
Abstract
Background:
The evaluation of motor imagery in persons with prolonged Disorders of Consciousness (pDOC) is a practical approach to differentiate between patients with Minimally Conscious State (MCS) and Unresponsive Wakefulness Syndrome (UWS) and to identify residual awareness even in individuals with UWS. Investigating the influence of motor observation on motor imagery could be helpful in this regard.
Objective:
In order to corroborate the clinical diagnosis and identify misdiagnosed individuals, we used EEG recordings, to assess the influence of the low-level perceptual and motoric mechanisms on motor observation on motor imagery, taking into account the role of the high-level cognitive mechanisms in patients with pDOC.
Methods:
We assessed the influence of motor observation of walking in first-person or third-person view (by a video provision) on motor imagery of walking in the first-person view on the visual N190 (expression of motor observation processing), the readiness potential (RP) (expressing motor preparation), and the P3 component (high-level cognitive processes) in a sample of 10 persons with MCS, 10 with UWS, and 10 healthy controls (CG). Specifically, the video showed a first-view or third-view walk down the street while the participants were asked to imagine a first-view walking down the street.
Results:
CG showed greater N190 response (low-level sensorimotor processing) in the non-matching than in the matching condition. Conversely, the P3 and RP responses (high-level sensorimotor processing) were greater in the matching than in the non-matching condition. Remarkably, 6 out of 10 patients with MCS showed the preservation of both high- and low-level sensorimotor processing. One UWS patient showed responses similar to those six patients, suggesting a preservation of cognitively-mediated sensorimotor processing despite a detrimental motor preparation process. The remaining patients with MCS did not show diversified EEG responses, suggesting limited cognitive functioning.
Conclusions:
Our study suggests that identifying the low-level visual and high-level motor preparation processes in response to a simple influence of motor observation of motor imagery tasks potentially supports the clinical differential diagnosis of with MCS and UWS. This might help identify UWS patients which were misdiagnosed and who deserve more sophisticated diagnoses.
Keywords
Introduction
The differential diagnosis of patients with prolonged disorders of consciousness (PDOC), including minimally conscious state (MCS) and unresponsive wakefulness syndrome (UWS)(Laureys et al., 2010), relies on behavioral assessment using specific scales, for example, the Coma Recovery Scale— Revised (CRS-R)(Giacino, Kalmar & Whyte, 2004), which aims to assess purposeful motor responses as signs of awareness. However, owing to several biasing factors, such as arousal fluctuations and attentional and sensorimotor deficits (Wade, 2018), advanced paraclinical approaches (neurophysiology and functional neuroimaging) are required to correctly diagnose significant percentage of patients. Specifically, these patients present with a clinical– paraclinical dissociation of awareness, which has been referred to as covert awareness, cognitive– motor dissociation, functional locked-in syndrome, non-behavioral MCS, and higher-order cortex motor dissociation (Giacino et al., 2018a,b). These conditions are characterized by a dissociation between a dysfunctional behavioral motor output and a preserved cognitive output, which is measurable only by using paraclinical approaches to empirically demonstrate command following or similar indirect measurements of brain response to external stimuli that represent unequivocal signs of awareness (Klein, 2017; Kondziella et al., 2020). Furthermore, robust and relevant stimuli can be used to detect covert awareness otherwise labeled as UWS. For example, this has been achieved by providing patients with the subject’s name and detecting the event-related (de)synchronization (Fellinger et al., 2011). Providing a patient having UWS with relevant and entraining information in the context of simple motor tasks can more readily trigger the stimulus-related neural process, potentially supporting awareness. Indeed, relevant stimuli can give important bottom-up information that, in turn, might facilitate top-down attention processes (Jain & Ramakrishnan, 2020). Conversely, highly demanding cognitive tasks (requiring sustained attention, sensory processing, or remembering an instruction) can result in false-negative results and missing unruly behavior in non-communicative individuals (Kondziella et al., 2015).
Among the active paradigms that could be easy to perform on a patient with PDOC, a test of the interaction between motor observation and imagery might be promising (Chen, Shan, Guo & Yuan, 2013; Pistoia, Sacco, Carolei & Sarà, 2013; Naro et al., 2018; Kingwell, 2013).
In this regard, it has been demonstrated that action observation (watching movements performed by others) triggers a corresponding motor representation in the observer, which can interfere with the individual’s action representation (i.e., action simulation, the internal representation of motor programs without overt movement)(Jeannerod, 2001; Brass, Bekkering & Prinz, 2001; Brass, Derrfuss & Von Cramon, 2005; Brass et al., 2003; Stürmer, Aschersleben & Prinz, 2000). In such cases, high-level cognitive mechanisms at the level of awareness might help individuals distinguish the self from the observed other (Steinbeis, 2016). This can be demonstrated by recording specific visual event-related potentials (ERPs) while testing the influence of action observation on motor execution (Deschrijver, Wiersema & Brass, 2017). These consist of the stimulus-locked response (N190), which is a marker of the visual processing of body parts (Arzy, Thut, Mohr, Michel & Blanke, 2006; Myers & Sowden, 2008; Thierry et al., 2006), the response-locked readiness potential (RP), which correlates with the complexity of motor preparation (Leuthold & Schröter, 2011; Rigoni, Brass, Roger, Vidal & Sartori, 2013), and the central P3 component, which is related to the self-versus-other distinction (Deschrijver, Wiersema & Brass, 2015; Graux et al., 2013; Knyazev, 2013; Kühn et al., 2011; Perrin et al., 2005; Sebanz, Knoblich, Prinz & Wascher, 2006). Overall, these components are the neural correlates of the high-level awareness processes of social cognition (Brass et al., 2005; Santiesteban et al., 2012; Spengler, Bird & Brass, 2010; Spengler, Von Cramon & Brass, 2009a, 2009b), which are involved in the detection of self-relevant stimuli and the ability to follow instructions (Schäfer, Wentura & Frings, 2020). These processes might represent signs of cognition that are thought to be based on functional connectivity within the temporoparietal junction and the medial prefrontal cortex (Brass et al., 2005; Sowden & Catmur, 2013; Spengler et al., 2009a, 2009b, 2010).
