).
In the study conducted by Battaglini et al. (2017), the effect of a-tDCS at low SNR was weak. This was unexpected since a number of vision studies showed that a-tDCS boosts weak signals (Antal, Nitsche, & Paulus, 2001; Reinhart, Xiao, McClenahan, & Woodman, 2016). Given this property of a-tDCS, we expected a-tDCS, particularly at low SNR, to boost the capability to pool together motion directions relevant to extract global motion. Pooling is well-established as an operation essential to the computation of global motion (Dakin et al., 2005); moreover, this operation may be boosted by brain stimulation (Pavan et al., 2019). In the present study, we hope to disentangle the role of the different polarity of tDCS in boosting either the local or global computation involved in coherent motion perception. We did this by manipulating the number of dots in the stimulus area. Many studies have analysed how dot density affects the segregation of signal from noise directions. Most of these studies used the random dot kinematogram as a stimulus (RDK) and assessed how dot density affected the capability to segment target signals from noise. In RDK, global motion perception is limited by the false correspondence between dots in successive frames (Barlow & Tripathy, 1997; Eagle & Rogers, 1997). Barlow and Tripathy (1997) showed near-invariance of the coherent motion threshold with dot density. Eagle and Rogers (1997) found that the maximum spatial displacement at which observers were able to confidently discriminate the direction of motion (Dmax) increased as dot density decreased. Different results were obtained when the global motion stimulus was more appropriate to activate a pooling mechanism (Dakin et al., 2005). The most appropriate stimulus is the one used in the equivalent noise paradigm, in which noise is manipulated by varying the standard deviation of the distribution of motion directions presented, which forces the visual system to integrate all directions of motion. This paradigm has been used to assess the role of different factors in local and global computation of coherent motion: attention (Ling, Liu, & Carrasco, 2009), perceptual learning (Cavanaugh et al., 2015; Gold, Bennett, & Sekuler, 1999), and transcranial random noise stimulation (Pavan et al., 2019). The other stimulus, widely used to investigate motion perception, is CM. CM is similar to RDK but differs in that thresholds do not refer to a local limit (Dmax), but rather to a global limit (i.e., the minimum percentage of signal elements that allowed to perceive CM). With both of these stimuli, the coherent motion threshold decreases as dot density increases. In particular, according to Dakin et al. (2005), the precision of the observer’s representation of each element direction (local noise) as well as how many of these estimates are averaged (i.e., global pooling, global sampling) are limited by the number of dots (not dot density). Specifically, local noise dependency on the number of dots saturates at a relatively low number of dots (∼ 60 dots), whereas, for a larger number of dots, global sampling/pooling increases with the number of dots. Moreover, when using a CM stimulus, Dakin et al. (2005) found that global motion computation also improved with the number of dots (they tested stimuli consisting of 16, 64 and 256 dots), although the number of directions that are integrated is limited.
Based on these premises, if a-tDCS boosts weak signals (Antal et al., 2001; Reinhart et al., 2016) and boosts the capability to pool together motion directions relevant to extract global motion, we would expect changes in performance at low SNR (coherence around 20–30%) and at high dot numerosity (>250 dots). This would indicate that tDCS affects global pooling, that is the number of estimates of local motion that can be averaged. First, we measured the modulation of detection performance by tDCS when detection performance was at subthreshold level, that is, when accuracy, in a 2AFC task, was just above chance level. Second, we used a detection task, because unlike discrimination, it relies (mostly) on the neuronal population having a preferred direction close to that which is meant to be detected (Hol & Treue, 2001; Regan & Beverley, 1983). This way, we can (theoretically) maximize the effect of a-tDCS. Third, we aimed to assess the role of dot numerosity. In particular, we asked whether baseline performance (at subthreshold level) is varied by tDCS in a way that depends on the number of dots present in a given stimulus area.
