Abstract
BACKGROUND:
Genetic manipulation on the NO-sGC-cGMP pathway has been rarely achieved, partially due to complexity of the soluble guanylyl cyclase (sGC) enzyme.
OBJECTIVE:
We aim to develop gene therapy directly targeting the pathway to circumvent cytotoxicity and tolerance after prolonged use of NO-donors and the insufficiency of PDE inhibitors.
METHODS:
In this study, we constructed lentivirus vectors expressing GUCY1A3 and GUCY1B3 genes, which encoded the
RESULTS:
Transductions of human HEK293 cells with the constructs were successful, as indicated by the fluorescent signal and altered gene expression produced by each vector. Overexpression of GUCY1A3 and GUCY1B3 resulted in increased sGC enzyme activity and elevated cGMP level in the cells. Expression of PDE5A shRNA resulted in decreased PDE5A expression and elevated cGMP level. Co-transduction of the three lentiviral vectors resulted in a more significant elevation of cGMP in HEK293 cells without obvious cytotoxicity.
CONCLUSION:
To the best of our knowledge, this is the first study to show that co-expression of exogenous subunits of the soluble guanylyl cyclase could form functional enzyme and increase cellular cGMP level in mammalian cells. Simultaneous expression of PDE5A shRNA could alleviate feedback up-regulation on PDE5A caused by cGMP elevation. Further studies are required to evaluate the effects of these constructs in vivo.
Introduction
Ischemic and hypertensive heart disease are the leading causes of death in the world over the past decade. Over 7.4 million people die of these diseases annually and the number is still rising [1, 2]. Multiple molecular pathways have been shown to regulate the critical balance between cell death and cell survival in heart ischemia [3]. Especially the discovery of nitric oxide (NO) as an endogenous signaling molecule and as a mediator of the cardiovascular effects was acknowledged by the Nobel Prize in Physiology and Medicine in 1998 [4].
The best characterized target for NO signal transduction is the heme-containing enzyme soluble guanylyl cyclase (sGC) [5]. sGC is a heterodimer contains
Understanding the NO-sGC-cGMP pathway has initiated significant translational interests to manage cardiovascular disease by manipulating cellular levels of cGMP both before and after the disease onset. So far, such efforts were almost exclusively embodied in the use of NO donors and phosphodiesterase (PDE) inhibitors [8, 9]. NO donors are not ideal for long-term treatment of heart diseases due to their partial patient responses, development of tolerance over time and short retention time in the patients’ plasma [9, 10]. The use of PDE inhibitors is also likely to be limited since the efficacy of such molecules depends on endogenous cGMP generation [11]. Although targeting the NO-sGC-cGMP axis with molecular medicines may help overcome these issues and lead to promising novel therapy for cardiovascular disease, it has not been studied whether co-expression of the
In this study, we successfully elevated cGMP level in the HEK293 cell line by simultaneous overexpression of the
Materials and method
Cell culture
Human Embryonic Kidney 293 (HEK293) cell line was purchased from American Tissue Culture Collection (ATCC) and cultured as recommended. Briefly, the cells were maintained in DMEM supplemented with 10% heat-inactivated fetal bovine serum (FBS) in an atmosphere of 95% air and 5% CO
Lentivirus
Lentiviral vectors carrying the
HEK293 cells were transfected with lentiviral vectors using a standard procedure. Briefly, cells were seeded to be 40% confluent at a 35 mm culture dish. 10
Gene expression analysis
Total RNA was isolated from cells using Trizol reagents (Life Technology, Shanghai, China) following the manufacturer’s protocol. Degradation of RNA was examined by 28 s versus 18 s ratio on 1% agarose gels. RNA concentrations were measured using a Nanodrop Vis-UV spectrophotometer (Thermo Scientific, USA).
