
Abstract
BACKGROUND:
When viewing a far target, the gain of the horizontal vestibulo-ocular reflex (VOR) is around 1.0, but when viewing a near target there is an increased response. It has been shown that while this convergence-mediated modulation is unaffected by canal plugging and clinically practical transmastoid galvanic stimulation, it is eliminated by a partial peripheral gentamicin lesion.
OBJECTIVE:
The aim of this study was to determine if convergence increases the gain during peripheral hyposensitivity elicited by cold thermal irrigation.
METHODS:
The high frequency VOR gain was measured using video head impulse testing immediately after the cold caloric stimulus in 9 healthy human subjects with the lateral semicircular canals oriented approximately earth-vertical.
RESULTS:
Before caloric irrigation, near viewing (15 cm) increased the average VOR gain by 28% (from 1 to 1.28). Cold (24°C) water irrigation of the right ear decreased the gain to 0.66 (far viewing) and 0.82 (near viewing) (22% difference). Although vergence also increased the gain for impulses to the left to the same degree before caloric stimulus, the caloric irrigation itself (applied to the right ear) did not influence the gain for contralateral impulses.
CONCLUSION:
In our experiments vergence increased the gain of the human angular VOR during peripheral hyposensitivity elicited by cold thermal irrigation. These results suggest that cold irrigation does not abolish the function of the nonlinear/phasic vestibular afferent pathway.
Introduction
The vestibulo-ocular reflex (VOR) stabilizes images on the retina by rotating the eyes at the same speed and in opposite direction of head motion. In everyday situations, the reflex has to adapt quickly to different requirements during unpredictable accelerations. For instance, it has been long established that the gain of the angular vestibulo-ocular reflex (aVOR) is greater than unity when viewing near targets [7, 16]. This vergence-mediated increase of the aVOR gain compensates for the relative translation of the eyes with respect to the target during the head rotation because if the head turns while fixating on a close target, the eyes should rotate through a larger angle than the head, to stabilize retinal images.
It has been suggested that this vergence-dependent gain increase is mediated by a highly modifiable, phasic component of the aVOR [7]. Until now, three types of experiments have been carried out to test the above hypothesis: ablation of peripheral hair cells and afferents using gentamicin [11], inhibition of low frequency VOR using canal plugging [10] and galvanic stimulation [2, 9]. Gentamicin and galvanic stimulation are supposed to act mainly on the irregular/phasic afferent input, canal plugging leaves irregular afferent activity untouched. Gentamicin and plugging do not cause acute vestibular asymmetry, in contrast to galvanic stimulation. The gain of the nonlinear/phasic pathway was more modifiable than that of the linear/tonic pathway during long-term adaptation using magnifying spectacles [3] and in the recovery of contralesional aVOR-gain after unilateral labyrinthectomy [6]. Gentamicin abolished the vergence-mediated gain increase possibly through damaging Type I hair cells [11] and their afferents. In contrast, after canal plugging, when the phasic pathway was possibly preserved, vergence-mediated gain increase was also present, although in the other experiment gentamicin reduced the far viewing aVOR gain during high-acceleration head impulses to approximately the same level as canal plugging (approx. to 0.5) [10]. The authors argued that, in contrast to gentamicin, canal plugging did not preferentially reduce the vestibular responses of irregularly discharging afferents. In these experiments the afferent resting rate was probably symmetric before the impulses. There is one more important result to complete the picture: the aVOR gain during far target-viewing is unaffected by galvanic stimulation, which suggests that aVOR gain during high frequency head impulses may be dominated by input from regular afferents [12].
Our goal was to design a new combination of these stimulus qualities by measuring vergence-evoked VOR gain increase during unilateral cold caloric stimulation. This setup can be compared to canal plugging (because it hypothetically leaves the irregular afferents intact) but with the difference that there is an acute vestibular asymmetry. Also, it can be compared to galvanic stimulation, because there is an acute asymmetry, but with the difference that the irregular pathways may be left intact during the stimulation. Therefore, the setup in our present study will answer the following question: does an acute vestibular asymmetry abolish the vergence-mediated gain increase?