Method
Participants
Twenty-eight healthy students (11 males, mean age = 23.72, age range = 19–27 years) with normal or corrected-to-normal vision were recruited from the University of Padova. All of them completed all the experimental sessions. Exclusion criteria included a history of psychiatric disorders, drug abuse, active medication, pregnancy, and susceptibility to seizures. Participation in the study was voluntary. Individuals were naïve to the experimental procedure as well as its purpose. Informed consent was obtained from all participants, and the study was conducted in accordance with the Declaration of Helsinki (1964). The experimental methods received ethical approval from the University of Padova (protocol number 3058).
Stimuli and apparatus
Participants were placed in a dimly lit room and seated 57 cm away from the display screen. Viewing was binocular. Stimuli were generated with MATLAB Psychtoolbox (Brainard, 1997; Pelli, 1997) and displayed on a 19-inch Asus monitor with a refresh rate of 60 Hz. The screen resolution was set to 1920×1080 pixels, and each pixel subtended ∼1.5 arcmin. The luminance of the background was 0.7 cd/m2. The stimulus was a square window of 10 deg filled with white moving dots, each one with a diameter of 0.075 deg. CM resulted from a temporal sequencing of 8 frames. In the first frame, the dots were randomly positioned within the square window, and then displaced by 0.05 deg in each subsequent frame (i.e., Brownian motion) (Pilly & Seitz, 2009), producing a speed of 3 deg/s. The stimulus was displayed in the centre of the screen for 133 ms (8 frames at a frame duration of 16.67 ms). The dots had a limited lifetime of 66 ms; they were then reallocated to a random position. In addition, moving dots that travelled outside the window were replaced by new dots at different, randomly selected locations within the square window. Different dot numerosities were used in this study (25; 50; 100; 204; 413; 845; 1700 dots/deg2). The direction of the coherently moving dots was rightward; the other dots moved in random directions (Fig. 1).
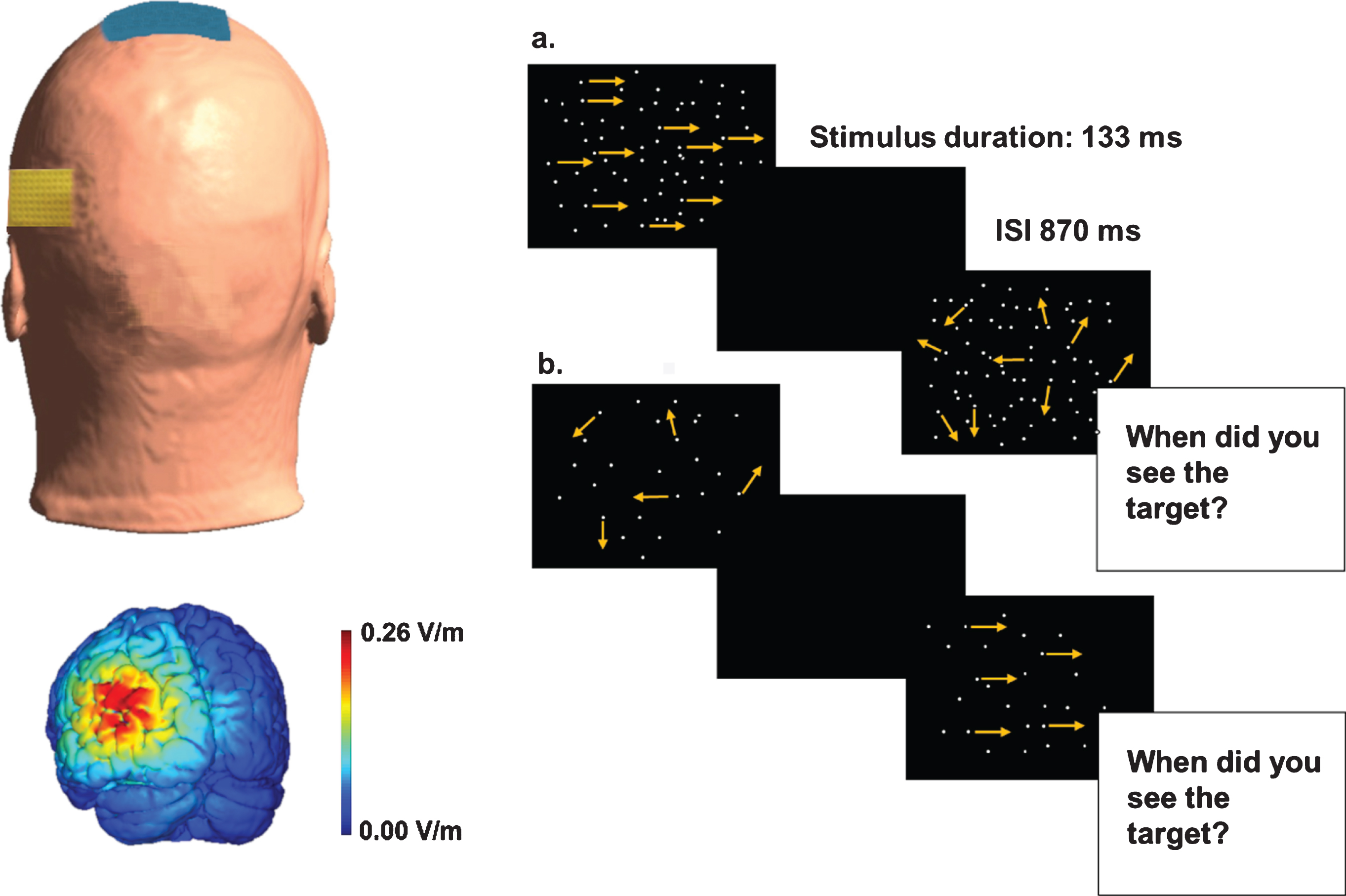
