
Abstract
BACKGROUND:
Rituximab resistance is one of the great challenges in the treatment of diffuse large B-cell lymphoma (DLBCL), but relevant biomarkers and signalling pathways remain to be identified. Coptis chinensis and its active ingredients have antitumour effects; thus, the potential bioactive compounds and mechanisms through which Coptis chinensis acts against rituximab-resistant DLBCL are worth exploring.
OBJECTIVE:
To elucidate the core genes involved in rituximab-resistant DLBCL and the potential therapeutic targets of candidate monomers of Coptis chinensis.
METHODS:
Using the Traditional Chinese Medicine System Pharmacology Database and Analysis Platform (TCMSP), the Similarity Ensemble Approach and Swiss Target Prediction, the main ingredients and pharmacological targets of Coptis chinensis were identified through database searches. Through the overlap between the pharmacological targets of Coptis chinensis and the core targets of rituximab-resistant DLBCL, we identified the targets of Coptis chinensis against rituximab-resistant DLBCL and constructed an active compound-target interaction network. The targets and their corresponding active ingredients of Coptis chinensis against rituximab-resistant DLBCL were molecularly docked.
RESULTS:
Berberine, quercetin, epiberberine and palmatine, the active components of Coptis chinensis, have great potential for improving rituximab-resistant DLBCL via PIK3CG.
CONCLUSION:
This study revealed biomarkers and Coptis chinensis-associated molecular functions for rituximab-resistant DLBCL.
Introduction
Diffuse large B-cell lymphoma (DLBCL) accounts for 85% to 90% of all lymphomas [1]; 20% of patients are primary refractory (progressing during or immediately after treatment) to first-line standard therapy (R-CHOP), and 30% of patients relapse after achieving complete remission (CR) with R-CHOP [2]. The objective response rate (ORR) for patients with recurrent/refractory disease receiving second-line therapy was 26%, with a median overall survival of only 6.3 months [3]. Therefore, R-CHOP, as the first-line regimen, remains the core treatment for DLBCL. Patients with DLBCL who fail first-line therapy, especially those with primary refractory disease and early recurrence, have a poor prognosis and short survival. Rituximab resistance is an important cause of R-CHOP failure [4]. Approximately 30%–60% of indolent non-Hodgkin’s lymphoma (iNHL) patients who are not receiving rituximab treatment (including those who relapse or receive only chemotherapy) have certain drug resistance to rituximab [5]. Nearly 60% of DLBCL patients who were previously sensitive to rituximab therapy developed rituximab resistance at the time of retreatment [6]. Rituximab resistance is one of the great challenges in the treatment of DLBCL.
Progression during rituximab treatment or relapse within 6 months after treatment was considered rituximab resistance [7]. Downregulation, adjustment, mutation or loss of CD20 is an important mechanism of rituximab resistance. Acquired CD20 mutations were reported by Terui et al. These mutations diminish the mean fluorescence intensity of CD20, which may affect antibody binding and induce drug resistance [8]. Another mechanism of CD20 loss in rituximab-resistant tumour cells is known as “shaving”, or knockout. Binding of rituximab to CD20 results in the translocation of CD20 to lipid valves within the cell membrane, after which Fc
Workflow of the study.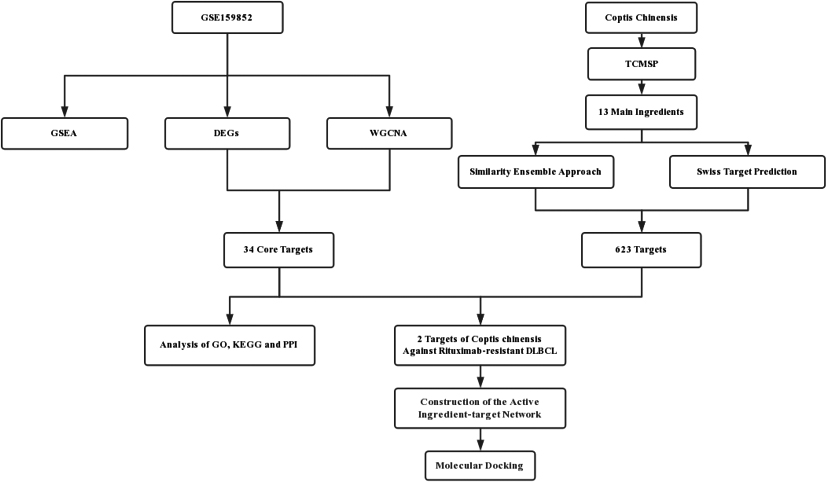
Rituximab resistance promoted the development of new CD20 monoclonal antibodies. Humanized anti-CD20 agents, including ofatumumab, IMMU106 (veltuzumab), and ocrelizumab, which are considered second-generation anti-CD20 agents and 3rd-generation anti-CD20 agents, include AME-133v (ocaratuzumab), PRO131921 and GA101 (obinutuzumab), respectively. These new monoclonal antibodies do not embed into lipid valves after binding to CD20 and induce high levels of ADCC and apoptosis without triggering CDC. However, these treatments have a limited effect on rituximab-resistant patients. In the GAUSS study, 175 iNHL patients who relapsed after treatment with rituximab regimens were randomized to receive either rituximab (375 mg/m2
