
Abstract
Locomotor steering is contingent upon orienting oneself spatially in the environment. When the head is turned while walking, the optic flow projected onto the retina is a complex pattern comprising of a translational and a rotational component. We have created a unique paradigm to simulate different optic flows in a virtual environment. We hypothesized that non-visual (vestibular and somatosensory) cues are required for proper control of a straight trajectory while walking. This research study included 9 healthy young subjects walking in a large physical space (40×25m2) while the virtual environment is viewed in a helmet-mounted display. They were instructed to walk straight in the physical world while being exposed to three conditions: (1) self-initiated active head turns (AHT: 40° right, left, or none); (2) visually simulated head turns (SHT); and (3) visually simulated head turns with no target element (SHT_NT). Conditions 1 and 2 involved an eye-level target which subjects were instructed to fixate, whereas condition 3 was similar to condition 2 but with no target. Identical retinal flow patterns were present in the AHT and SHT conditions whereas non-visual cues differed in that a head rotation was sensed only in AHT but not in SHT. Body motions were captured by a 12-camera Vicon system. Horizontal orientations of the head and body segments, as well as the trajectory of the body’s centre of mass were analyzed. SHT and SNT_NT yielded similar results. Heading and body segment orientations changed in the direction opposite to the head turns in SHT conditions. Heading remained unchanged across head turn directions in AHT. Results suggest that non-visual information is used in the control of heading while being exposed to changing rotational optic flows. The small magnitude of the changes in SHT conditions suggests that the CNS can re-weight relevant sources of information to minimize heading errors in the presence of sensory conflicts.
Locomotion is an essential activity of our daily lives. In order to accomplish the desired locomotor task, one must be able to propel the body forward while maintaining postural equilibrium and simultaneously adapting to changes in environmental conditions. Heading can be defined as ‘the compass direction in which the longitudinal axis of a ship or aircraft points’ [18]. Locomotor heading in particular can be represented as the movement direction (angular orientation) of the subjects’ body centre of mass (CoM) or its tangential velocity projected onto the horizontal plane [10, 11]. Such spatial orientation is detected by visual, vestibular and proprioceptive cues combined to generate an internal representation of self-motion in the context of the external environment. In particular, the multi-sensory fusion [6] of linear and angular head acceleration signals from the vestibular system, as well as proprioceptive information from the occulomotor and head-neck systems and body somatosensation [1, 29] leads to a single representation of self-motion and orientation in space that allows for functional motor actions. Motor efference also appears to play a role in the judgment of heading direction during eye or head or gaze rotations [3, 27]. An efference copy of the oculomotor command was suggested as a means to reduce the gain of the vestibulo-ocular reflex during gaze redirection [22]. When an active eye/head rotation is performed while walking, it is not only the visual and non-visual information induced by the head/eye movements, but also the sensorimotor information associated with locomotion, that has to be processed for interpretation of heading direction. Such complex sensorimotor integration appears to be processed smoothly by the intact central nervous system (CNS), as healthy young individuals show little deviation from the desired heading direction when executing active head turns while walking [2, 27]. However, persons with a central nervous system disorder (such as stroke) can experience deviations in their walking trajectory when executing voluntary head turns while walking, possibly due to a defective integration of sensory information along with the motor command [12].
Vision is a powerful component which provides information about the environment and self-motion during locomotion [21]. One important component of the visual signal is optic flow, broadly defined as the pattern of visual motion projected on the retina during self- or environmental motion, which allows an individual to modulate their direction and speed of walking, based on visual feedback [7]. When someone travels in a linear path, a radial pattern of optic flow is experienced, with a focus of expansion (FOE) originating from the direction of heading [31]. However, it often happens that, as one turns the head or eyes while walking, the flow information being presented at the retina (also referred to as retinal flow) is no longer uniquely linear, but is also comprised of a rotational component. This results in a shifted pseudo-FOE which no longer corresponds to the direction of heading [16, 26]. This combination of translational and rotational optic flow creates a visual pattern that can either depict a rotating head/eye while walking forward, or be interpreted as walking along a curved path [1, 28].