Given the clinical picture of patients with PDOC, particularly those with UWS, action execution is not easy, and it is difficult to discern the influence of an action on perception, the influence of perception on an action, or conflict monitoring. Therefore, we sought confirmation on whether a strategy based on motor imagery (i.e., imagining the execution of an action without physically performing it) is a valid “surrogate” of motor execution in patients with chronic disorders of consciousness. In fMRI- or EEG-based active paradigms tailored for patients with PDOC, an action can be considered an effect of motor imagery, or it is conceivable that an action simulation is equivalent to motor execution or action observation (Jeannerod, 2001; Lorey et al., 2013; Hardwick, Caspers, Eickhoff & Swinnen, 2018; Gazzola & Keysers, 2009; Conson, Sarà, Pistoia & Trojano, 2009; Eaves, Riach, Holmes & Wright, 2016). In this regard, some studies combined motor imagery and action observation (Mulder, 2007; Conson, Sarà, Pistoia & Trojano, 2009; Kim, Frank & Schack, 2017; Eaves, Riach, Holmes & Wright, 2016; Emerson, Binks, Scott, Kenny & Eaves, 2018; Hardwick, Caspers, Eickhoff & Swinnen, 2018; Vogt, Di Rienzo, Collet, Collins & Guillot, 2013), and highlighted the neural networks and mechanisms underlying the control over the content, vividness, and awareness of action simulation (Holmes and Calmels, 2008; Rektor, Sochůrková & Bocková, 2006; Zhu, Sun & Wang, 2013; van Wijk, Beek & Daffertshofer, 2012; Calmels, Hars, Holmes, Jarry & Stam, 2008; Calmels et al., 2006a, 2006b). Specifically, motor imagery and action observation share neural substrates with movement execution (Greenwald, 1970; Prinz, 1987; Stock & Stock, 2004; Jeannerod, 1994; Shin, Proctor & Capaldi, 2010; Lee-Hand & Knott, 2015; Crammond, 1997; Grèzes & Decety, 2001; Jeannerod, 2001; Naro et al., 2018). Therefore, activation of a neural network during various motor situations should be related to functional equivalence between motor imagery, action observation, and movement execution (Grèzes & Decety, 2001; Jeannerod, 2001), even though some networks are involved in motor imagery and action observation (Hétu et al., 2013; Caspers, Zilles, Laird & Eickhoff, 2010). Motor imagery recruits a network of premotor-parietal cortical regions beyond the thalamus, putamen, and cerebellum. Action observation involves a cortical premotor-parietal and occipital network without significant subcortical contributions. Lastly, motor execution engages sensorimotor– premotor areas, along with the thalamus, putamen, and cerebellum. Therefore, whereas there are some key differences in the area recruited among the motor tasks, the activation of premotor-parietal and somatosensory networks is consistent, which is a fundamental prerequisite for hidden awareness recognition (Nani & Tanaka, 2019; Morales & Lau, 2020; Tsuchiya, Oizumi, Yamada & Saigo, 2020; Cohen, Ortego, Kyroudis & Pitts, 2020). Consequently, motor imagery would interact with action observation in ERP components, as previously demonstrated in motor execution and action observation (Deschrijver et al., 2017; Nagai & Tanaka, 2019; Hardwick et al., 2018). However, to the best of our knowledge, the former interaction has yet to be demonstrated. The model of equivalence between motor imagery and motor execution is also consistent with the ideomotor theory of action, according to which, observing, imagining, or representing an action excites the motor programs used to execute that same action; in other words, an action is generated by having a thought about that action (Greenwald, 1970; Prinz, W; Stock & Stock, 2004; Jeannerod, 1994; Shin et al., 2010; Lee-Hand & Knott, 2015). This occurs when, for instance, a patient observes motor actions that are either congruent or not with the imagined one (Riach, Holmes, Franklin & Wright, 2018; Vogt, Di Rienzo, Collet, Collins & Guillot, 2013; Ramsey, Cumming, Eastough & Edwards, 2010; Roberts, Wood & Wakefield, 2020; Eaves, Riach, Holmes & Wright, 2016; Deschrijver et al., 2015, 2017; Rauchbauer, Pfabigan & Lamm, 2018). It also occurs when providing a patient walking on a treadmill for gait training with the third-person or first-person view of a human avatar walking through a landscape (Caramazza, Anzellotti, Strnad & Lingnau, 2014; Hioka et al., 2019; Jin-seop & Kyoung, 2012; Calabrò et al., 2017; Kilner & Blakemore, 2007; Gallagher, 2011; Galvan Debarba et al., 2017; Gorisse, Christmann, Armand Amato & Richir, 2017), analogous to the different effects that first-person and third-person views of motor observation have on motor execution/imagery (Fourkas, Avenanti, Urgesi & Aglioti, 2006; Hanakawa, 2016; Abbruzzese, Avanzino, Marchese & Pelosin, 2015; Mulder, 2007; Emerson, Binks, Scott, Kenny & Eaves, 2018; Brass, Bekkering & Prinz, 2001; Stürmer, Aschersleben & Prinz, 2000; Catmur & Heyes, 2011; Eaves et al., 2016; Vogt et al., 2013). Indeed, one tends to mentally perform an observed movement in an automatic and unaware manner, as indicated by brain responses that do not support aware cognitive processes, even without an overt behavioral output (i.e., perceptual induction). However, any difference between what one might have expected to see in a motor observation/imagery task and what is seen evokes brain responses that support an aware cognitive process (i.e., intentional induction) (Meers, Nuttall & Vogt, 2020; Bruton et al., 2020; Eaves et al., 2016; Overgaard, 2012). Therefore, a conflicting scenario between motor imagery (i.e., the action one imagines and expects to see) and action observation (what is observed) might result in aware sensorimotor processing (Gatti et al., 2013, 2019; Mulder, 2007; Eaves et al., 2016). This can occur at either the content level [e.g., a movement observation that is (in) congruent with that imagined] (Riach et al., 2018; Vogt et al., 2013; Ramsey et al., 2010; Roberts et al., 2020; Eaves et al., 2016; Deschrijver et al., 2015, 2017; Rauchbauer et al., 2018) or the motor scenery level (e.g., visual perspective and motor scenarios)(Fourkas et al., 2006; Hanakawa, 2016; Abbruzzese et al., 2015; Mulder, 2007; Emerson et al., 2018; Brass et al., 2001; Stürmer et al., 2000; Catmur & Heyes, 2011; Eaves et al., 2016; Vogt et al., 2013), and is consistent with recent observations on virtual-reality-based human gait (Caramazza et al., 2014; Hioka et al., 2019; Jin-seop & Kyoung, 2012; Calabrò et al., 2017; Kilner & Blakemore, 2007; Gallagher, 2011; Galvan Debarba et al., 2017; Gorisse et al., 2017).