Coptis chinensis and its active ingredients have been shown to have antitumour effects on many cancer species by inducing cell cycle arrest and apoptosis, inhibiting angiogenesis and inflammation, and suppressing invasion and metastasis [20]. Berberine, the most representative active ingredient of Coptis chinensis, eliminates DLBCL cells in vitro and in vivo by inhibiting c-myc expression, downregulating CD47 expression, and enhancing phagocytosis by macrophages [21]. In addition, berberine in combination with other chemotherapeutic agents has been reported to inhibit tumour progression. For example, the coadministration of berberine with evodiamine augmented the apoptosis of SMMC-7721 cells [22]. Berberine combined with cisplatin has a strong synergistic inhibitory effect on the growth of HeLa cells [23]. Coptis chinensis contains a variety of structural analogues of berberine, as well as a number of other components that have not been studied in depth; these compounds are potential antitumour agents with low price and few side effects. However, the impact of the monomeric components of Coptis chinensis on rituximab-resistant DLBCL has not been reported.
In this study, key genes associated with rituximab-resistant DLBCL were obtained by analysing differentially expressed genes (DEGs) and performing weighted gene coexpression network analysis (WGCNA). The biological functions and related signalling pathways of these genes were further determined by functional enrichment analysis, gene set enrichment analysis (GSEA), protein-protein interaction (PPI) network-based analysis, etc. Additionally, using network pharmacology and various online databases, we identified active targets of Coptis chinensis for the treatment of rituximab-resistant DLBCL and performed molecular docking to validate drug-target interactions, providing new references for the diagnosis and treatment of this disease. A schematic diagram of the integrated process is displayed in Fig. 1.
Bioinformatics analysis
Identification of DEGs
Gene profiles of patients with rituximab-resistant DLBCL were downloaded from the GEO database (
Coexpression network construction for WGCNA
First, we evaluated the variant genes to test their availability and used the R package WGCNA to construct a gene coexpression network. Then, we constructed an adjacency matrix to describe the correlation strength between the nodes. The matrix was constructed using the power function A_mn
Acquisition of core targets
Overlapping genes of DEGs and hub genes were obtained as core targets of rituximab-resistant DLBCL using Venny 2.1.0.
GSEA
GSEA software (version 3.0) was obtained from the following website: DOI:10.1073/pnas.0506580102,
Enrichment analyses and network visualization
The potential gene functional annotations and pathway enrichment associated with the common DEGs were elucidated. GO and KEGG analyses were performed using the clusterProfiler package of R software (version 3.10.1). The enrichplot and DOSE packages were used to visualize the enrichment results to aid interpretation.
Establishment of a protein-protein interaction (PPI) network
PPI data of the obtained core targets were extracted from the Search Tool for the Retrieval of Interacting Genes/Proteins (STRING,
Network pharmacology
Selection of monomers in Coptis chinensis
The main compounds of Coptis chinensis were retrieved from the TCMSP database (
Acquisition of pharmacological targets of Coptis chinensis
The selected main components of Coptis chinensis were input into the Similarity Ensemble Approach (
Identifying targets of Coptis chinensis against rituximab-resistant DLBCL
The core targets of rituximab-resistant DLBCL were intersected with the pharmacological targets of Coptis chinensis to identify potential targets against rituximab-resistant DLBCL. The APN was constructed and visualized via Cytoscape software (version 3.9.1).