Two main theories have been proposed to explain how an individual discriminates between rotating head/eye movements and curved paths. The first theory suggests that the retinal optic flow in itself contains enough information to distinguish between rotational and translational flows, and hence properly signal heading direction. Some reports state that this theory is supported under conditions involving slow rotation rates or the presence of dense motion parallax and reference objects [1, 30], but accurate heading judgments and adjustments from optic flow were also reported for rotation rates as high as 16°/s to 20°/s, independent of the presence of other visual cues [26, 16]. The second theory suggests that non-visual (extra-retinal) information is used to dissociate the rotational and translational components from the retinal flow. With this additional extra-retinal information, one makes more accurate judgment of heading direction than without [1, 24]. To date, it is uncertain as to what extent those psychophysical theories, which are based mainly on experimental data collected in a static and/or sitting position, also apply to locomotion. We have previously shown that subjects deviate from their initial walking trajectory when exposed to rotational flows externally induced through a virtual reality display during locomotion [25]. Although the deviations were smaller than those experienced and expected with mediolateral translational flows [25, 31], subjects exposed to rotational flows were, at least partially, correcting for a perceived change in walking trajectory. In those studies, however, subjects were instructed to control heading from optic flow in the virtual environment’s frame of reference. This tells us little about how one uses different sources of sensorimotor information to control a straight locomotor trajectory in the physical or real dimension while exposed to rotational optic flows induced by active (or simulated) head rotations.
The question addressed in this study was whether rotational optic flows generated internally (induced by active head turns) or externally (through visually simulated head rotations) would induce different effects on the control of straight locomotor trajectory. For this purpose, we compared the walking trajectory and body segment orientations when exposed to rotational flows induced by active versus visually simulated head rotations. We hypothesized that the rotational flow generated externally by simulated head rotations would lead to deviations in the body’s centre of mass (CoM) trajectory and body segment orientations, as compared to active head rotations that could maintain a straight walking trajectory with no body segment orientation changes.
Experimental procedures
Subjects
Nine young healthy adults, 4 males and 5 females (age: 23.3±3.8 years, height: 167.8±5.9 cm, weight: 65.2±9.2 Kg), volunteered to participate in this study conducted in the Virtual Reality and Mobility Laboratory at the Jewish Rehabilitation Hospital (Centre Intégré de Santé et Services Sociaux de Laval). Subjects were free of orthopaedic, neurological or visual problems that may have interfered with locomotion. Written informed consent approved by the IRB of the Centre for Interdisciplinary Research in Rehabilitation of Greater Montreal was given by all participants.
Instrumentation
Three-dimensional body kinematics were acquired at 120 Hz using a Vicon-512 motion analysis system with 10 high-resolution (1000×1000 pixels) cameras (M60). The virtual environment (VE) was displayed in an NVisor helmet-mounted display (HMD) and the field of view of the HMD was 47° horizontal and 37° vertical (60° diagonal). The HMD had 3 markers attached to it (front, left and right), which formed a rigid body representing the head, allowing the Vicon system to calculate head position and orientation (pitch, roll, and yaw) in 3 dimensions. The virtual reality system used was generated and controlled by CAREN-2 (Computer Assisted Rehabilitation ENvironments, by Motek Medical BV), which is a closed-loop system which has the ability to synchronize the view of the virtual room based on the location and orientation of the head [25]. A real-time control algorithm allowed for the VE displayed in the HMD to be updated in real time as a function of the subjects’ head displacement.
Experimental set-up and protocol
Anthropometric measurements were recorded and thirty-nine reflective markers were placed on specific body landmarks in order to create a full body marker set, as specified by the Vicon Plug-in Gait model. Subjects were fitted with the HMD, which allowed them to view the VE consisting of a large, rich-textured (40 m×25 m) virtual room with pillars located at its end [25]. Head coordinates were fed real-time through the Tarsus engine from Vicon to the VR (Caren-2) system that controlled the VE.
Subjects were first habituated and familiarized to walking in the VE before any data were collected. The subjects were randomly exposed to rotational flows with pseudo-FOEs shifting from 0° to 40° to the left (–40°) or right (+40°), or no flow rotation (0°). The rotational flows were triggered either externally (visually simulated head turn: SHT) or by having the subject rotate their head (active head turn: AHT). In the SHT condition, the virtual environment was rotated while the subject was walking and fixating on a red stationary target located 2 m straight ahead, at eye level (Fig. 1). Note that in this condition, the target remained fixed and centered with respect to the HMD screen. In the AHT condition, subjects were walking while fixating a target of similar characteristics rotating from 0° to 40° (Fig. 1). In order to verify whether the presence of a target had an effect on the steering behaviour, the simulated condition was also performed without a target (SHT_NT).