Reverse transcription and amplification of genes was conducted using 100 ng of total RNA and a two-step RT-PCR protocol from Life Technology according to the manufacturer’s protocol (Life Technology, Shanghai, China). Gene expression was also examined quantitatively using the SYBR green method (Life Technology, Shanghai, China) using an Applied Biosystems 7900HT system. Relative expression of each gene was calculated as Delta CT vs. the house keeping gene GAPDH (CT
Protein extraction and enzyme-linked immunosorbent assay (ELISA)
Cells were digested by trypsin and collected in 1 ml Eppendorf tubes with 200
Statistical analysis
For gene expression analysis and ELISA, all samples were measured in triplicate. Data were presented as mean
Results
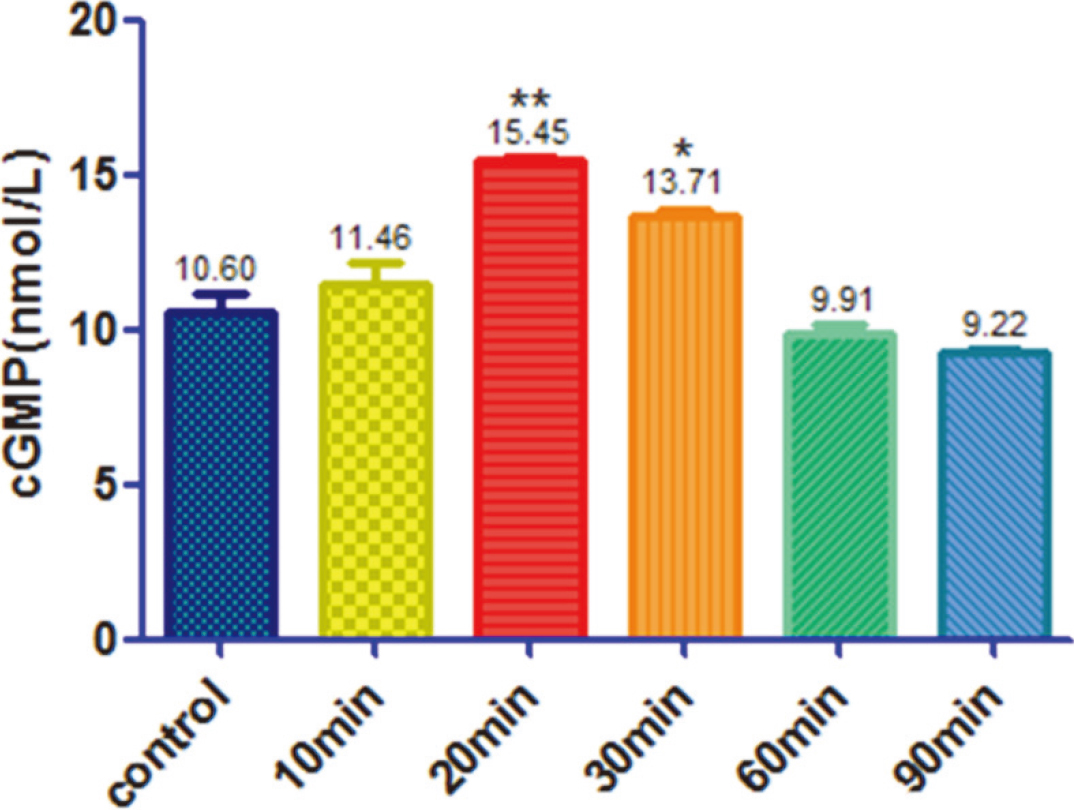
HEK293 cells were responsive to SNP treatment
HEK293 cells were treated with 10
Lentivirus transduction of the 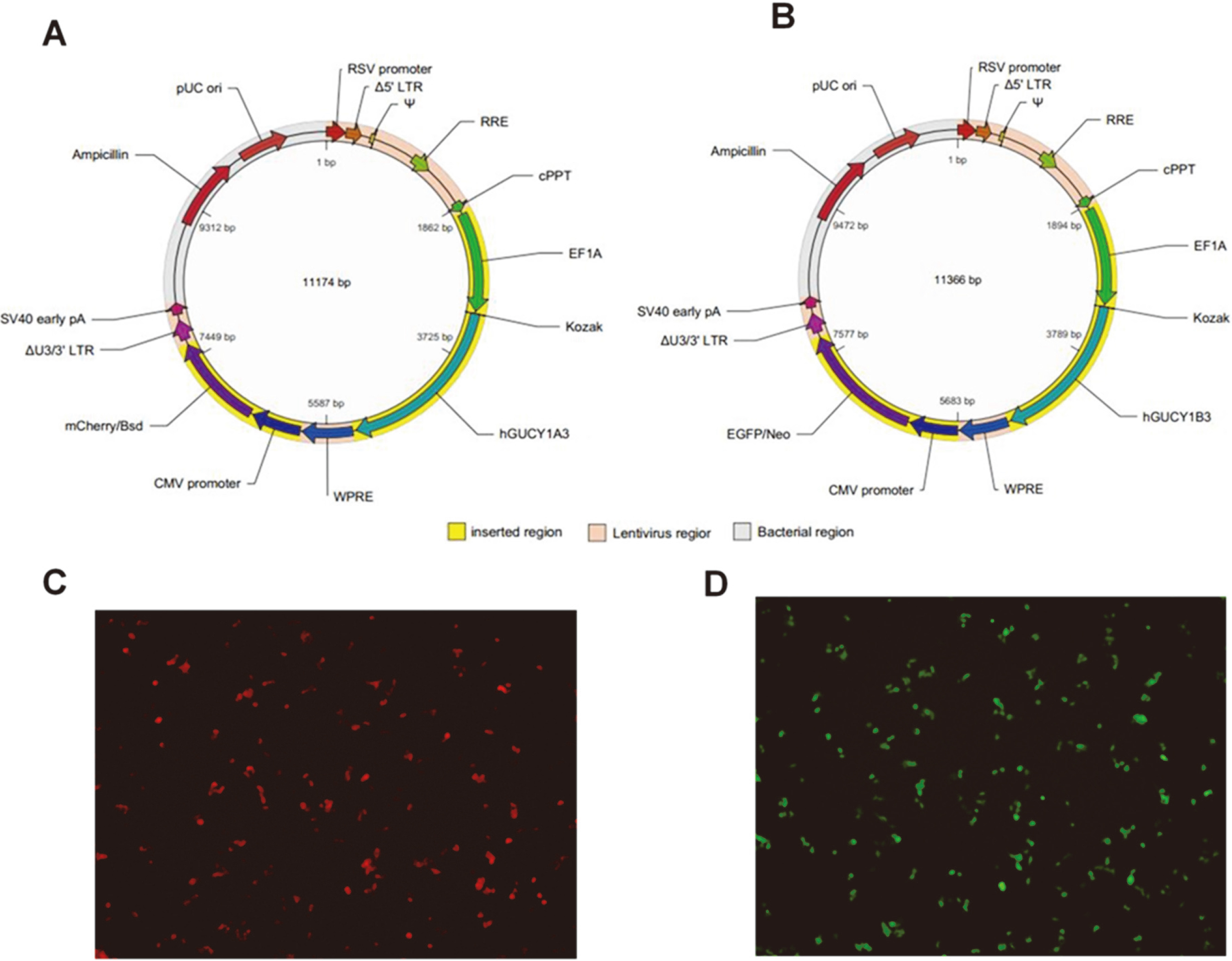
Lentiviral vectors expressing the human GUCY1A3 or GUCY1B3 genes were constructed (Fig. 1A and B). Cherry or green fluorescence was observed as expected in HEK293 cells 24 hours after transfection with corresponding lentiviral vectors (Fig. 1C and D). A minimal transfection rate of
We then examined expression of our target genes in the transduced HEK293 cells using primers specific for GUCY1A, GUCY1B3, PDE5A and the house keeping gene GAPDH by RT-PCR and real time RT-PCR (Fig. 2A and B). Comparing to control cells that were transduced with empty vector, co-transduction of GUCY1A3 and GUCY1B3 significantly increased expression of both the