Based on these premises, our study aimed to reveal which ERP features during motor observation/imagery tasks account for differential diagnosis and covert awareness in a sample of patients clinically labeled with UWS compared to MCS patients and control group (CG) individuals. Indeed, patients with covert awareness (both patients with UWS and those with MCS who typically fluctuate between awareness and unawareness even within a short period) may fail to follow commands (Cruse et al., 2012). Therefore, finding a way to capture the residual awareness of these patients is mandatory within the framework of active paradigm testing (i.e., those aimed at identifying unequivocal signs of awareness, including command following and cognitive-emotional responses) (Andrews et al., 1996; Cruse et al., 2011, 2012; Owen et al., 2006; Monti et al., 2010; Schnakers et al., 2008). Capturing the effects of action observation on motor imagery might be promising in terms of conscious-level achievement in specific sensorimotor processing, but this is yet to be demonstrated in patients with severe awareness impairment, including PDOC. We only know that action observation does not necessarily entrain awareness, even in healthy people, and that motor imagery, as used in the current active paradigms, might be challenging for people with severely limited cognitive abilities, including those with PDOC. A simple motor task that probes the effects of action observation on motor imagery, might instead more easily reveal the residual cognitive abilities of patients with PDOC, because it would represent a cognitively silent stimulus to be processed (as in the case of self-referential sensorimotor tasks) in a severely deprived of stimuli and damaged brain (Owen, 2019; Stafford, Owen & Fernández-Espejo, 2019; Chleboun, Hux & Snell, 2009; Haugg et al., 2018; Kondziella, 2018). Specifically, we can hypothesize that ERP features consistently differ between patients with UWS and MCS in the degree of awareness and cognitive impairment. The former might not show any significant response to the task (consistent with the degree of awareness and cognitive impairment). In contrast, the latter might show significantly different ERPs as a result of action observation on motor imagery (i.e., a summation effect of a properly processed expected scenario— the first-person view— compared to unexpected ones— a third-person view). Furthermore, a simple approach such as this (in terms of procedures and instrumentation) should offer the advantages of millisecond-range resolution, low cost, non-invasiveness, and high adaptability for clinical use, including bedside monitoring.
Materials and methods
Participants
Twenty patients were enrolled in this exploratory study: 10 were diagnosed with UWS and 10 with MCS. A sample of 10 age- and gender-matched healthy individuals (CG) was also involved. Two independent raters diagnosed PDOC patients using the CRS-R (Giacino, Kalmar & Whyte, 2004).
The exclusion criteria were: a PDOCPDOC condition lasting less than 3 months after the injury to the brain; pre-existing severe neurological or systemic diseases; actual critical conditions (i.e., inability to breathe independently, hemodynamic instability); the use of cortical-excitability-modifying drugs, other than l-DOPA, baclofen, and antiepileptic drugs; absence of visual and auditory evoked potential. Clinical-demographic information is reported in Table 1. A relative or legal representative of each patient, and the healthy participants, gave written informed consent. The local ethics committee (IRCCS Centro Neurolesi Bonino-Pulejo; Messina, Italy) approved the study.
Clinical-demographic characteristics of the sample
Clinical-demographic characteristics of the sample
Legend: DAI diffuse axonal injury, WMH white matter hyperintensity, F frontal, Fb frontobasal, FP frontoparietal, _h hemorrhagic lesion, _is ischemia, SAH subarachnoid hemorrhage, TPO temporo-parieto-occipital, dd disease duration, T traumatic, V vascular, A anoxic.
Patients lay in their beds (GC subjects sat in a comfortable armchair) in a dimly lit and sound-attenuated room. The CRS-R arousal protocol was carried out before starting the experimental session. Participants were instructed (both verbally and using a video recording immediately before the experiment began) to imagine, while watching a video, walking down a street in a first-person perspective. Notably, they were instructed to freely observe any part of the video picture being shown, without paying attention to any particular detail (as the surrounding scenery might have been different); instead they were to focus on their muscle activation as if they were genuinely walking down a street. A black screen was presented before each trial for 2 secs, serving to notify the patient to prepare for motor imagery. The video showed either a walk down a street from a first- or third-person perspective, or a first-person view of a street (a brief example for each condition, respectively provided in Video_1). Therefore, in the first two of these scenarios, the patient’s imagined walk was either from the same perspective as the video they observed or different. Thirdly, in the no-movement trial, the participant was required to perform the motor imagery of a first-person view of walking down a street while watching a stationary first-person view of a street. Each motor imagery trial lasted 10 secs. Each task was presented 75 times such that the same task was repeated consecutively no more than twice, but in an otherwise random order.
The video was displayed on a 6-inch screen placed approximately 15 cm in front of the participant. The participant’s head was held in position by a fixing device. This, and clinical inspection that attention was being maintained, ensured the patient watched the video for the time required by the experiment We also checked for episodes of under-arousal, that is, a contract between the upper and lower eyelids maintained continuously for more than 3 secs. In such circumstances, the task was paused, the arousal facilitation protocol was administered, and then the task was resumed.
EEG recording and analysis
Electroencephalograms were recorded during the entire task (approximately 15 mins) using a standard headset with 21 electrodes placed according to the International 10– 20 system, wired to a BrainQuick device (Micromed; Mogliano Veneto, Treviso, Italy). Two EOG channels were also used to measure eye movements (bipolar electrodes placed with left and right canthal montage, and above and below the left eye). The sampling rate was 512 Hz, the band-pass filter was 0.1– 45 Hz, and both mastoids were set as references. Electrode impedance was always below 5 kΩ. Data were segmented in [– 2;5] sec epochs according to task. Then, bad epochs (determined by visual inspection and ICA) were discarded, to give approximately 50 epochs for each trial.
We first built the topographical maps of the average N190 and P3 for the CG, and identified the electrodes of interest (P5– P7 and Pz, respectively)(Fig. 1). In this regard, the N190 and P3 were time-locked to the onset of the frame ([170:220]ms and [310;430]ms, respectively). The RP consisted of the response-locked gradient shift preceding the steep negative slope before onset of motor imagery at electrode FCz ([– 400;– 100]ms) (Fig. 1) (Leuthold & Schröter, 2011; Rigoni et al., 2013; Shibasaki & Hallett, 2006). Therefore, we performed the analyses on the exported mean curve area from pooled P5– P7, Pz, and FCz using two-way ANOVA with the factors group (three levels: UWS, MCS, and CG) and trial (two levels: first-person and third-person view). We opted to analyze only these two conditions because the baseline condition (the no-movement trial) was not suited to the EEG setting but rather to keep patient participation in the task high. Indeed, the effects of motor observation on motor imagery in neuroimaging studies are usually analyzed without involving the baseline condition (e.g., Brass et al., 2005). The laterality effect for the N190 was also included in the analysis. Greenhouse– Geisser corrections were applied if appropriate. Bonferroni-corrected post-hoc t-tests were conducted.