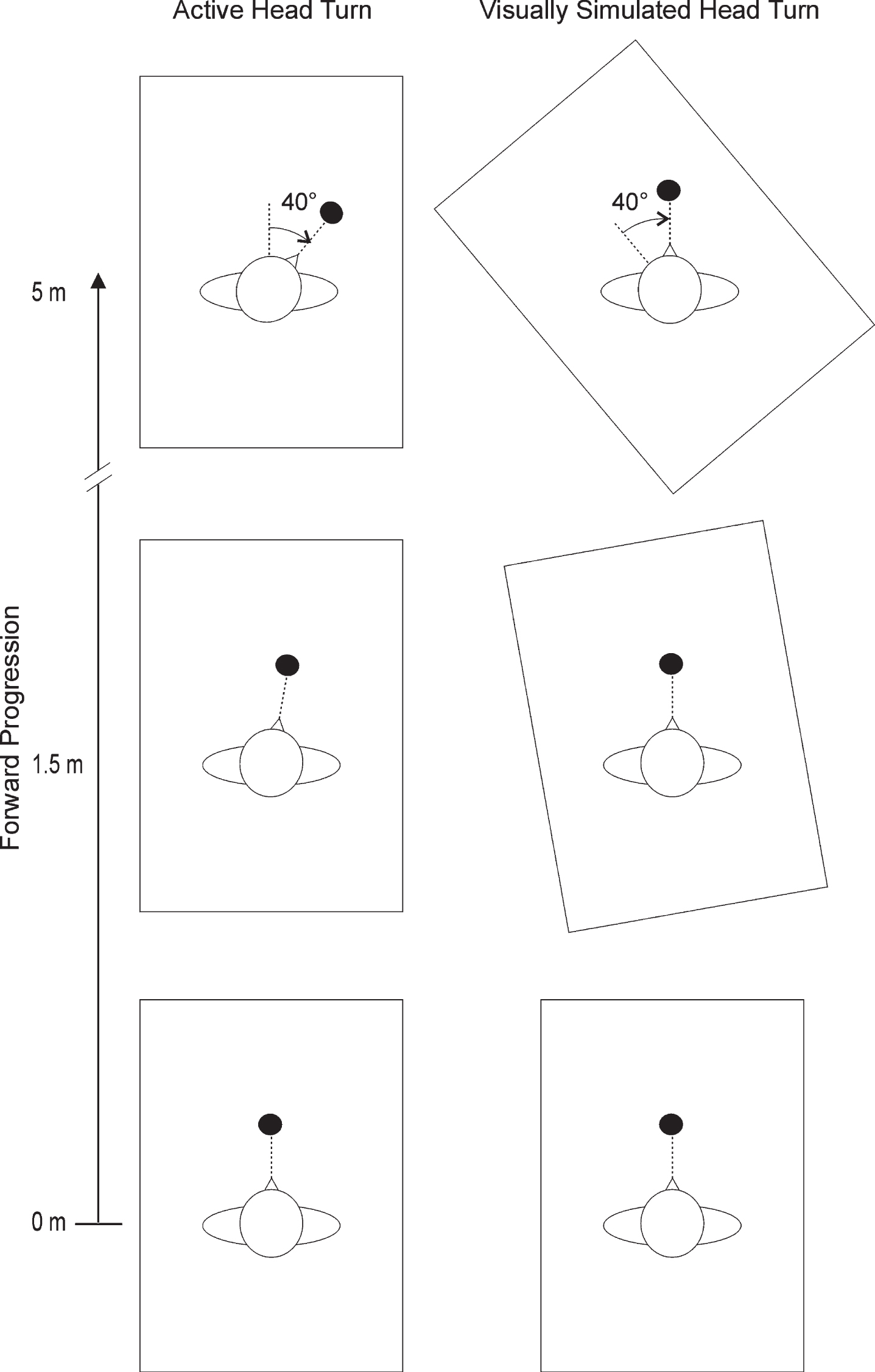
Schematic bird’s eye view of an active (left panel) and a visually simulated (right panel) head turn to the right. Subjects were walking in a virtual environment (represented by rectangular box) while fixating on a round target (represented by black dot) located at 2 m in front of them. At 1.5 m of forward walking, the target (active head turn condition) or the scene (simulated head turn) started to rotate, until it reached a final location of 40° at 5 m of forward walking. Note that the virtual room (40 m×25 m) and target location represented are not drawn to scale. The simulated head turn condition with no target is not shown.