Stimulation Setting and Task. Upper left: the position of the electrodes used during tDCS (for illustration purpose only. The size of the electrodes in relation to the size of the head might not be very precise). The electrode used as reference was placed on Cz (blue sponge), and the active electrode was on MT (yellow sponge). Bottom left: graphical representation of the electrical field strength (V/m) at the moment when the current reaches the maximum anodal intensity. Right side: description of a trial of the CM detection task in case of high dot density (a) or low dot density (b).
A battery-driven constant-current stimulator (BrainStim, EMS) delivered the tDCS through a pair of saline-soaked sponge electrodes. The size of the stimulating electrode (considering the sponge) placed over the left 1 MT was 72×60 mm and the size of the reference electrode (considering the sponge) placed over the vertex (Cz) was 115×95 mm, rendering the large reference electrode inert due to low current density (Fig. 1). The current was initially ramped up over 30 seconds to an intensity of 1.5 mA, and then kept constant. In the sham session, the current was delivered only for the first 30 s and the last 30 s of the 12 min session.
CM detection task
Each trial consisted of the random presentation of two intervals: one interval showed the CM in which a few target dots moved rightwards among a field of randomly moving dots; the other interval showed the randomly moving dots pattern. Participants were asked to perform a CM detection task. They had to indicate which of the two intervals contained the target dots (two-interval forced choice, 2IFC), by pressing, respectively, the “z” or the “m” button on the keyboard. To prevent any (short) adaptation effect (Kanai & Verstraten, 2005), the second interval followed the first after 870 ms. When the second stimulus disappeared, participants made their decision and answered. The following trial started a second after the response was generated. The successive trial did not start until an answer was registered.
Procedure
Participants came three times to the laboratory on three different days. Experiment 1 was run on day 1 in two sessions: in the first, individual CM threshold was calculated, in the second session the percentage of coherent moving dots producing a sub-threshold performance was found. Experiment 2 consisted of three different online (i.e. during the CM task) stimulation sessions, namely anodal tDCS, cathodal tDCS and sham, performed in counterbalanced order between subjects (single-blind design) in three different days, with one day of interval between sessions for washout purposes (Experiment 2 started on day 1 after Experiment 1 ended).