Methods
Participants
Nine healthy individuals (4 women, 5 men, average age 32 years; min.:18; max.:51) with no history of otologic or cochleovestibular disorders were recruited. The study was approved by the institutional review board of the Petz Aladar Hospital, Györ, Hungary (PAMOK Hospital Protocol number 76-1-18/2015). The protocol was in accordance with the ethical standards laid down in the 1964/2013 (7th revision) Declaration of Helsinki for research involving human subjects and the investigators have obtained written informed consent from each participant. Before the measurements, the right eardrum was examined.
Video head impulse test (vHIT)
Video HIT (vHIT) was carried out with the ICS Impulse® video goggles (GN Otometrics, Taastrup, Denmark) in the plane of the lateral semicircular canal (SCC). For head impulses, the position of the right eye was recorded with 250 samples/s. We did not measure binocular eye position, but in order to control the vergence angle we asked the probands about possible double vision. Since none were reported, we presume that both eyes were appropriately converged. Because the vergence angle could not be measured directly, a limitation of the study may be that the hysteresis properties of Panum’s area will help maintain fusion for ocular disparities of up to 2° [4, 14], which would introduce an error of up to ∼10% in vergence angle for a target at 15 cm. Another concern may be that the rotational stimuli in these studies were head-on-body rotations delivered passively (head thrusts) with the axis of rotation approximately centered with respect to the otoliths. The axis of rotation for these stimuli is located behind the eyes, and compensatory eye movements in this condition are expected to be larger for the abducting than for the adducting eye. However, Lasker et al. [7] observed comparable gains for both eyes (when measured as peak eye velocity/peak head velocity).
Baseline HIT testing and caloric stimulation was done using standard calibration with the lateral SCCs in an earth-vertical position, so the subject’s head was inclined by 30° from supine, with the Reid’s baseline elevated 60°. Subjects were tested in a well-lit room (to ensure a small pupil) with an eye-level target straight ahead (and above because of the reclination) at a distance of 120 cm in front of them (far viewing) and 15 cm (near viewing). Caloric irrigation lasted 40 seconds and was carried out in the right ear in all subjects at a temperature of 24°C. As a baseline measurement and after irrigation, 6–8 high-acceleration head impulses were administered in an alternating fashion (3–4 to the right and 3–4 to the left (peak head velocity >120°/s) over a time interval of 30 sec. Near and far viewing has been applied alternatingly: after three to four impulses to the right and left the viewing distance has been changed from near viewing to far viewing. For example the protocol for a baseline measurement: three unpredictable, passive high acceleration head impulses to the right, then to the left for near viewing (15 cm), then the same with far viewing (120 cm), then again three impulses to the right and to the left for near viewing and again for far viewing. Then this protocol was repeated at the end of the thermal irrigation. This protocol created 6 to 8 gain values of individual impulses for every subject, for every side, for every distance per baseline measurement and again for the thermal irrigation.
Data analysis
Raw data of head and eye velocity traces were exported into a Matlab-file (The MathWorks, Natick, MA), and the aVOR gain (maximal eye velocity divided by inverted maximal head velocity) and latency (time difference between the maximum head velocity and maximum eye velocity) were measured off-line. Maximal head velocity value and time was determined by searching for the curve maximum value in the time interval between 0 and 50 ms after the head velocity maximum. Impulses, which were contaminated by a nystagmus fast phase were discarded and the first 6 impulses (three to the right and three to the left) were used. This protocol generated 6 gain values of individual impulses for every subject, for every side, for every distance per baseline measurement and again for the thermal irrigation. Statistics were done using Graphpad Prism® Software.