Subjects were instructed to walk straight in the physical world, as opposed to the virtual world being viewed in the HMD, and to maintain the red target in the centre of the HMD screen. After 1.5 m of forward walking, the optic flow began to be rotated due to the AHT or SHT. That rotation lasted over 3.5 m, such that after 5 m of forward walking, the rotational flow experienced by the subjects was±40° or remained at 0°. Each direction (–40°, 0°, +40°) was repeated four times across the three conditions (AHT, SHT, SHT_NT) for a total of 36 trials that were block-randomized for each subject. The main outcome measure retained for analysis was heading orientation, defined as the mean instantaneous angular deviation of CoM trajectory in the horizontal plane. Secondary outcome measures included CoM mediolateral deviation as well as horizontal orientation of the head, thorax, pelvis and left foot. Data were examined in the physical world, which represented the coordinate system of interest, and averaged over the distance comprised between 4.75 m and 5 m of forward walking. Virtual head rotation was also examined to ensure that the perceived head rotation (stimulus) was identical across conditions. Data were thereafter averaged across trial conditions and across subjects.
Repeated measures 2-way analyses of variance (ANOVA) were performed to assess main or interaction effects due to head turn conditions (AHT, SHT and SHT_NT) and final pseudo-FOE location (–40°, 0°, +40°). A p-value <0.01 was accepted as significant for main and interaction effects, after adjustment with the Bonferonni test. Posthoc comparisons were made with Tukey’s test of pairwise comparisons. Statistics were performed in Statistica 7.1.
Results
In all three head turn conditions, the instruction was to walk straight in the physical world while being presented with a rotational flow pattern, whether simulated or actively induced. A perfect maintenance of a straight heading direction would result in no deviation of CoM trajectory, as well as no segment reorientation in the horizontal plane in the physical world (other than when an active head turn was present). Figure 2 illustrates examples of CoM trajectories and head reorientation patterns across head turn conditions and pseudo-FOE locations. Subtle changes in CoM trajectory were observed when the flow was rotated from the neutral (0°) condition. In the example illustrated, the subject veered slightly in the direction of the pseudo-FOE shift in the AHT condition, but away from the pseudo-FOE location in SHT and SHT_NT conditions. CoM trajectories were also identical between the physical and virtual worlds, indicating a rotation around the subject’s head without any translation. For head rotations, the difference across conditions in the physical world accounts for the fact that an active head turn was performed in the AHT condition, while it was visually simulated in the SHT and SHT_NT conditions. In the virtual world, head orientation traces were similar across optic flow conditions, indicating that subjects were exposed to similar amounts of perceived head rotation.

Traces of individual trials for all three final locations (40° left, 0°, 40° right) of focus expansion (pseudo-FOE) for the (A) active head turn (AHT), (B) visually simulated head turn (SHT) and (C) visually simulated head turn with no target (SHT_NT) conditions. For each condition, mediolateral (ML) displacement of body centre of mass (CoM) (left panels) and head yaw (right panels) are illustrated as a function of CoM anteroposterior (AP) displacement, both in the physical (upper graphs) and virtual (lower graphs) worlds.
In the physical world, significant main effects due to pseudo-FOE shifts and interaction effects between head turn conditions and pseudo-FOE shifts were found for CoM heading orientation (Fig. 3), as well as most body segment horizontal orientations (Fig. 4). On average, CoM heading orientation deviated away from the pseudo-FOE location for the SHT and SHT_NT conditions, but not the AHT condition. While no significant differences in CoM heading orientation emerged for any directions of rotational flow between the different head turn conditions, a differential effect of pseudo-FOE location was observed in the simulated head turn conditions only, as CoM heading orientation was found to differ between the left (–40°) vs. right (+40°) pseudo-FOE locations (p < 0.01 to p < 0.05). Similar results were obtained for CoM mediolateral deviation.