Molecular docking analysis
Molecular docking analysis was performed using AutoDock Vina software (version 1.5.7). The molecular structures of the monomers in Coptis chinensis were obtained from the TCMSP database. The protein structures were acquired from the PDB database (
Analysis of core targets on rituximab-resistant DLBCL. (a) Volcano map of DEGs related to rituximab-resistant DLBCL. (b) Heat map of DEGs related to rituximab-resistant DLBCL. (c) Cluster dendrogram of the co-expression network modules (1-TOM). (d) Cluster plot analysis of the relationship between modules. (e) Heatmap of the correlation between the module eigengenes and clinical traits of DLBCL. (f) Scatter plot analysis of the antiquewhite2 module. (g) Venn diagram depicting the overlap of DEGs and hub genes from WGCNA.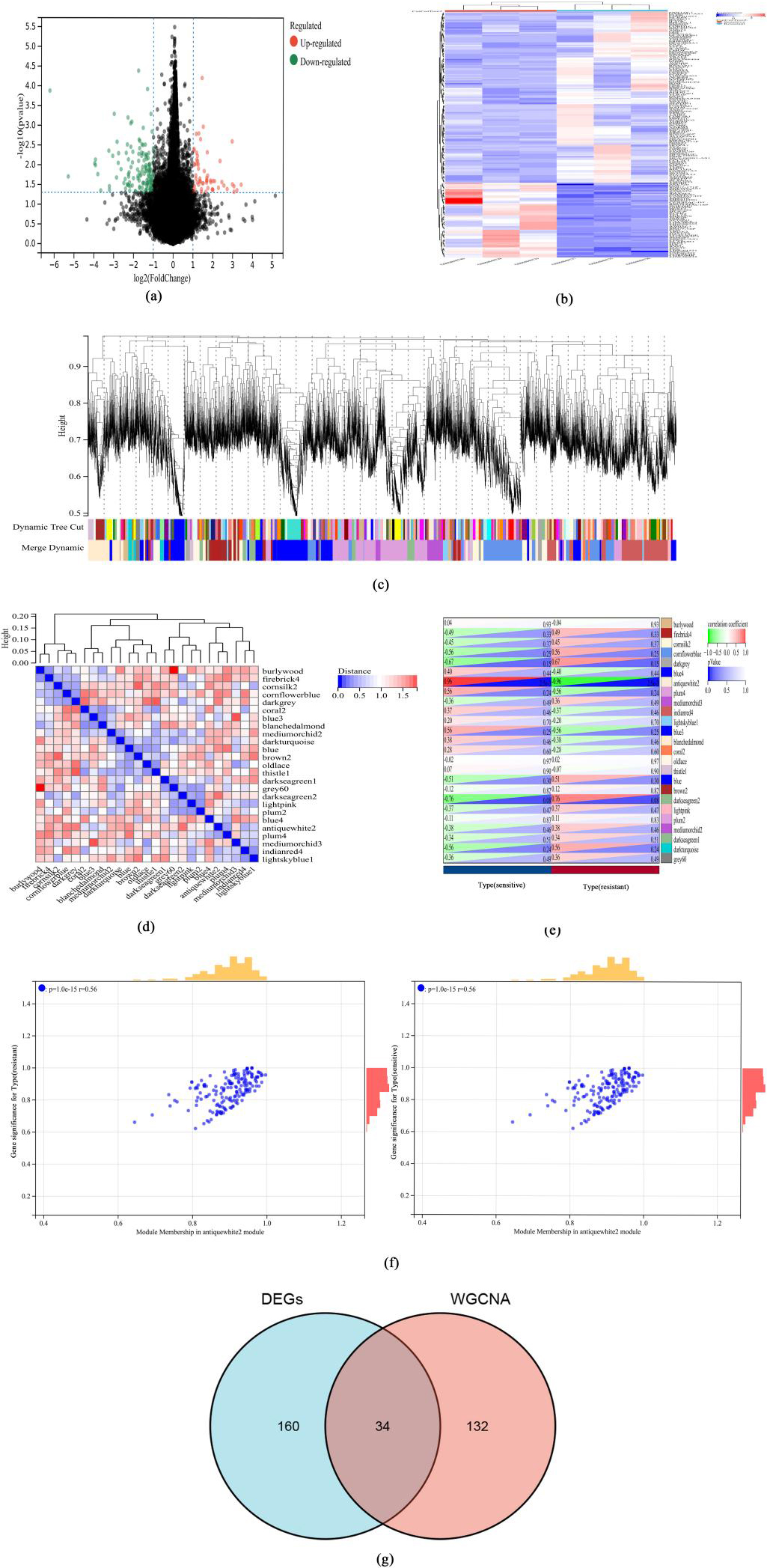
Gene Set Enrichment Analysis. (a) Biological process pathways. (b) Molecular function pathways. (c) KEGG pathways.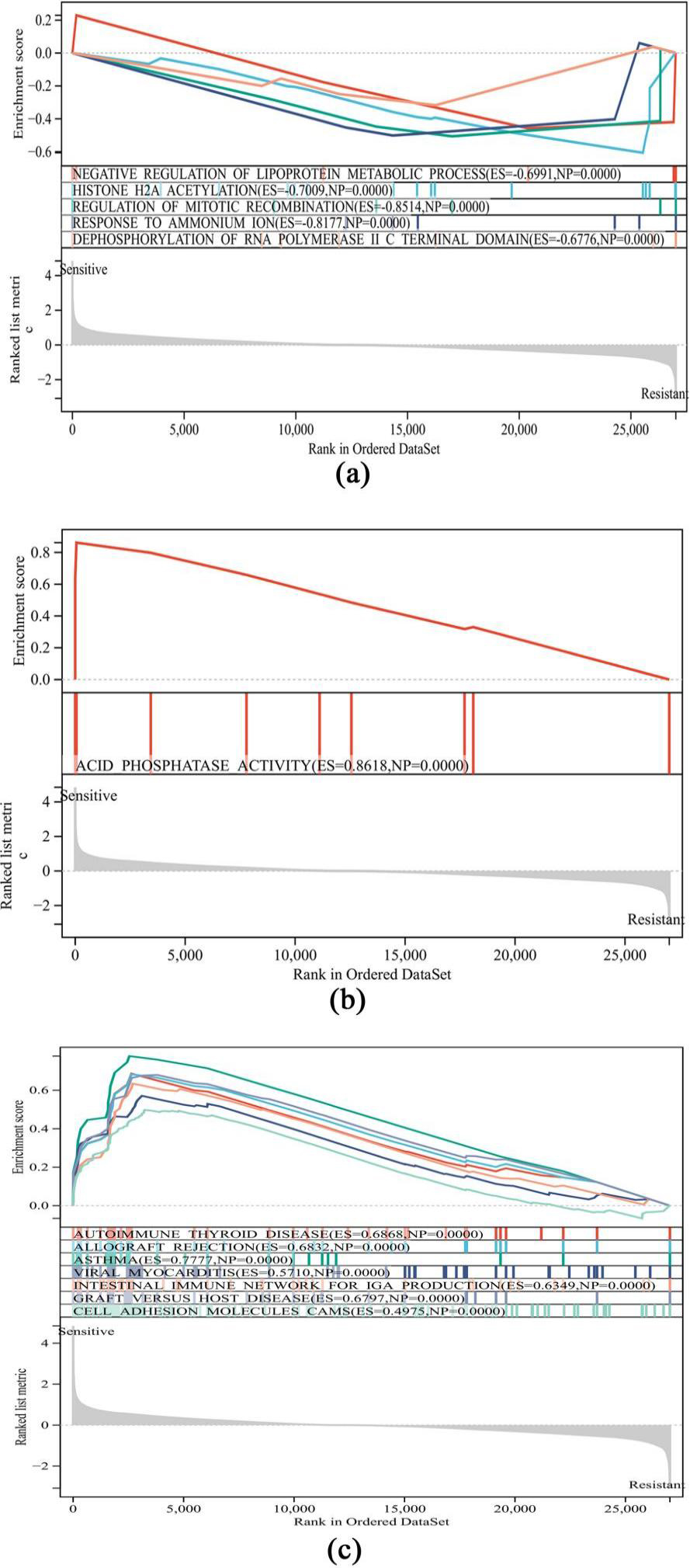
Identification of the core targets of rituximab-resistant DLBCL
A total of 194 DEGs related to Rituximab-resistant DLBCL were identified (Fig. 2a), of which 59 genes were upregulated and 135 were downregulated (Fig. 2b). For WGCNA, a total of 25 modules were identified using the dynamic tree cut package (Fig. 2c and d). We determined the correlations between the modules and the occurrence of rituximab-resistant DLBCL by establishing a module-trait relationship. The antiquewhite2 module (Fig. 2e) was significantly correlated with the occurrence of rituximab-resistant or sensitive DLBCL (coefficient of
GSEA