Results
In Fig. 1 we show all results for all individuals for baseline measurements and after cold caloric irrigation of the right ear. Data groups (9 individuals; 54 impulses each column) were compared using a non-parametric ANOVA (we did not assume Gaussian distributions), the Friedman’s test and Dunn’s post-tests. The results show the following:
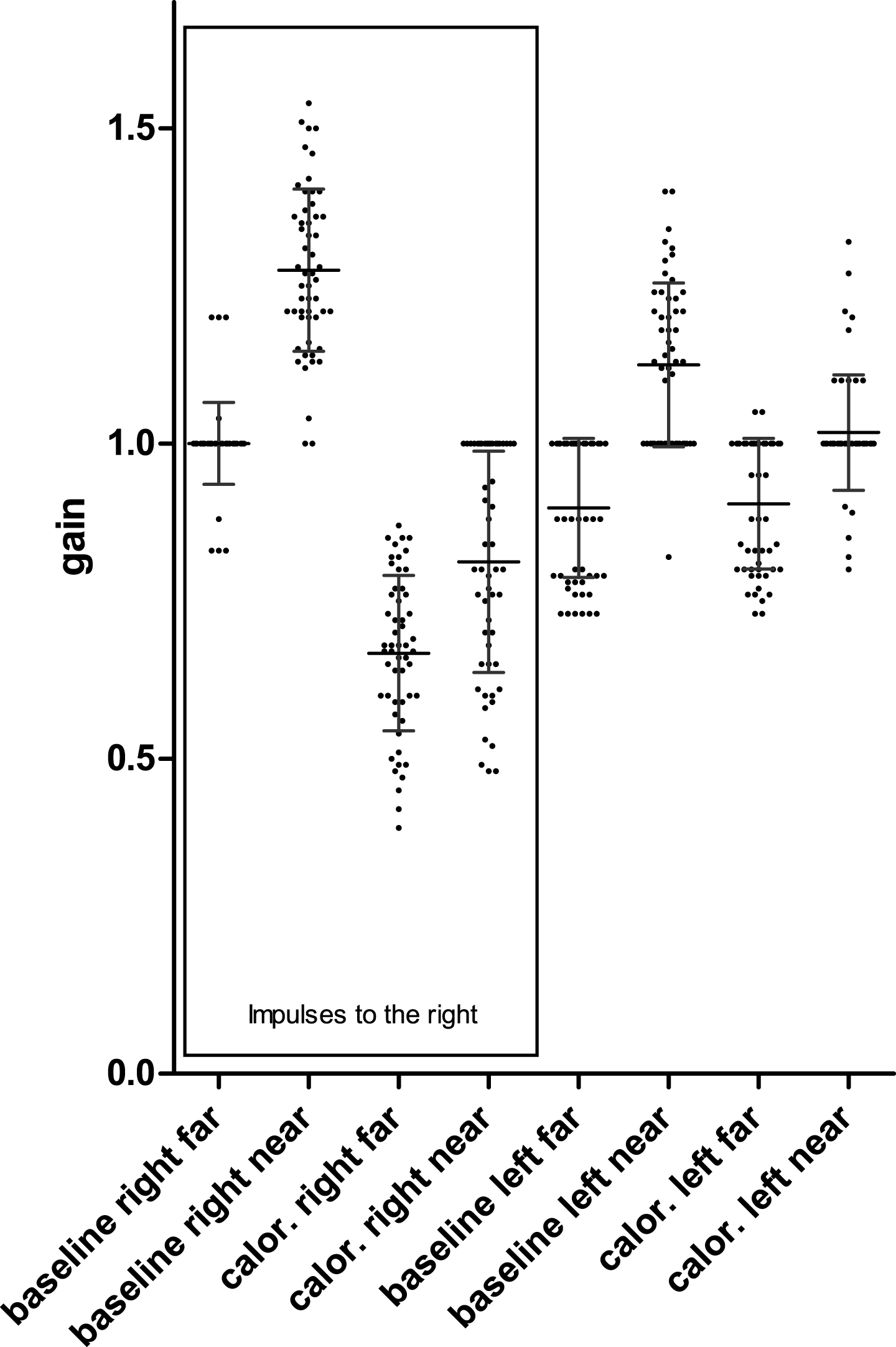
All results for all individuals for baseline measurements and after cold caloric irrigation of the right ear. Data groups (54 impulses collected from 9 individuals in each column) were compared using a non-parametric ANOVA and the arrows show significant differences according to the results of the Dunn’s post-tests. (***) means p≤0.001; (*) means p≤0.05. ‘baseline right far’ means impulses to the right before the thermal irrigation, ‘left’ impulses to the left etc. ‘calor.’: impulses after cold caloric irrigation.