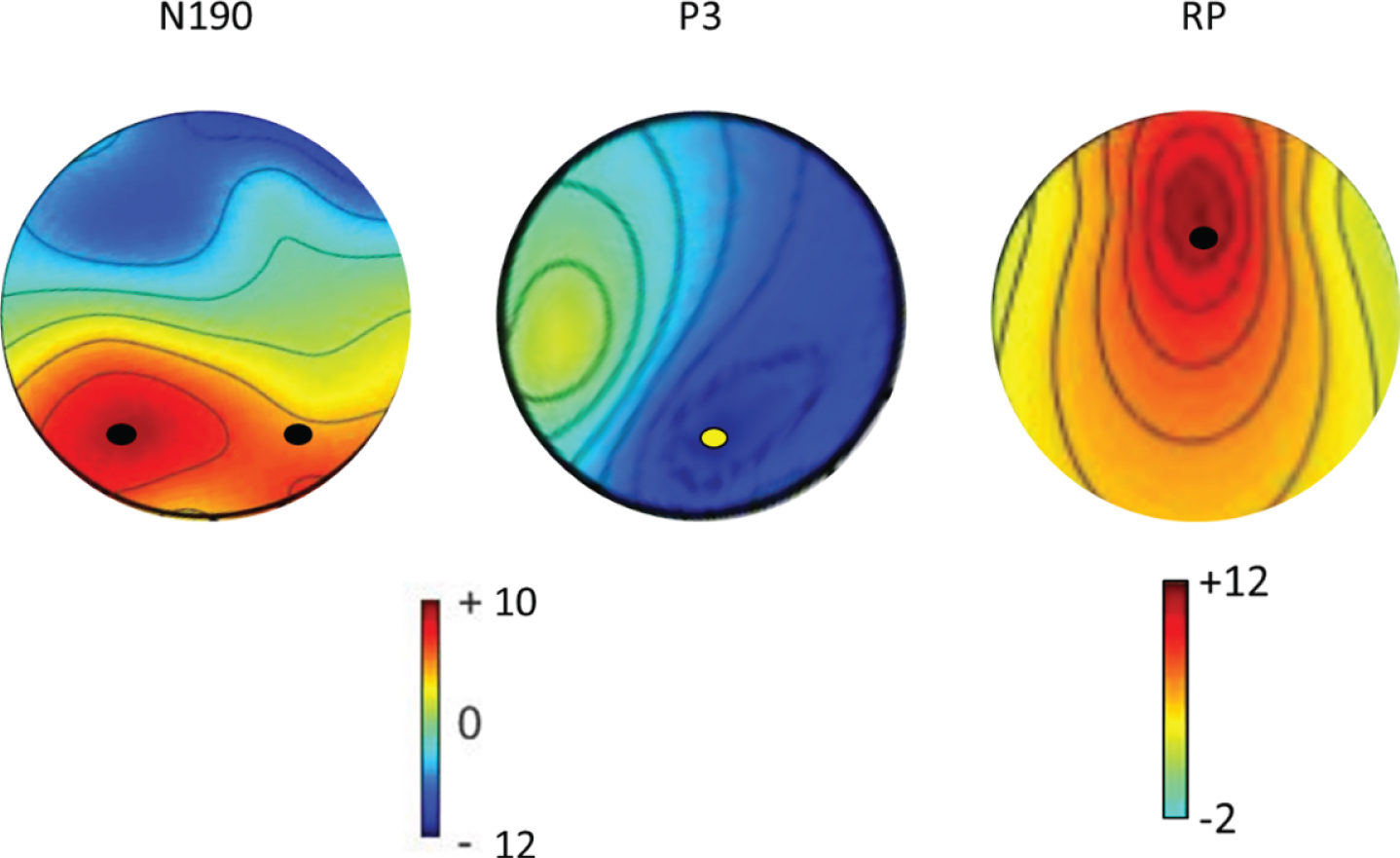
Topography maps of the average ERPs (N190, P3, and RP) in their respective periods of interest and relevant electrode sites in the control group. The amplitude is reported in the color bar (μV for N190 and P3, Laplacian transformations for RP μV/m2). Electrodes of interest (P5, P7 for N190, Pz for P3, and FCz for RP) are marked by a dot.
Within-group evaluation
The CG showed a significant trial effect for each of the evoked components, which were clearly appreciable at about – 200 ms (RP; F (1,9) = 18, p < 0.0001, η2 = 0.996), 200 ms (N190; F (1,9) = 20, p < 0.0001, η2 = 0.997), and 350 ms (P3; F (1,9) = 43, p < 0.0001, η2 = 0.999) with respect to the stimulus onset (the beginning of the video) (Figs. 2-3). At the individual level, all participants showed responses of different magnitudes to the first-person and third-person view trials. These responses also had distinct topographies.
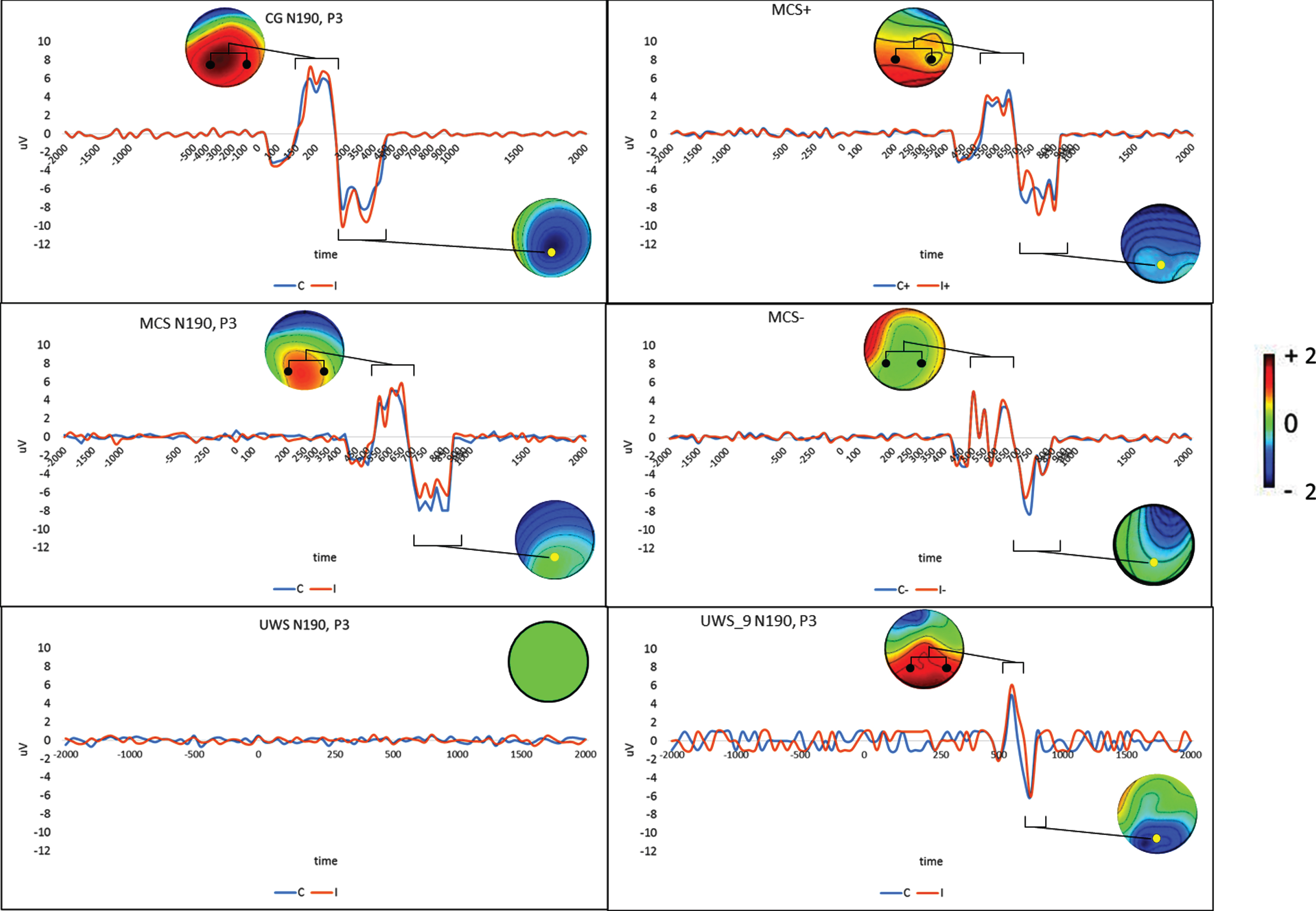
N190 and P3 components over their relevant electrodes (pooled-P5/P7 for N190 and Pz for P3, respectively, marked by a dot in the topographical map) for the first-person (C) and third-person view (I) conditions in the control group (CG; n = 10) and the MCS (n = 10) and UWS samples (n = 10) (left column). The differences between patients with MCS who were able to differentiate (MCS+; n = 6) or not (MCS-; n = 4) between C and I condition and the response profile in the UWS patient no. 9 are reported in the right column. Topographical maps of the I– C difference for each ERP (expressed as μV in the color bar) in their respective periods of interest (square graphs) are also reported.