Gene expression, enzyme activities and cellular cGMP levels after lentivirus transduction of the 
Gene expression and cellular cGMP levels after shRNA transduction in HEK293 cells. A: Lentivirus vector map of PDE5A shRNA. B: HEK293 cells exhibited green (EGFP) fluorescence after transduction. C: After transduction with lentivirus vectors harboring scramble or PDE5A shRNA, expression of the PDE5A gene was examined by RT-PCR. The expression of GAPDH served as an internal control. D: Cellular level of cGMP was also examined. SH1-SH3: three different designs of PDE5A shRNA; P: PDE5A; G: GAPDH. 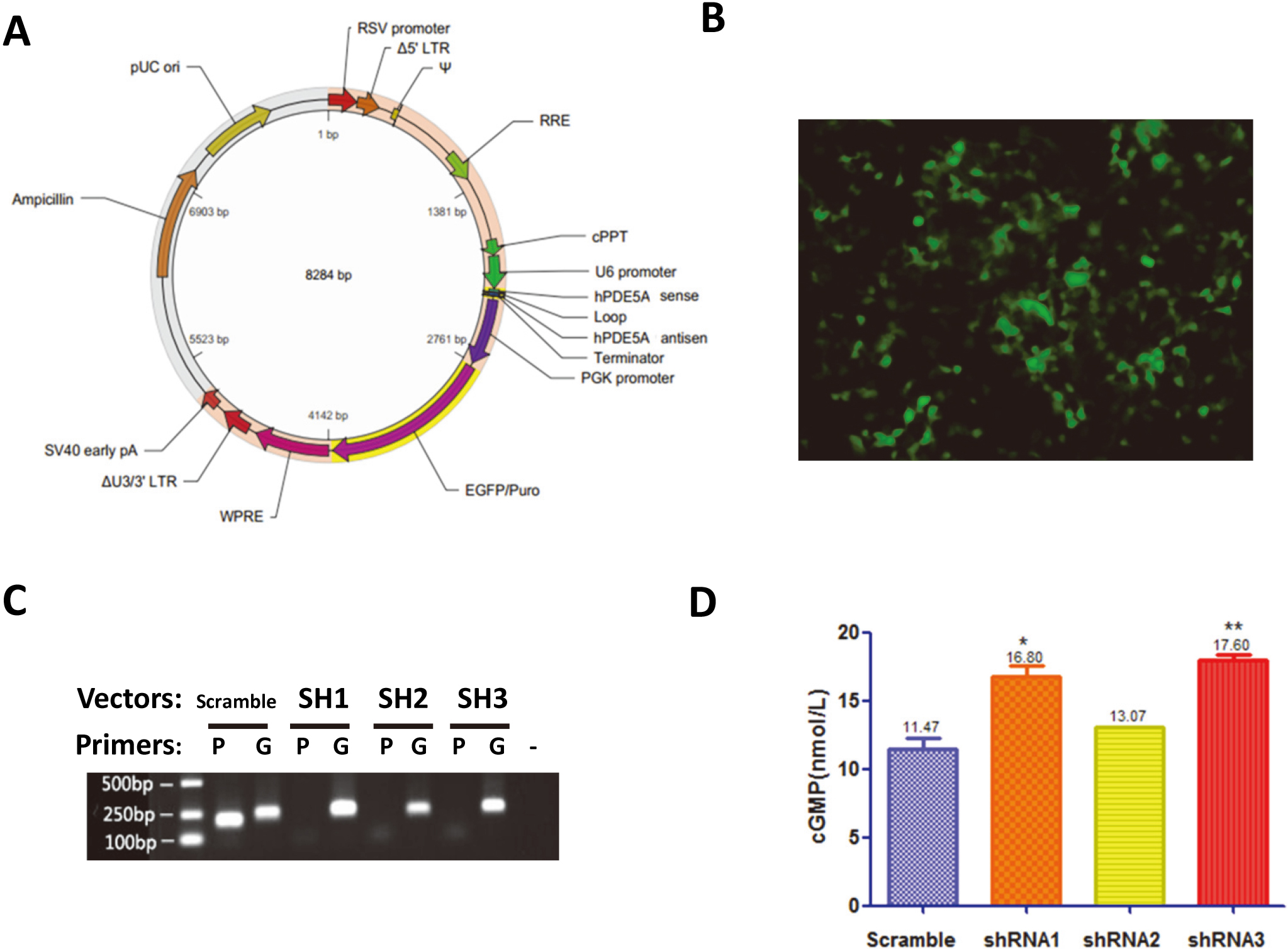
Cellular cGMP level after co-transduction of sGC subunits and PDE5A shRNA. 