Baseline measurements: Baseline aVOR-gains for head turns to the left did not differ from those to the right (neither during far nor near viewing) Highly significant difference between far viewing and near viewing in both directions. The gain increased in average 27.7 % for impulses to the right during convergence.
Measurements immediately after cold caloric irrigation of the right ear: Highly significant difference between baseline and caloric during far viewing for impulses to the right (caloric decreased the aVOR gain from unity gain to 0.67 in average), but no difference for impulses to the left. Highly significant difference between impulses during far viewing and near viewing (vergence) after caloric irrigation with impulses to the right (in average 22 % increase when near viewing) and significant difference between far viewing and near viewing during impulses to the left.
There was no significant difference between gain increase (gain change) due to vergence during impulses to the right before and after caloric irrigation as assessed using paired two tailed ‘t’ test (p value = 0.961, change in average (± SD) during baseline measurement: 27.7 % (± 13.2) and after caloric irrigation: 22.2 % (± 18.2). Baseline average (± SD) latency of the aVOR was 8 ms (± 3 ms), was the same during leftward and rightward head rotations and did not change significantly with viewing distance or after caloric irrigation as calculated by Friedman’s test and Dunn’s post-tests.
In summary, convergence increased the aVOR gain for impulses in both directions during baseline measurements and this increase was not abolished after cold calorics.
The baseline measurements in this study can be considered a reproduction of our previous experiments [15]. Those former results have been confirmed by the new data: the ipsilateral (and only the ipsilateral) aVOR gain could be lowered by the cold caloric irrigation. Our new results show that vergence-mediated gain increase was not abolished by the partial peripheral hyposensitivity. Apparently, similar to canal plugging [10] and unlike low doses of gentamicin in humans [11], cold caloric irrigation does not preferentially inhibit the vestibular responses of irregularly discharging afferents.
What may be the exact mechanism of the caloric hyposensitivity? The aVOR reflex during far viewing is probably transmitted by the linear pathway via regular afferents [13]. The origin of the gain decrease has to be more peripheral than the linear afferent nerves, because the shift of the set-point in a linear system should not influence the response amplitude. Our experimental setup creates a marked asymmetry of the afferent discharge, as it can be judged by the ensuing caloric nystagmus. This asymmetry is caused by the sustained geotropic (ampullofugal) deviation of the hair cell stereocilia on the side of the thermal irrigation. The deflection decreases the spike activity probably in the afferents by detuning the mechanoelectric transduction (MET) channels in the stereocilia. The transduction channels in the stereocilia of the vestibular (and cochlear) hair cells work like finely tuned critical oscillators e.g., operating near their dynamic instability, around a Hopf-bifurcation (for review see [1]). A bifurcation occurs when damping is close to zero. On one side of the bifurcation, damping is negative and the unstable system oscillates spontaneously, while on the other side, damping is positive and the oscillator is stable, yet neither sensitive nor finely tuned. It is when poised near the bifurcation that the critical oscillator displays the key features of active amplification: high sensitivity, nonlinear amplification, and level-dependent frequency tuning. Stereocilia are tuned to a specific stimulus frequency. Around the bifurcation, that means, when the stereocilia is around the operating point and when it is stimulated with a frequency to which it is tuned, the stereocilia react with by a burst of nonlinear activity, which amplifies the signal during transmission. This activity is averaged between several afferents, and this summation constitutes the afferent signal. However, at other stimulus frequencies or when the stereociliar bundle is deflected away from its operating point the mechanoelectrical transmission is linear and the gain is lower (because of the missing amplification). Here we suggest that the gain decrease after cold thermal irrigation originates in the mechano-electrical transduction (MET) channels of the type I and II hair cells in the peripheral regions of the lateral semicircular canal crista. In the present paradigm, deflecting the cupula decreases the spontaneous activity