GSEA was also conducted between rituximab-resistant and rituximab-sensitive DLBCL patients to explore the possible pathways and gene sets involved. The results revealed 5 GO-biological process (BP) pathways, 1 GO-molecular function (MF) pathway and 7 KEGG pathways. The GO-BP pathways were associated with N-negative regulation of the lipoprotein metabolic process, histone H2A acetylation, regulation of mitotic recombination, response to ammonium ions and dephosphorylation of the RNA polymerase IIC terminal domain (Fig. 3a). Only the acid phosphatase activity pathway in the GO-MF was related to rituximab resistance (Fig. 3b). The representative pathways identified by KEGG analysis included autoimmune thyroid disease, allograft rejection, and asthma (Fig. 3c).
Functional enrichment and PPI network of the core targets
The 34 core genes were subjected to GO and KEGG enrichment analyses. GO enrichment analysis revealed that genes involved in the positive regulation of alkaline phosphatase activity, antigen receptor-mediated signalling pathway, immune response-activating cell surface receptor signalling pathway and other pathways were involved (Fig. 4a). Additionally, in the KEGG pathway analysis, the top 10 pathways were found to be involved in cell adhesion molecule (CAM) metabolism, viral myocarditis and inflammatory bowel disease (IBD) (Fig. 4b). Next, STRING analysis was used to determine the PPIs mediated by the 34 core genes (Fig. 4c), and the 5 genes associated with the genes according to degree were RAC2, MS4A1, PIK3CG, EZR and EVI2B.
Screening of active ingredients and related pharmacological target genes
According to the TCMSP database, Coptis chinensis contains 48 ingredients in total. The patients were screened for OB
Main compounds of Coptis chinensis
Main compounds of Coptis chinensis
Functional enrichment and PPI network of the core genes. (a) Top 10 results of GO enrichment analysis. (b) Top 10 results of KEGG enrichment analysis. (c) Analysis results of PPI network.