The N190 responses measured at both parietal electrodes were greater for participants viewing the third-person perspective than for those viewing the first-person perspective (post hoc t-test t (9) = 3.25, p = 0.009; laterality effect p > 0.1). The P3 response at the parietal midline electrodes was instead greater for participants viewing the first-person perspective than for those viewing the third-person perspective (post hoc t-test t (9) = 3.35, p = 0.008). The RP response, measured by the frontocentral midline electrodes, was greater for participants viewing the first-person perspective than for those viewing the third-person perspective (post hoc t-test t (9) = 3.67, p = 0.005)(Figs. 2-3).
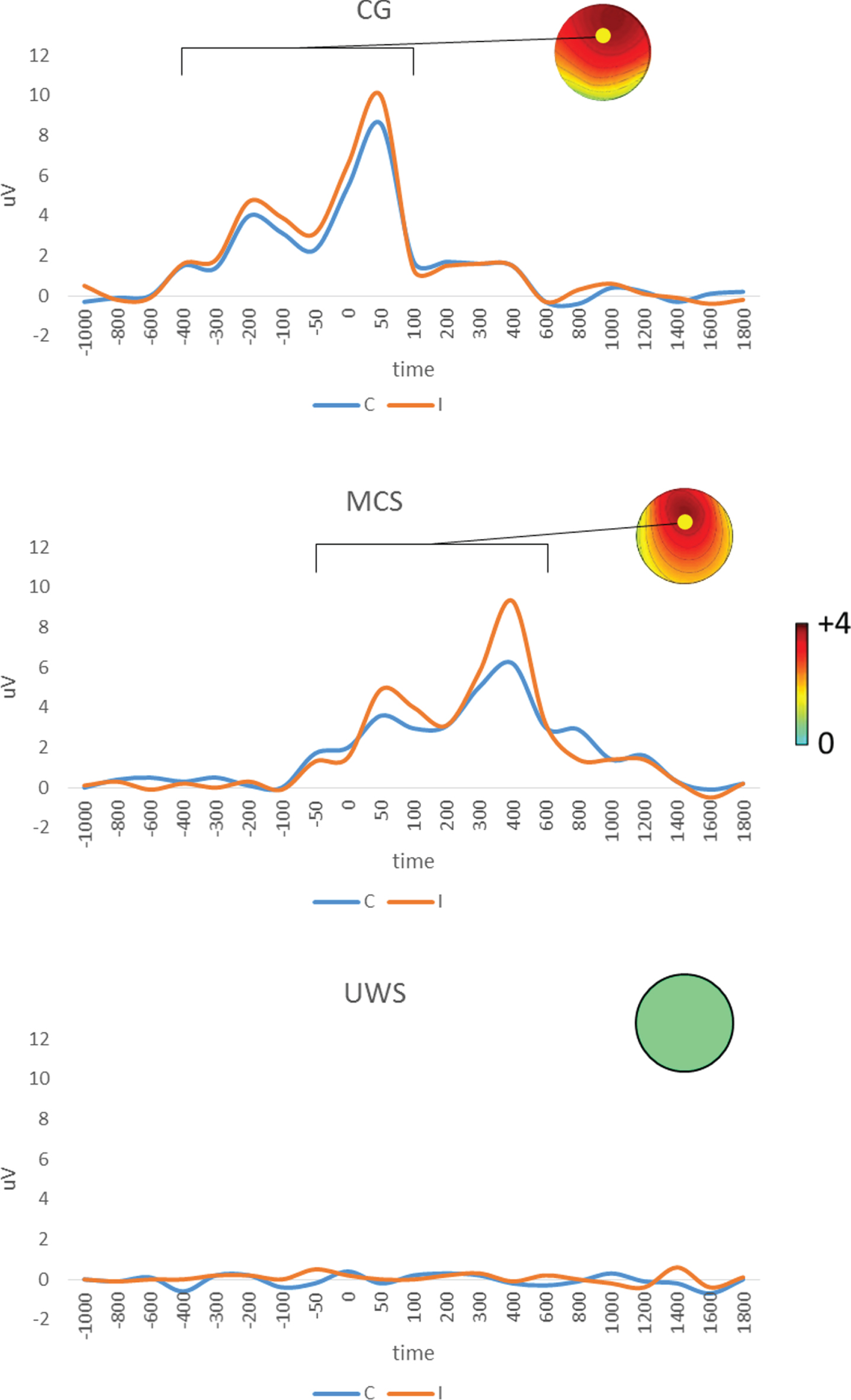
RP component as pooled event-related potentials over FCz (marked by a dot in the topographical map) for the first-person (C) and third-person view (I) conditions in the control group (CG; n = 10) and the MCS (n = 10) and UWS (n = 10) samples. Topographical maps of the I– C difference for each ERP (expressed as μV in the color bar) in their respective periods of interest (square graph) are also reported.