in the afferents and possibly the nonlinear content by removing the stereocilia from the bifurcation. This also decreases their sensitivity to sudden, high frequency accelerations: this is why the gain of the aVOR decreases. We also argue, that the remaining nonlinearity, which is necessary to drive the irregular afferents and the phasic pathway, originates in the MET-channels of the type I hair cells in the central regions of the crista. The acceleration signal of a strong cold-water caloric stimulus did not drive the irregularly firing afferents in an area to make them insensitive to high acceleration, possibly because of the ability of type I hair cells for fast adaptation. According to our results, the sterociliae of the peripheral and central hair cells were not uniformly affected by the cold caloric inhibition: the overall gain decrease was from unity gain down to 0.66, whereby vergence increased the gain approximately by the same amount of percentage before and after caloric irrigation. Type I hair cells, positioned in the central zone of the cupula, are less sensitive to low frequency stimulation, which the sustained caloric reaction constitutes. They are tuned to higher frequencies, and quickly adapt to the deflected position of the cupula. Therefore, in spite of the sustained deflection during head impulses they might still be able to provide information about the high frequency impulses through their irregular afferents, thereby making fast, vergence-mediated changes of the gain possible. In these structures (the MET-channels and the parallel pathways) this remarkable feature of the peripheral vestibular endorgan arises: in spite of suffering sustained deflections the cupulae is still able to inform the central vestibular structures about unpredictable, high frequency angular accelerations.
Based on the above hypothesis, we also suggest that the nonlinearity encountered in the phasic vestibular pathway in general could be originated in the transduction channels of the hair cells. It is likely that this phasic response is occurring pre-synaptically [5]. In this hypothetical framework type I or II hair cells vary their contribution depending on the axon they are connected to (David Lasker, Johns Hopkins University School of Medicine, personal communication). Some axons, such as the calyx only afferents, have the ability to transmit high frequency stimuli. Because there is a correlation between the regularity of discharge and the value of the rotational 1st order lead term, there may be a link between the post-synaptic gain of an afferent and the high frequency dynamics of that afferent. Increases in post-synaptic gain will cause an increase of the sensitivity to synaptic noise, which will in turn increase the irregularity of firing. The easily modifiable nonlinear/phasic pathway probably provides the signal by which sensitivities of corresponding neurons (position-vestibular-pause and eye-head-velocity neurons) in the vestibular nuclei can be increased during near viewing [8]. It has been suggested that irregular vestibular nerve afferents modify the gain and phase of the vestibular response of these secondary vestibular neurons [2].
We have to mention here the confounding effects of the possible utricular stimulation, because with near viewing there is a sideways acceleration and the linear VOR is mediated by the otolithic organs. Migliaccio et al. [10], who examined vergence-mediated gain increase using diagonal head impulses after canal plugging, argued that otolith contribution played a minimal role in the vergence-mediated gain increase in their experiments partly because of geometrical reasons. We also think that utricular activation/inhibition played a negligible role in our results because caloric reaction does not influence utricular afferent activity and other parameters were not changed during stimulation.
In conclusion we suggest that cold thermal irrigation constitutes a low frequency stimulation that causes an asymmetry of the linear/tonic vestibular afferent activity, and partly inhibits the gain during high frequency/high acceleration head movements through the sustained cupular deflection (by shifting from the optimal operating point). However it does not abolish the context-dependent gain modulation, because the highly modifiable nonlinear/phasic pathways retain their function.
Conflict of interest statement
The authors report no conflict of interest.
Footnotes
Acknowledgments
We thank the director of our hospital László János Tamás and the chief of the ENT department, Tamás Tompos for their support. We also thank an anonymous reviewer for improving the paper by useful suggestions.