Coptis chinensis-associated molecular functions for rituximab-resistant DLBCL. (a) Venn diagram depicting the overlap of the core genes and the pharmacological targets. (b) Active ingredient-target network. (c) Molecular docking of berberine, quercetin, epiberberine and palmatine to PI3K
As shown in Fig. 5a, 2 intersection genes, PIK3CG and EZR, were potential targets of Coptis chinensis against rituximab-resistant DLBCL. The targeting relationship between Coptis chinensis and the intersection genes was determined by the active ingredient-target network (Fig. 5b). PIK3CG was the gene associated with the highest number of active components and included berberine, quercetin, epiberberine and palmatine. The corresponding ingredient of EZR was berberrubine. The interaction between the active ingredients and targets was validated using molecular docking analysis. The results illustrated that phosphoinositide 3-kinase subunit gamma (PI3K
Molecular docking information on active ingredient-target of Coptis chinensis against rituximab-resistant DLBCL
Molecular docking information on active ingredient-target of Coptis chinensis against rituximab-resistant DLBCL
Rituximab, a targeted monoclonal antibody against CD20, is effective at prolonging survival in DLBCL patients and has become a standard component of DLBCL treatment. However, the main obstacle to the efficacy of RTX is its resistance. Therefore, it is highly clinically important to potentiate the R-CHOP regimen by investigating the mechanisms of rituximab resistance and identifying drugs that may block the relevant mechanisms. This study is the first to identify core targets in rituximab-resistant DLBCL patients using differentially expressed gene (DEG) analysis, weighted gene coexpression network analysis (WGCNA) and other bioinformatics techniques and reveal that berberine, quercetin, epiberberine and palmatine may bind to Coptis chinensis through the core gene PIK3CG via network pharmacology and molecular docking analysis.