The MCS group showed a significant trial effect for each of the evoked components, which were clearly apparent at approximately – 200 ms (RP; F (1,9) = 60, p < 0.0001, η2 = 0.999), 200 ms (N190; F (1,9) = 19, p < 0.0001, η2 = 0.997), and 350 ms (P3; F (1,9) = 34, p < 0.0001, η2 = 0.999) with respect to the onset of stimulation (the beginning of the video)(Figs. 2-3). Six out of 10 of these participants showed different magnitudes of responses to the first-person and third-person view trials. Their responses had the same distinct topographies of the CG but were significantly delayed and with a clear temporal dispersion (i.e., a polyphasic morphology). The remaining MCS patients showed ERPs with the same latency, but without a difference in magnitude between first- and third-person view trials, and less temporal dispersion and RP (Figs. 2-3)(N190 post hoc t-test t (9) = 3.25, p = 0.01; P3 post hoc t-test t (9) = 8.8, p < 0.0001; and RP post hoc t-test t (9) = 3.2, p = 0.01).
We could not elicit even the slightest response in any of the patients in the UWS group. However, one (n.9) showed N190 and P3 responses to the provided video that were similar, albeit even smaller and more delayed, to those observed in those MCS patients unable to differentiate between first-person and third-person view trials (Fig. 2). An RP response was not detected.
ANOVA testing did not show a significant group ×trial interaction with respect to the N190 response. Therefore, there were no significant differences between the groups concerning the distinction between first- and third-person views. The group× trial×laterality interaction was also not significant. Conversely, we found a significant group×trial interaction concerning the P3 response (F (2,54) = 2.87, p = 0.04, η2 = 0.891). Indeed, the difference in magnitude between the first-person and third-person view trials was not significantly different between the CG and MCS group (p = 0.2), but it was significantly greater in the CG than in the UWS group (p < 0.001) and in MCS versus UWS (p = 0.03).
ANOVA also revealed a significant trial×group main interaction in the RP response (F (2,54) = 7.2, p = 0.001, η2 = 0.981). Indeed, the difference in magnitude between first-person and third-person view trials was significantly greater in the CG than in the MCS group (p = 0.02), in CG versus UWS (p = 0.007), and in MCS versus UWS (p = 0.01).
Discussion
Action observation is defined as the deliberate and structured observation of human movement (Neuman & Gray, 2013). This evokes a motor representation in the sensorimotor areas that neither implies movement nor occurs consciously. Motor imagery is defined as the mental rehearsal of human movement without overt movement (Guillot & Collet, 2008). It undoubtedly implies awareness, but can be difficult for patients with PDOC to carry out. In this study, we assimilated a first-person view of walking down the street to the action observation of human walking in an attempt to maximize the cognitive entrainment of patients with PDOC, rather than merely providing a view of a movement as per the standard paradigms of action congruency without a difference in perspective (Riach et al., 2018; Vogt et al., 2013; Ramsey et al., 2010; Roberts et al., 2020; Eaves et al., 2016; Deschrijver et al., 2015, 2017; Rauchbauer et al., 2018). Motor imagery tasks have to be simple to execute but cognitively entraining, as are those already integrated into fMRI and EEG studies (Cruse, Chennu, Fernández-Espejo, Payne, Young & Owen, 2012; Bodien, Giacino & Edlow, 2017; Fernández-Espejo, Norton & Owen, 2014; Owen, Coleman, Boly, Davis, Laureys & Pickard, 2007; Marino et al., 2017; Owen & Coleman, 2008; Vogel, Markl, Yu, Kotchoubey, Lang & Müller, 2013; Naro & Calabrò, 2020).
The utility of coupling both tasks to activate cognitive processes in patients with PDOC arises from the capacity of the coupled tasks to enhance the motor output, as reported in rehabilitation and sports studies (de Vries & Mulder, 2007; Ste-Marie, Law, Rymal, McCullagh & Hall, 2012). This principle can then be harnessed to reveal covert behavioral output in individuals with PDOC. Given that motor execution is hugely challenging in patients with PDOC, we adopted a motor imagery task as a valid surrogate of motor execution because these two forms of motor simulation are associated with activity in regions of the motor system that partly overlap with those involved in motor execution (including the supplementary motor area, the premotor cortex, the superior parietal lobe, and the intraparietal sulcus)(Jeannerod, 2001; Bruton, Holmes, Eaves, Franklin & Wright, 2020). Furthermore, we looked at the responsiveness of healthy individuals to such a motor observation– imagery task. We found that all healthy individuals showed a more significant N190 response when motor observation and motor imagery did not match (i.e., third- and first-person views, respectively) than if motor observation and motor imagery matched (i.e., both from the first-person perspective). Conversely, the P3 and RP responses were more significant if motor observation and motor imagery corresponded (i.e., both first-person views) than if they did not (i.e., third- and first-person views, respectively).
Recent studies have investigated the efficacy of combining the two motor simulation types in terms of the congruency effect (Riach et al., 2018; Vogt et al., 2013; Ramsey et al., 2010; Roberts et al., 2020; Eaves et al., 2016; Deschrijver et al., 2015, 2017; Rauchbauer et al., 2018; Eaves, Behmer & Vogt, 2016; Eaves, Haythornthwaite & Vogt, 2014; Eaves, Riach, Holmes & Wright, 2016; Eaves, Turgeon & Vogt, 2012). By contrast, we approached our patients by mirroring a walk along a street either from a first-person or a third-person perspective, as either condition can activate the sensorimotor areas related to gait using mirror neuron system-related areas, as suggested by studies that provided healthy and post-stroke individuals with virtual reality feedback of a human avatar while walking during robot-aided gait training (Fourkas et al., 2006; Hanakawa, 2016; Abbruzzese et al., 2015; Mulder, 2007; Emerson et al., 2018; Brass et al., 2001; Stürmer et al., 2000; Catmur & Heyes, 2011; Eaves et al., 2016; Vogt et al., 2013; Galvan Debarba et al., 2017; Gorisse et al., 2017; Caramazza et al., 2014; Hioka et al., 2019; Jin-seop & Kyoung, 2012; Calabrò et al., 2017; Kilner & Blakemore, 2007; Gallagher, 2011; Galvan Debarba et al., 2017; Gorisse et al., 2017). There could be a concern that the perspective through which a participant observes the actions of others can affect the neural effects evoked by the action observation itself, independent of the identity of the action per se (Fourkas et al., 2006; Hanakawa, 2016; Abbruzzese et al., 2015; Mulder, 2007; Emerson et al., 2018; Brass et al., 2001; Stürmer et al., 2000; Catmur & Heyes, 2011; Eaves et al., 2016; Vogt et al., 2013). However, EEG recordings measured different brain signals in individuals walking alongside a walking human avatar from those without (i.e., a first-person and third-person view of walking, respectively) (Fourkas et al., 2006; Hanakawa, 2016; Abbruzzese et al., 2015; Mulder, 2007; Emerson et al., 2018; Brass et al., 2001; Stürmer et al., 2000; Catmur & Heyes, 2011; Eaves et al., 2016; Vogt et al., 2013; Galvan Debarba et al., 2017; Gorisse et al., 2017; Caramazza et al., 2014; Hioka et al., 2019; Jin-seop & Kyoung, 2012; Calabrò et al., 2017; Kilner & Blakemore, 2007; Gallagher, 2011). Furthermore, for each condition, it is plausible that other visible individuals who are walking or moving near or from afar, will be mirrored by the brain in any case. The conditions also include their movements, walking, following someone, or their head movement. However, all of these additional stimuli are necessary to naturally entrain the limited cognitive reservoir of PDOC patients, as previously suggested (Cruse et al., 2012; Bodien et al., 2017; Fernández-Espejo et al., 2014; Owen et al., 2007; Marino et al., 2017; Owen & Coleman, 2008; Vogel et al., 2013; Naro & Calabrò, 2020). Furthermore, the surroundings in first- or third-person views have not been reported to influence motor performance or brain signal output in neurological individuals (De Keersmaecker, Lefeber, Geys, Jespers, Kerckhofs & Swinnen, 2019; Myung-Joon, 2020; de Rooij, van de Port, Visser-Meily & Meijer, 2019; Naro & Calabrò, 2020). Besides, these additional stimuli were inconsistent from one video to another, and participants were asked to not focus on any particular aspect of the video but rather to concentrate on their own first-person view as if they were walking down the street. Therefore, these additional potential stimuli are irrelevant.