Experiment 1: Threshold assessment and verification
It is well known that CM performance depends on several factors, such as the task that participants must perform and eccentricity (Hol & Treue, 2001; Thompson, Hansen, Hess, & Troje, 2007). To ascertain individual threshold for the experimental conditions of the present study, in the first session of Experiment 1, we used a single one-up and one-down staircase (Levitt, 1971), where the SNR changed according to the participant’s response. The session terminated after either 100 trials or 12 reversals. The staircase tracked the coherence level producing 75% of correct responses. The stimulus consisted of 100 white moving dots. To find a subthreshold SNR, we used, in the second session of Experiment 1, the same stimulus (100 white moving dots) and estimated the percentage of coherent moving dots producing an accuracy between 60% and 70%. Therefore, we reduced the threshold value by 10% and observed the performance of a block of 30 trials with a fixed SNR. If the percentage of correct responses was not between 60–70%, we repeated the block, adding or taking away two dots, until we attained our goal (mean block 2.62, ds 1.14).
Experiment 2: Dot numerosity effect
In the main experiment, the individual SNR was fixed and set to that obtained in the second session of Experiment 1. Unlike Experiment 1, we varied the number of dots (25; 50; 100; 204; 413; 845; 1700 dots/deg2), while maintaining fixed the stimulus area, to assess the effect of the absolute number of dots on the strength of the tDCS effect (see introduction). Participants completed 168 trials, 24 for each level of noise. From the proportion of correct answers, we estimated d’ (sensitivity index 2 ) using Signal Detection Theory (SDT), at each noise level.
Results
First, we assessed whether the accuracy in the sham condition of Experiment 2 depended on the threshold value obtained in the baseline Experiment 1. We fitted a regression line to the individual accuracy data obtained in the sham session and the number of dots required to reach a performance between the 60% and 70% correct detection that was measured in the second session of Experiment 1. The R2 was equal to 0.00754, indicating that there was no relationship between the accuracy in the sham condition and the threshold value obtained in the baseline experiment.
Then, we consider d’ values (Fig. 2) obtained in Experiment 2 (Green & Swets, 1966; Macmillan & Creelman, 2004; Prins, 2016). Before analysing the effect of stimulation, we performed a preliminary analysis to evaluate the effect of task repetition. We ran an ANOVA with Day (day 1, day 2, day 3) and Dot Numerosity as within factors. None of the effects, Day (F (1.575,42.516) = 0.039; p = 0.96; η2p = 0.001), Dot Numerosity (F (6,162) = 1.994; p = 0.07; η2p = 0.07), and the interaction Day×Dot Density (F (12,324) = 0.996; p = 0.45; η2p = 0.036) was significant. According to these results, the number of trials in the three sessions was not sufficient to determine an improvement in performance due to task repetition.
After, we analysed sensitivity (d’) using a repeated measures ANOVA, with Stimulation Condition and Dot Numerosity as within factors. The analysis revealed a significant effect of Stimulation Condition (F (1,27) = 6.74; p = 0.005; η2p = 0.2), whereas Dot Numerosity (F (6,162) = 1.994; p = 0.07; η2p = 0.07) and the interaction between these two factors did not yield significant results (F (6,162) = 1.379; p = 0.17; η2p = 0.05). The t-tests, with Bonferroni correction, showed that d’ values obtained in the a-tDCS session were higher, compared to those obtained in the c-tDCS (pcorr = 0.02) and the sham sessions (pcorr = 0.033). Neither of the post-hoc comparisons showed a significant difference between Dot Numerosity levels.

Graph shows d‘ obtained in the sham, c-tDCS, and a-tDCS conditions. On each boxplot, the bottom and top edges of the box indicate the 25th and 75th percentiles, respectively. The whiskers extend to the most extreme data points. Black lines inside the box represent the medians and black squares represent the means. Asterisks indicate significant difference (p < 0.05).