Three of the 34 core targets of rituximab-resistant DLBCL, CD99, RAC2, and MS4A1, were reported to be associated with the efficacy or adverse effects of the R-CHOP regimen. Junshik Hong et al. reported that, in DLBCL patients receiving the R-CHOP regimen, 2-year event-free survival (EFS) and overall survival (OS) were better in CD99
GSEA of all genes, as well as GO and KEGG enrichment analyses of 34 crossover genes, suggested that phosphatase activity, cell adhesion molecules (CAMs), viral myocarditis, asthma, and allograft rejection play important roles in rituximab-resistant DLBCL. The level of alkaline phosphatase (ALP) was reported by Ching-Qing Cai et al. to be a new independent predictor of central nervous system relapse in DLBCL patients receiving the R-CHOP regimen [27]. In CAMs, cellular experiments have shown that knockdown of ICAM-1 can cause rituximab resistance [28]. There have also been reports about severe and fatal enterovirus-associated myocarditis after the use of rituximab [29, 30] and, conversely, the efficacy of rituximab for different types of asthma [31, 32] and renal transplant patients [33].
This network pharmacology study revealed that EZR and PIK3CG are potential key genes involved in the inhibition of rituximab resistance in Coptis chinensis. Ezrin, the coding product of EZR, is expressed in the stomach, kidney and intestine [34]. It is usually located in cell folds, cell pseudopodia, microvilli, cleavage grooves and contraction rings and participates in the regulation of cell adhesion [35]. EZR is an important factor related to tumour invasion [36]. PIK3CG encodes PI3K
The candidate active ingredients in Coptis chinensis that block rituximab resistance are berberine, quercetin, epiberberine, and palmatine. There are no reports of epiberberine or palmatine combined with rituximab or DLBCL. Berberine exerts antitumour effects on DLBCL by modulating the c-myc/CD47 axis, but its effect on rituximab-resistant DLBCL is unclear [21]. CD47 enhances tumour cell viability and migration via activation of the PI3K/Akt pathway [39, 40]. Xin Li et al. reported that quercetin enhances the apoptosis of DLBCL cells through the inhibition of the STAT3 pathway [41]. In DLBCL, the activation of STAT3 and its kinase JAK1 is caused by the autocrine production of IL-6 and IL-10 in the activated B-cell-like subtype (ABC). STAT3 regulates multiple oncogenic signalling pathways, including the NF-
Our study has several limitations. First, the small sample size (3 sensitive and 3 resistant samples) may have affected the accuracy of the results. Second, bioinformatics and network pharmacology have challenges such as inherent variability and poor reproducibility of biological data; thus, the results need further validation at the animal or cellular level. Finally, further preclinical studies and clinical trials are needed to support the use of Coptis chinensis for the treatment of rituximab-resistant DLBCL.
Conclusion
In this study, we revealed potential efficacy predictive biomarkers in DLBCL patients treated with rituximab. Berberine, quercetin, epiberberine, and palmatine, which target PIK3CG, may be viable options for patients with rituximab-resistant DLBCL. Although further pilot studies are needed to determine the true clinical potential of these findings, our findings could lead to a new solution for treating rituximab resistance in DLBCL or other diseases.
Ethics statement
The data used in this study were obtained from public databases. No ethics approval was required for this study.
Funding
This study was supported by the Natural Science Foundation of Fujian Province (Grant No. 2023J011276), the Fujian Provincial Health Technology Project (Grant No. 2022CXA027) and Fujian Cancer Hospital Project (Grant No. 2023YN11).
Author contributions
Qiuling Zhao and Shengqiang Huang performed this study and contributed to the R language programming, acquisition, and interpretation of the data visualizing tasks. Lin Yang created the tables and figures. Ting Chen performed the molecular docking. Xiuliang Qiu modified the formatting procedure. Ruyi Huang and Liangliang Dong drafted and edited the manuscript. Wenbin Liu reviewed the manuscript. All authors read and approved the final manuscript.
Availability of data and materials
All data can be obtained from the corresponding author upon request.
Footnotes
Acknowledgments
Not applicable.
Conflict of interest
The authors declare that they have no competing interests.