To investigate whether the co-expressed
PDE5A shRNA decreased PDE5A expression and cellular cGMP level
Elevated level of cGMP may lead to the feedback up-regulation of PDE5A, a phosphodiesterase that specifically degrades cGMP in cardiomyocytes. To counter such potential feedback regulation on PDE5A, we designed PDE5A shRNA and investigated their effects in HEK293 cells (Fig. 3A). Transfections were successful for all constructs as indicated by the EGFP fluorescence observed in HEK293 cells 24 hours after transfection (Fig. 3B). As shown by the results of RT-PCR, the control cells exhibited strong expression of PDE5A, which were efficiently suppressed after transduction by any of the three shRNA constructs (Fig. 3C). Moreover, comparing to the control cells, down-regulation of PDE5A expression had led to an up to 53% significant increases in the cellular level of cGMP (
Co-Expression of GUCY1A3, GUCY1B3 and PDE5A shRNA in HEK293 cells increased cellular cGMP level collectively
We further investigated the synergetic effect of constructs. When co-expressed with GUCY1A3, GUCY1B3 and PDE5A shRNA, the cellular level of cGMP in HEK293 cells was slightly higher than cells transduced with GUCY1A3 and GUCY1B3 alone, but significantly higher than the control cells transduced with empty vector (
Discussion
NO-sGC-cGMP signaling and its cross talk with a variety of signal transduction molecules are important in many vital physiology processes. Numerous studies suggest that activation or enhancement of the pathway inhibits hypertrophy, decreases fibrosis, and protects against cardiac ischemia-reperfusion (I/R) injury [14, 15]. Although with great translational interests, treatments targeting the pathway have been limited to NO donors and PDE inhibitors that are not ideal. Stimulators or activators directly acting on the sGC are currently under development to avoid cytotoxicity, insufficiency or others negative side-effects of prolonged use of such agents [6]. Here, we report a genetic method that can moderately and stably elevate cGMP level in human cells, which underlies a potential gene therapy applicable to a range of cardiovascular diseases.
NO mediates its functions through its primary receptor sGC. Activated sGC converts GTP to cGMP and pyrophosphate (PPi). It has been well known now that cGMP acts as a ubiquitous second messenger in intracellular signaling cascades, which serves to regulate the activity of a number of downstream proteins [14, 16]. As a key player of the pathway, diminished expression and function of sGC has been shown to contribute to the pathogenesis of several cardiovascular disorders [17, 18]. Moreover, the chromosomal region harboring GUCY1A3 and GUCY1B3, which encodes the
The expression of sGC subunits is modulated at different levels, i.e. mRNA abundance, stability of mRNA and protein degradation [22, 23, 24, 25, 26]. Several studies reported that the regulation of steady-state sGC mRNA or function levels occurs in response to environmental stimuli such as oxidative stress and electric stimulation [6, 27]. Due to the clinical importance of targeting sGC activity, great efforts have been made on the development of haem-dependent sGC stimulators and haem-independent sGC activators [6]. On the other hand, no study has attempted to increase sGC level genetically in mammalian cells despite of the theoretical feasibility. We therefore co-expressed the
We constructed lentiviral vector harboring PDE5A shRNA to alleviate phosphodiesterase activity and cGMP degradation. Feedback upregulation of PDE5A expression by cGMP has been reported previously [11], but not observed in the current study. It is possible that the constructs we used only increase cGMP levels moderately (1.5–1.7 times) that was not sufficient to upregulate PDE5A. Nevertheless, expression of PDE5A shRNA in HEK293 cells resulted in decreased PDE5A expression and elevated cGMP level. HEK293 cells transduced with lentiviral vectors harboring PDE5A shRNA in combination with GUCY1A3 and GUCY1B3 exhibited the highest level of cGMP.
As a prototype of future molecular therapeutics, we have used lentiviral vectors that can infect diverse types of non-dividing cells with high efficiency. Use of lentiviral vectors to modulate gene expression has become standard practice in many areas of biomedical science [29]. Although the safety of lentiviral vectors has been largely improved, endogenous gene disruption is still a concern. Such off-target effects could be controlled by designing targeted insertion or by limiting the amount of transducing units used. To improve the safety of our construct, hypoxia responsive element (HRE) could be added to the promoter regions to allow conditional expression of the constructs only under conditions such as heart ischemia that lack of oxygen [30]. Evidence is emerging that the subcellular location of sGC and cGMP may be critical for their functions, although the underlying mechanisms are yet to be clarified [14, 16]. Such critical information should also be incorporated in our design of the constructs in the future.
Conclusion
Interventions targeting the NO-sGC-cGMP pathway are promising treatments for cardiovascular diseases such as heart ischemia. Genetic manipulation on the pathway has been rarely achieved, partly due to the complexity of the soluble guanylyl cyclase enzyme. We constructed lentiviral vectors harboring GUCY1A3 and GUCY1B3 that encode the
Footnotes
Conflict of interest
None to report.
Funding
This work was supported by grant 16JCYBJC42900 from the Natural Science Foundation of Tianjin City. The funding agency did not have a financial interest in the subject matter discussed in this manuscript.
Supplementary data
Cellular cGMP levels after treatment of 10 umol/L sodium nitroprussiate.