Contrary to our expectations, we did not find an interaction between Stimulation Condition and Dot Numerosity. However, from visual inspection of Fig. 3, it is possible to argue in favour of our prediction of an effect of Dot Numerosity, as d’ values obtained in the a-tDCS condition is higher than that obtained in the sham condition, in particular at high dot numerosity.
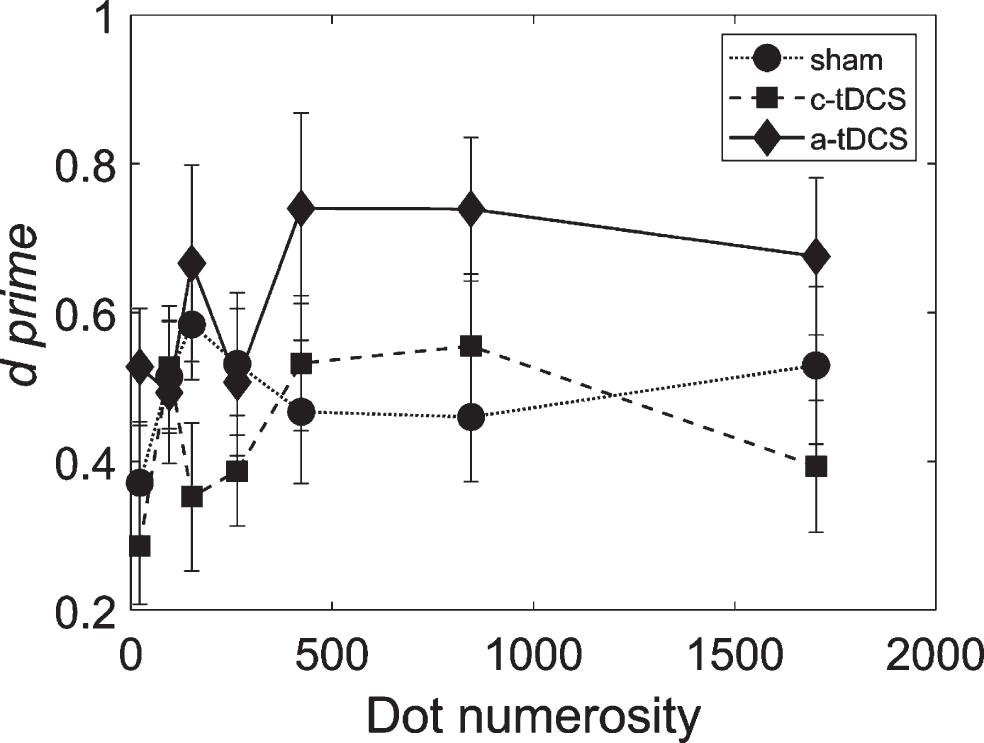
Graph shows d‘ obtained in the sham (circles and dotted line), c-tDCS (squares and broken line) and a-tDCS (diamonds and black line) condition as a function of dot numerosity. Bars represent s.e.m.
To better evaluate the extent to which tDCS modulated baseline performance at the different levels of dot number, we computed individual sensitivity enhancement indices (SE = d’tDCS–d’Sham) for both polarities of stimulation conditions. We used a one-sample t-test to assess, for each dot numerosity, whether SE was higher than, lower than, or equal to zero, indicating increased, reduced, or unaffected sensitivity (d’) enhanced by tDCS. SEs differed from 0 only in the a-tDCS conditions and only at medium-high levels of density. Indeed, the t-test and Benjamin-Hochberg procedure for multiple comparisons performed against the null hypothesis of zero SE had significant results only at medium-higher numerosities (413: pcorr = 0.0099; 845: pcorr = 0.0032).