Heading errors and CoM mediolateral (ML) displacement for all three final locations (40° left, 0°, 40° right) of focus expansion (pseudo-FOE) and for the active head turn (AHT), the visually simulated head turn (SHT) and the visually simulated head turn with no target (SHT_NT) conditions. Mean (±1SE) values are represented in the physical world coordinates. Statistically significant pairwise comparisons are indicated, with p < 0.05 (*), p < 0.01 (**).
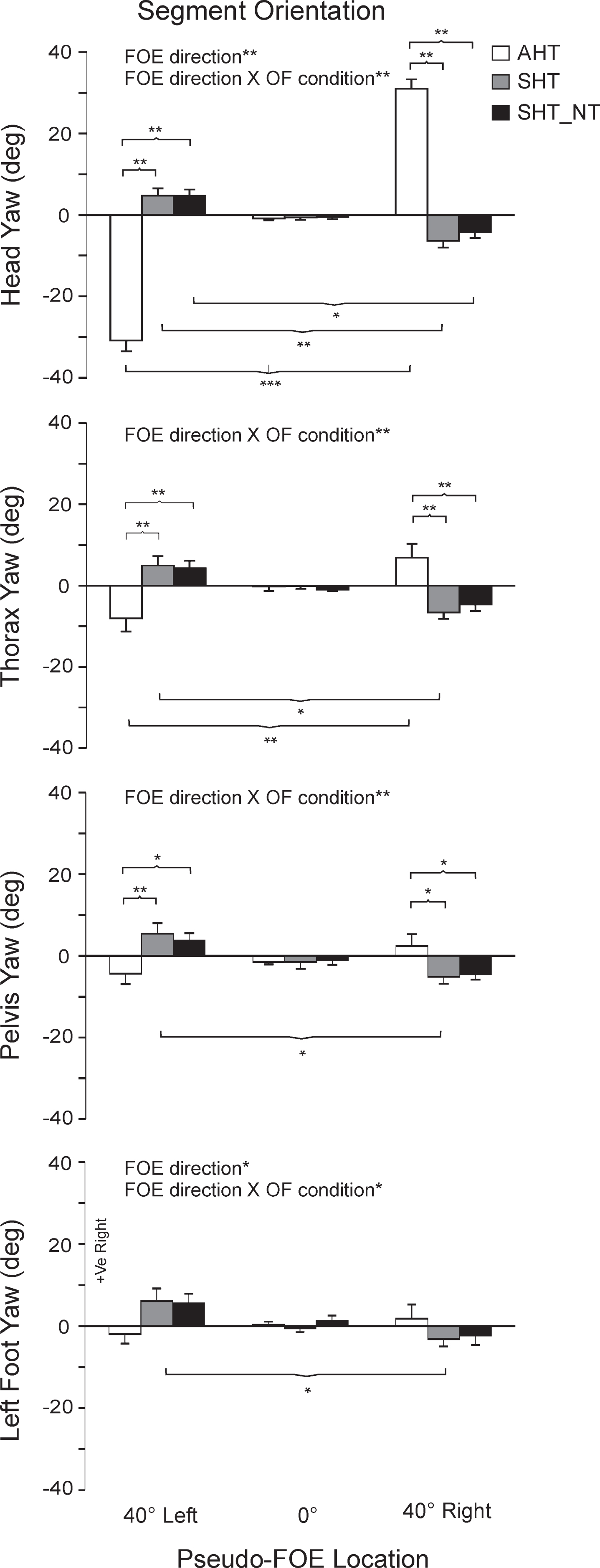
Mean (±1SE) values of body segment orientations for all three final locations (40° left, 0°, 40° right) of focus expansion (pseudo-FOE) for the active head turn (AHT), the visually simulated head turn (SHT) and the visually simulated head turn with no target (SHT_NT) conditions. Data are represented in the physical world. Statistically significant pairwise comparisons are indicated, with p < 0.05 (*), p < 0.01 (**), p < 0.001 (***).