Our data are consistent with those reported in functional neuroimaging studies on the influence of action on perception in heightening visuomotor processing, as indicated by the N190 response (greater for participants viewing the third-person perspective than for those viewing the first-person perspective). This is thought to be the expression of the processing of body-related visual information within the temporo-parieto-occipital junction (Borhani, Borgomaneri, Ládavas & Bertini, 2016; Borhani, Ládavas, Maier, Avenanti & Bertini, 2015; Thierry et al., 2006). Furthermore, our data agree with the effect of perception on the action in heightening the individual motor representation, as indicated by the RP and P3 responses being greater for the first-person view versus the third-person view (Brass et al., 2005; Spengler et al. 2009a, 2009b, 2010). These are thought to be, respectively, the expression of motor preparation processes within the pre-supplementary motor area, premotor cortex and primary motor cortex (Vercillo, O’Neil & Jiang., 2018), and the sensorimotor processing within the temporoparietal junction and centroparietal areas in response to conflicting or non-motor observation– imagery, stimulus-related evaluation and decision, novelty processing, and updating of working memory (Asanowicz et al., 2020). All such issues are grounded in analogy to ERP studies investigating a visual (N190), motor (RP), and cognitive (P3) components, which underlie the influence of, respectively, action on perception, perception on the action (RP), and of (in)congruency processing (Riach et al., 2018; Vogt et al., 2013; Ramsey et al., 2010; Roberts et al., 2020; Eaves et al., 2016; Deschrijver et al., 2015, 2017; Rauchbauer et al., 2018). One might disagree that we substituted the congruency effect with a more general discrepancy between a person’s view of an observed/imagined motor scenario (i.e., perspective). However, by complementing the motor observation– imagery tasks, we replicated the results obtained when adopting a real congruent/incongruent motor task (Riach et al., 2018; Vogt et al., 2013; Ramsey et al., 2010; Roberts et al., 2020; Eaves et al., 2016; Deschrijver et al., 2015, 2017; Rauchbauer et al., 2018), confirming that the sensorimotor processes underlying the effect of motor observation on motor imagery rely on both high-level (P3) as well as low-level (N190 and RP) cognitive processes (even when using a different perspective rather than a natural motor interference paradigm). Our data also indicate that such an approach faces the conscious level by sensorimotor information processing even when providing the patient with a different visual perspective on an observed imagined motor scenario. In this regard, to the best of our knowledge, the present study is the first to investigate the interaction between motor imagery and motor observation using EEG.
Nonetheless, there are some conflicting data in the literature on the meaning of motor action/observation-related EEG responses. For instance, the N190 response (visuomotor processing) is not always measurable, suggesting that high-level processing of self-versus-other motor actions might be specifically preserved, more so than other evoked responses (Riach et al., 2018; Vogt et al., 2013; Ramsey et al., 2010; Roberts et al., 2020; Eaves et al., 2016; Deschrijver et al., 2015, 2017; Rauchbauer et al., 2018). It is notable that the methodological differences among the aforementioned studies (including the stimulus type, setting, and EEG signal analysis) might account for such discrepancies. Nonetheless, the retention of self-versus-other sensorimotor processes might account for the differences found between MCS and UWS patients. Remarkably, six out of 10 patients with MCS showed preservation of both high- and low-level sensorimotor processing, although the amplitude of the responses was significantly lower and the latency was more delayed than in the CG. The other patients with MCS did not show diversified EEG responses, suggesting limited cognitive functioning. However, these individuals still showed high-level processing of self-versus-other motor actions, which was not the case for patients with UWS. One UWS patient behaved like those in the MCS group, that is, she showed a high-level self– other distinction centered on motor actions while exhibiting a compromised motor preparation process. Therefore, the changes in ERP magnitude in response to the various motor scenarios allowed patients with MCS and UWS to be differentiated at the group level but not at the individual level. The possibility to monitor patients at the bedside, the straightforward interpretation of EEG findings, and the low cost and non-invasiveness of the approach make the methodology reported here highly adaptable for clinical use.
ERP effects and awareness
The main concern with our data is whether the present brain responses necessarily indicate awareness. Purposeful following of motor commands during a clinical examination or functional neuroimaging/electrophysiological assessment is unequivocal proof of awareness. Given that the task we adopted induced both high- and low-level sensorimotor processing, one might be concerned about the preservation of awareness. Indeed, we might have facilitated the visual processing of an observed motor scenario, as suggested by the modulation of the N190 amplitude between different conditions, which could be explained by the functional preservation of the extra-striate body area of the visual system (Thierry et al., 2006) and the capacity of patients with MCS to functionally allocate cognitive resources. This response does not, therefore, necessarily indicate whether a subject is aware. Nevertheless, an alternative train of thought assumes that N190 is related to processes of visual (Vocks et al., 2010) or embodied self– other discrimination, which is considered functionally distinct from high-level, more cognitive self– other distinction (Arzy et al., 2006), consistent with the difference between first- and third-person views (Caramazza, Anzellotti, Strnad & Lingnau, 2014; Hioka et al., 2019; Jin-seop & Kyoung, 2012; Calabrò et al., 2017; Kilner & Blakemore, 2007; Gallagher, 2011; Galvan Debarba et al., 2017; Gorisse, Christmann, Armand Amato & Richir, 2017). In keeping with this idea, the first-person view is more likely to be referred to one’s own body. Conversely, referring action observation in the third-person view to one’s own body is more exacting, and is more likely to evoke different brain responses, although this does not indicate preservation of awareness per se. The notion that N190 modulation suggests awareness preservation may be supported by the fact that providing patients with n repetitions of the same motor scenario induces specific entrainment of the mirror neuron system, which is likely to occur at a “more conscious” level (Perlovsky & Ilin, 2013; Caramazza et al., 2014; Capa et al., 2011; Naro et al., 2018; Naro & Calabrò, 2020).