A widely debated issue in motion perception concerns the way in which the visual system extracts coherent motion from a stimulus consisting of a group of dots that moves coherently against a background of randomly moving dots. There is general consensus that global motion processing relies on the efficiency of three mechanisms: the ability of the visual system to estimate local motion direction, the ability to perform a global integration of these local signals, and the ability to segment coherent motion from random motion. Recent studies have suggested that transcranial electrical stimulation may boost different levels of processing of coherent motion, depending on the type and polarity of stimulation and the level of activation of the visual system, below or at threshold (Battaglini et al., 2017; Pavan et al., 2019). For example, random noise stimulation has been shown to improve global pooling (Pavan et al., 2019). On the other hand, in a previous study (Battaglini et al., 2017), using a motion direction discrimination task, we found that a-tDCS increased the efficiency in estimating local motion direction at medium-high SNR (better performance at [supra]-threshold level of SNR), whereas c-tDCS boosted segregation of coherent motion signals from noise at low SNR (better performance at sub-threshold level of SNR). By investigating the effect of tDCS when performance was at subthreshold level, which is when the task was sufficiently difficult for tDCS to act optimally (Antal et al., 2001; Reinhart et al., 2016), we asked whether c-tDCS and a-tDCS modulated different mechanisms: - segmentation of coherent motion from random motion at low dot numerosity and - global pooling at high dot numerosity (Dakin et al., 2005).
Our results partially support this prediction. Indeed, c-tDCS did not improve detectability of coherent motion, with respect to sham, regardless of dot numerosity; this did not confirm previous results indicating a possible improvement of performance via noise reduction during motion discrimination (Battaglini et al., 2017) or speed discrimination task (Zito et al., 2015). However, it should be noted that the present paradigm differed in several respects from those used in previous studies. First, only one SNR, rather than many levels of subthreshold coherence, was used. Second, a detection task, rather than a discrimination task, was used. Third, observers had to perform only one daily session for each stimulation condition. Finally, in the studies by Battaglini et al. (2017) and Zito et al. (2015), the stimulus presentation was eccentric. For these reasons, further studies are required to determine whether one or more of these factors contributes to the lack of effect of c-tDCS.
The main result of the present study was the effect of a-tDCS at low SNR and high dot numerosity (>400 dots). Under these conditions we found an enhancement in sensitivity (d’) for CM when a-tDCS stimulation was applied, relative to the sham stimulation and c-tDCS. These results better specify the effect of a-tDCS on CM detection at (supra)-threshold level of SNR that we observed in our previous study (Battaglini et al., 2017). There, using a Spearman-Karber method, we found an overall displacement of the psychometric functions, describing how d’ changes as a function of coherence level, but we did not find a change in slope (Ulrich & Miller, 2004). This is indicative of a reduction of internal noise at medium coherence levels, which makes it easier to extract local direction of motion.
The present result illustrates the role of a-tDCS at low SNR. It indicates that a-tDCS is able to improve the sensitivity subthreshold for coherent motion, but probably only when numerosity is sufficiently large to promote an efficient activation of the global pooling mechanism.
Taken together, the results of our previous (Battaglini et al., 2017) and present studies support the hypothesis that a-tDCS may modulate the response of two different mechanisms. At low SNR, it enhances the efficiency of the computation of global direction of motion, particularly when the number of motion signals is appropriate; at medium SNR, our previous results suggest that a-tDCS may reduce the internal neural variability. This reduction of internal noise may improve the ability of the local motion mechanism to estimate the local motion direction of each dot. This double effect of a-tDCS can be fitted within the framework of the equivalent noise paradigm. Indeed, according to the equivalent noise model, if tDCS modified performance at high dot numerosity, this would indicate that tDCS affects global pooling. Conversely, if we had found an effect of tDCS at low dot numerosity, we might have interpreted this to reflect a reduction of internal noise (Dakin et al., 2005).