The head was reoriented in the direction of the active head turn in the AHT condition while a small reorientation (typically <8°) in the direction of CoM heading reorientation was systematically observed for the SHT and SHT_NT conditions (Fig. 4). The head reorientation pattern differed across head turn conditions when the pseudo-FOE was shifted (p < 0.001). Furthermore, head reorientation differed across all pseudo-FOE locations for the AHT condition (p < 0.0001), and between the right vs. left pseudo-FOE locations (p < 0.01 to p < 0.05) for the visually simulated head turns (SHT, SHT_NT). Thorax, pelvis and foot orientation displayed reorientation patterns similar to the head, although the magnitude of this reorientation became attenuated in more distal (lower) segments (e.g. the foot) (see detailed pairwise comparisons, Fig. 4). Pelvis and foot segment orientations remained similar across pseudo-FOE locations for the AHT condition and were only found to differ (p < 0.05) between left (–40°) vs. right (+40°) pseudo-FOE shifts for the SHT condition.
In order to verify whether the perceived head turn rotation (stimulus) was comparable across conditions, the amplitude and rate of virtual head rotations were also analyzed. As expected, there was no significant difference in virtual head rotation amplitude (p > 0.88) regardless of active or simulated head turn conditions. Note that the average head turn amplitude did not reach the full 40°, by 5° to 8°. Mean rotational optic flow rate induced by the head rotations, which was dependent on gait speed (1.0±0.1 m/s), was found to be 11.0±1.3°/s (mean±1SD, for all conditions and pseudo FOE¬ =0). Similar optic flow rates were observed for the AHT (mean±1SD: 10.5±1.6 deg/sec), SHT_NT (11.3±1.6 deg/sec) and SHT conditions (11.1±1.3 deg/sec).
We have created a unique paradigm which includes active and visually simulated head turns to investigate the multisensory control of straight locomotor trajectory by comparing the heading and body segment orientations in response to rotational optic flow generated internally vs. externally during walking. Our results revealed an interaction effect between the direction of pseudo-FOE shifts and head turn condition for heading orientation, such that the rotational optic flow caused the subjects to veer in the direction opposite to the head turns only when those head turns were simulated, i.e. externally generated rotational flow in the absence of actual head motions. Likewise, head, thorax and pelvis displayed small but systematic reorientations in the direction opposite to the pseudo-FOE shifts during the visually simulated head turns, which is consistent with the presence of a steering response [25]. When a horizontal rotational flow is combined with an expanding translational flow, as in the present study, the resulting flow can be perceived either as a head rotation while walking or as walking along a curved path [1, 28], the latter implying a change of trajectory. In the present study, the externally generated rotation flow generated by the visually simulated head turns shifted the perceived pseudo-FOE locations in the direction of the head rotations, therefore resulting in a perceived locomotor heading in the head rotation direction. As subjects were instructed to walk straight ahead, they corrected their perceived heading trajectory by veering in the direction opposite to the pseudo-FOE location or head turn direction. Such explanation is consistent with our previous findings showing that healthy subjects veer, in the physical world coordinates, in the direction opposite to perceived pseudo-FOE location when exposed to rotational flows [25]. The presence of walking trajectory deviations contralateral to the pseudo-FOE location in the simulated but not the active head turns in the present study suggest that non-visual cues are contributing to flow decomposition and hence to heading identification and steering, as suggested by previous psychophysical experiments on heading identification [15, 30]. An alternate explanation could be that non-visual cues contribute to the assessment and correction of an asymmetry in the optic flow field created by the head rotation, with no need to decompose directional components of the flow and identify heading from the optic flow itself. Regardless of whether the CNS correct for an asymmetry or a perceived change in FOE location, the fact that a straight locomotor trajectory was maintained when exposed to internally but not externally generated rotational flows (i.e. active as opposed to simulated head turn) suggests that non-visual cues do contribute to the control of locomotor heading when executing head turns while walking. Conversely, the fact that a trajectory deviation did occur in presence of an externally-generated rotational optic flow suggests that optic flow information is a powerful source of visual information that can impact on the control of locomotor heading. The multi-sensory fusion model supports the integration of both visual and non-visual elements in heading estimates [6].