The magnitude of RP indexed the mechanisms of imitative action preparation, albeit at a low level. In motor observation– imagery tasks, the RP was smaller in patients given the first-person view than in those receiving the third-person view. This could be because the patient’s actions were facilitated upon observing the scenario from a first-person perspective. However, this mechanism is below the awareness threshold (Clarke et al., 2019; Vogt et al., 2013).
Finally, the magnitude of P3, which could be related to high-level cognitive processes within the temporoparietal junction, the medial prefrontal cortex, and the precuneus (Asanowicz et al., 2020), was highly influenced by the first-person view, suggesting that P3 might be a neural correlate for self– other distinction. Both N190 and P3 data demonstrate that patients with MCS were able to differentiate among motor plans and between the self and other. These data are consistent with those related to self– other processing in PDOC patients, such as hearing one’s name or seeing one’s face (Staffen et al., 2006; Perrin et al., 2006). Moreover, our findings support the mirror neuron system having a pivotal role in visuomotor processing (Morsella, Godwin, Jantz, Krieger, & Gazzaley, 2016; Naro et al., 2018; Naro & Calabrò, 2020). Furthermore, P3 data show how processes underlying the modulation of the N190 amplitude might have exhausted the cognitive reserve for the difference in perspective in the motor scenarios, leading to fewer available resources for generating the P3 component (Kok, 2001; Polich, 2007). It is then reasonable to hypothesize that P3 amplitude may be related to the self– other distinction, even within the difference in perspective in the motor scenarios (Knyazev, 2013; Smigielski et al., 2020). Then, we might argue that high-level social-cognitive self– other distinction could be associated with awareness, even at a covert level, as potentially suggested by the findings on UWS patient n.9. However, four patients with MCS and one with UWS (n.9) showed similar ERPs in the first- and third-person view motor scenarios. The lack of a different response to the various motor scenarios might suggest a behavioral hyperimitation (perhaps owing to a dysfunctional mirror neuron system) because of a tonic hyperfacilitation of brain activity to recover function (Bagnato et al., 2013; Lapitskaya et al., 2013). Consistent with the concern that the behavioral responsiveness might be related to neural processes rather than high-level social cognitive ones, further studies involving motor imagery tasks are required to demonstrate a correlation between high-level social-cognitive self– other distinction and awareness.
Limitations and strengths
The abovementioned ERP findings, together with their possible anatomical substrates and functions (putatively belonging to the default-mode network) (Hardwick et al., 2018), might indicate the possibility of identifying preservation of awareness in patients with PDOC. Furthermore, the complexity in the timing of brain responses suggests a preserved functional connectivity within the internal and external awareness frontoparietal networks (Haugg et al., 2018; Bodien, Chatelle & Edlow, 2017; Naro et al., 2017), as well as a connectivity impairment (in MCS) or a breakdown (in UWS), as previously demonstrated (Casarotto et al., 2016).
The small and non-homogeneous sample represents the main limitation of the study. Consistent with the difficulty in recruiting a large sample of PDOC patients, further multicenter studies are required to confirm our promising findings.
Secondly, one may be concerned about the use of N190, RP, and, above all, P3 because other ERP components referring to, for example, enhanced attentional focus (N100) or motivational relevance of the presented stimuli (LPP) could have been included. These relevant aspects are potentially engaged in the various stages of task processing (Rauchbauer et al., 2018) and warrant further investigation.
Thirdly, RP (which is a marker of automatic motor preparation for a motor observation task) was temporally nested with N190 and P3, configuring a functional network supporting an aware response to different motor scenarios. However, we specifically focused on stimulus-locked N190, as this is a marker of visual processing of body part information (Arzy et al., 2006; Myers & Sowden, 2008; Thierry et al., 2006). The preservation of the neural networks supporting body part’s sensory processing is a fundamental prerequisite for real or imaginary motor planning and the subsequent sensory processing to distinguish self from other (Serino et al., 2015; MacIntyre et al., 2018). Since RP is known to magnify with the increasing complexities of motor preparation (Leuthold & Schröter, 2011; Rigoni et al., 2013), its preservation is a significant indicator of motor function maintenance, even at a subclinical level (i.e., not behaviorally overt), regardless of whether or not awareness is preserved.
Notably, we adopted an assessment of the centroparietal P3 component, since this is correlated with self-related processes in social cognitive paradigms (Graux et al., 2013; Holeckova et al., 2006; Knyazev, 2013; Kühn et al., 2011; Perrin et al., 2005; Sebanz et al., 2006) and high-level comparative processing (Longo, Musil & Haggard, 2012; Zhou et al., 2004; Kühn et al., 2011; Macaluso, 2006; Yamaguchi & Knight, 1991). These ERP components allow the influence of action intentions on perception (visual ERP) and the influence of action observation on action preparation (ERP components related to motor preparation) to be evaluated. Therefore, especially given that specific differences were appreciable in all of the ERP components, they might represent a means of potential sensorimotor processing at a conscious level related to the self-versus-other distinction and action preparation when observing a scenario from a first-versus-third person view.
5 Conclusions
Despite its limitations, our study suggests that identifying low-level visual and high-level motor preparation processes in response to simple, first/third-person-perspective-centered motor observation/imagery tasks might potentially support the clinical diagnosis of MCS and UWS. Our protocol could pave the way for more accurate diagnoses of PDOC by identifying the patients with UWS who require more sophisticated diagnostic approaches. This would facilitate more functional diagnostic procedures and better allocation of resources.
Disclosures
Footnotes
Acknowledgments
We want to thank Prof. Adrian Neal (Peerwith) and Prof. Andy Gray and for English language revision.
Funding
No funding to be reported.
Conflict of interest
None of the authors have potential conflicts of interest to be disclosed.
Ethical approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards. The Institutional Review Board of IRCCS Centro Neurolesi Bonino Pulejo (Messina, Italy) approved the study.
Informed consent
Persons provided their written informed consent to study participation and data publication.
Declaration
We hereby confirm that our references and in-Text-citations are in accordance with the guidelines of the Journal Restorative Neurology and Neuroscience and that reference in the reference list and in the manuscript are complete and match.