One qualitative aspect of our results is worth discussing. To clarify why the interaction between Stimulation Condition and Dot Numerosity was not significant, we fitted individual data in the a-tDCS condition with both a linear and a quadratic function. We found that the individual quadratic fit (mean R2 = 0.32, SD = 0.25), was better than the individual linear fit (mean R2 = 0.19, SD = 0.18). That is, an inverted U function seems to be the best description of the relationship between sensitivity and dot numerosity (see supplementary material). An inverted U-shaped function usually describes the stochastic resonance phenomenon, whereby the addition of random noise can enhance either detection of a subthreshold signal or the information that can be extracted from the signal. Stochastic resonance reflects the enhancement of signal detection when an optimal amount of external noise is added to the signal. This optimal amount of external “good-noise” coincides with the peak of the U-shaped function, whereas lower or higher external noise is ineffective. Previous studies have interpreted the effect of brain stimulation on detectability of CM (Pavan et al., 2019; van der Groen et al., 2018). However, the results of the present study may suggest that a “good-noise” effect does not result from an appropriate modulation of SNR but, rather, from having the appropriate number of motion signals involved in the global processing.
There has been speculation that internal noise reduction and increased capacity of global pooling may have different neural sources. Reduction of internal noise may originate from reduced variability of the firing of action potentials and/or in synaptic transmission (Faisal, Selen, & Wolpert, 2008; Manning, Dakin, Tibber, & Pellicano, 2014). Both factors reflect the efficiency of direction-sensitive neurons in the primary visual cortex. Indeed, an increased tuning to the direction of local motion may be related to a narrower bandwidth of neurons in V1; a-tDCS may mediate this process by means of decreasing the activation of GABAergic neurons (Stagg, Bachtiar, & Johansen-Berg, 2011; Stagg & Johansen-Berg, 2013) and thereby induce a shift towards excitation in the cortex. On the other hand, increased efficiency of global pooling may reflect an increased efficiency of neurons at a higher level of motion processing, such as the MT area, operating by integration of local motion signals (Britten, Shadlen, Newsome, & Movshon, 1992). Such a goal may also be related to the potential properties of a-tDCS to reduce the activation of GABAergic neurons or to increase the activation of glutamate neurons (Stagg et al., 2011; Stagg & Johansen-Berg, 2013). On the other hand, c-tDCS may mediate the capacity of segregating signal from noise by means of its potential to selectively activate GABAergic neurons (Stagg et al., 2011; Stagg & Johansen-Berg, 2013) and thereby induce a shift towards inhibition. In sum, the capability of tDCS to perturb the homeostasis of the system in either direction, towards inhibition or towards excitation, may be called into question.
Interesting, the variability in our study was quite large (see Fig. 3) suggesting that not all the participants react to a given stimulation in the same way. A number of studies have investigated factors that may enhance or reduce tDCS effects such as stimulation duration (Monte-Silva et al., 2013), individual baseline performance (Benwell, Learmonth, Miniussi, Harvey, & Thut, 2015; Learmonth, Thut, Benwell, & Harvey, 2015; Wu, Li, Liu, Xu, & Xiao, 2020) and gender (Rudroff, Workman, Fietsam, & Kamholz, 2020). Recently, Wu et al., (2020) showed a positive correlation in a coherent motion direction identification task (4AFC) between the initial and the final threshold performance after 20 minutes of a-tDCS (Wu et al., 2020). Participants with a poorer initial performance had a greater tDCS improvement. We run a correlational analysis between initial performance (the subthreshold values of SNR obtained in the second session of Experiment 1) and the tDCS improvement calculated as the difference between the mean anodal (cathodal) performance and the mean sham performance. In the a-tDCS condition, we observed a weak, but significant correlation (beta = 0.008; R2 = 0.23; p = 0.009), a greater tDCS effect was observed in those participants who had a poor initial performance. This is in line with the result obtained by Wu et al., (2020), and this may be an important factor to take into account in future research.