There are multiple sources of heading perception during voluntary head turns. Crowell and colleague [3] demonstrated that vestibular information, neck proprioception and the efference copy of the head motor command were non-visual cues contributing to a proper identification of heading direction during active head rotations performed while seated. Eye movement information was also shown to contribute during active gaze shifts [1, 23]. In the present experiment, active head turns were also compared to visually simulated head turns while the subjects were fixating on a visual target. A fine eye/head coordination is normally present while changing direction during walking [8–10], as well as during voluntary gaze (eye/head) shifts [19]. It would have been ideal to collect eye movements during the present experiment, such that the relative contribution of eye movement information could be appreciated. Unfortunately, valid eye movements could not be collected due to the technical difficulty of stabilizing the combined HMD/eye-tracking system with respect to the subjects’ pupil while walking. A closer examination of head rotation amplitudes shows that the perceived virtual head rotations were similar across conditions but did not reach the full 40°, despite the fact that subjects were required to keep the target centered in the HMD screen. This suggests that some degrees of eye movement most likely contributed to gaze fixation on target, at least in the active head turn condition. In the simulated conditions, the virtual head rotation was slightly reduced by the opposite physical head rotation which, as expected, accompanied the steering response. Therefore, while non-visual information generated from the head movement have certainly contributed to control locomotor heading while actively turning the head, oculomotor responses most likely played a role, which warrants further investigation.
Only small changes in heading direction were provoked by relatively large magnitudes of visually simulated head turns. The heading changes averaged only ∼4° in the SHT condition, corresponding to ∼0.33 m of CoM mediolateral deviation after 5 m of forward walking. There may be several explanations for this. Firstly, it was demonstrated that richly textured scenes with geometrical information that contains parallax information, as in the present experiment, resulted in a strong source of retinal flow that yielded more accurate heading judgments [15, 32]. Furthermore, our average optic flow rotation rate reached approximately 11°/sec. While some studies report accurate heading judgment from retinal flow field, ie. without the contribution of any extra-retinal cues, for optic flow rotation rates up to ∼ 1°/sec [1, 23], others have identified good performances for rotation rates as high as 16°/s to 20°/s [16, 26]. Due to parallax information and relatively slow rates of rotational flow in this experiment, retinal flow itself might, to some extent, have contributed to minimizing heading errors in the simulated head turn conditions. As voluntary head turns while walking can reach speeds as high as 400°/s [13, 20], however, it appears reasonable to conclude that non-visual cues would normally be contributing to the control of a straight trajectory when turning the head while walking. Secondly, the fact that gaze, although required to fixate onto a target, was free to shift and scan the travel path while walking, might have further contributed to minimize the heading errors [33]. Thirdly, instructions and expectations as to whether the individuals will be travelling along a straight or curved path also makes a difference in heading judgment [14]. In the present study, instructions were similar across conditions. The order of presentation of the active vs. simulated head turns condition were block randomized, and head turn directions were randomly presented and interspersed with catch trials (FOE at 0°) within each block. Expectations, if any, would have been minimized and counterbalanced among subjects. Moreover, other powerful sources of sensorimotor information associated with locomotion were present in this study, which was at variance with previous optic flow studies conducted in stationary sitting. The CNS has the ability to re-weight and up-regulate accurate sources of sensory information when subjected to incongruent sensory information [17]. It is likely that the small steering changes observed in the simulated head turn condition also reflects the ability of the CNS to re-weight the sensory information in favour of eye, neck, and leg proprioceptive information, as well as vestibular information, which all signalled that no change in heading orientation occurred. Finally, previous studies have shown that an interaction in steering strategies (optic flow vs. egocentric) was present when both optic flow and target elements were utilized during locomotion [2, 32]. Results from our study, however, show that the presence or absence of a target does not affect CoM heading direction and body segment orientation in the simulated head turn conditions. These results are consistent with the fact that the target element in the present study, which remained still and centered with respect to the HMD screen in the simulated head turn condition (SHT_T), could not be used as a goal to walk to.
In conclusion, this current study provides further insight on the role of visual and non-visual cues in the control of locomotor heading during horizontal head rotations. While results indicates that optic flow information is powerful and influences locomor heading, they also suggest that non-visual information is needed for an appropriate interpretation of complex optic flows such as those induced during head turns while walking in order to head in the desired direction. Through the understanding of the role and interaction of retinal flow and non-visual information, we can better understand the deficits in locomotor steering with vestibulopathy, and design virtual environments that will allow them to use or restore this ability.
Footnotes
Acknowledgments
The authors would like to thank all volunteers who participated in this study, as well as Christian Beaudoin for his technical assistance, and Jessica Berard for her assistance in data collection. The study was funded by the Canadian Institutes of Health Research (grant # MOP-77548). Anouk Lamontagne is supported by a salary award from the Fonds de recherche du Québec - Santé.