Studies underline the importance of transcranial current stimulation in rehabilitative training aiming at producing neuroplastic changes (Bolognini, Pascual-Leone, & Fregni, 2009; Campana, Camilleri, Pavan, Veronese, & Lo Giudice, 2014; Peters, Thompson, Merabet, Wu, & Shams, 2013). Although it is difficult to state which components of the electric field (amplitude, phase, polarity, type of stimulation) are most predictive of behavioral outcome, results were promising and worth further investigation. Several researches used transcranial current stimulation in rehabilitation protocol (Alber, Moser, Gall, & Sabel, 2017; Bola et al., 2014; Gall et al., 2016; Sabel et al., 2020), however to our knowledge, only one study combined coherent motion discrimination and tDCS (Halko et al., 2011). In this study, the CM-tDCS protocol had the potential of reducing visual field defects in a patient with hemianopia due to stroke. Our results may provide useful indications for increasing the benefit of the protocols based on CM training. They suggest that before planning a clinical experiment, it should be taken into account that cortical plasticity induced by tDCS is activity-dependent, that is it depends on “the state of excitability of the system” (Fertonani & Miniussi, 2016). Therefore, to maximize the boosting effect of tDCS in rehabilitative protocols using the CM task, it is relevant to understand the mechanisms involved in this task and the way their response is modulated by brain stimulation. Concerning coherent motion task, recent and the present study (Battaglini et al., 2017; Pavan et al., 2019) showed that transcranial current stimulation may boost three different mechanisms involved in this task: the ability of the visual system to estimate local motion direction, the ability to perform a global integration of these local signals, and the ability to segment coherent motion from random motion. Which of these mechanism is modulated by tDCS, depends not only on the SNR but probably also on dot numerosity. The activity dependent model would predict an effect of tDCS when SNR ranges from just below to just above threshold and indeed the three mechanisms are modulated by tDCS when SNR is within this range. Moreover, a stimulation dependent empowerment of the global pooling mechanism would occur when, at subthreshold SNR, many coherent dots have their response close to the threshold value, as occurs when dot numerosity is high. To conclude, the present study showed that tDCS targeting the MT area may be used to boost motion processing efficiency at different levels of the visual system. This result has relevant clinical consequences. Indeed, it has been shown that coherent motion has rehabilitative potential applicable to a range of conditions, such as amblyopia (Hess, Mansouri, Dakin, & Allen, 2006), migraine (Tibber, Kelly, Jansari, Dakin, & Shepherd, 2014), aging (Arena, Hutchinson, Shimozaki, & Long, 2013; Bocheva, Angelova, & Stefanova, 2013), dyslexia (Chouake, Levy, Javitt, & Lavidor, 2012), and hemianopia (Huxlin et al., 2009). Our results may be useful to improve the efficacy of rehabilitation protocols to treat visual deficits and advance our understanding of the neural bases of these rehabilitative effects.
Authors contribution statement
L.B and C.C. designed and implemented the experiments. F.M. collected and preprocessed the data. F.M. and L.B. analyzed the data. All authors interpreted the results, L.B. and C.C. wrote the main manuscript.
Disclosure of funding sources
The study was supported by a grant form MIUR (Dipartimenti di Eccellenza DM 11/05/2017 n.262) to the Department of General Psychology
Declaration of competing interest
The authors declare that they have no conflict of interest related to publication of this manuscript.
Footnotes
Left instead of Right MT was chosen because previous studies have shown that left stimulation produces a greater modulation of the visual motion task than does right stimulation (Accornero, Voti, La Riccia, & Gregori, 2007; Antal, Kincses, Nitsche, Bartfai, & Paulus, 2004; Antal & Paulus, 2008).
d’ is a statistic used in the Signal Detection Theory. It is the standardized difference between the means of Signal Present and Noise (Signal Absent) distributions. Larger absolute values of d’ mean that a participant is more sensitive to the difference between the Signal Present and Noise (Signal Absent) distributions. d’ values near zero indicate chance performance.