
Abstract
BACKGROUND:
Vibration applied on the mastoid has been shown to be an excitatory stimulus to the vestibular receptors, but its effect on vestibular perception is unknown.
OBJECTIVE:
Determine whether mastoid vibration affects yaw rotation perception using a self-motion perceptual direction-recognition task.
METHODS:
We used continuous, bilateral, mechanical mastoid vibration using a stimulus with frequency content between 1 and 500 Hz. Vestibular perception of 10 healthy adults (M±S.D. = 34.3±12 years old) was tested with and without vibration. Subjects repeatedly reported the perceived direction of threshold-level yaw rotations administered at 1 Hz by a motorized platform. A cumulative Gaussian distribution function was fit to subjects’ responses, which was described by two parameters: bias and threshold. Bias was defined as the mean of the Gaussian distribution, and equal to the motion perceived on average when exposed to null stimuli. Threshold was defined as the standard deviation of the distribution and corresponded to the stimulus the subject could reliably perceive.
RESULTS:
The results show that mastoid vibration may reduce bias, although two statistical tests yield different conclusions. There was no evidence that yaw rotation thresholds were affected.
CONCLUSIONS:
Bilateral mastoid vibration may reduce left-right asymmetry in motion perception.
Introduction
The vestibular system, located in the inner ear, senses three-dimensional angular and linear motion of the head, and provides critical cues for gaze stability, posture and perception. Mechanical vibration applied to the mastoid is known to stimulate the vestibular system. For example, a common clinical “vibration test” for unilateral vestibular hypofunction involves the application of mechanical vibration to the mastoid [40, 90], with the elicitation of nystagmus in the presence of bilateral mastoid vibration (in the range of 60 Hz to 100 Hz) indicating unilateral peripheral dysfunction. Vibration applied on the mastoid has been shown to be an excitatory stimulus to the vestibular receptors [18, 91], using both sinusoidal stimuli (i.e., “pure tones”) ranging from 50 Hz to 4000 Hz and short impulsive stimuli (i.e., “clicks”). It has been proposed that when vibration is applied on the mastoid, the compressional and shear waves generated by the vibration are propagated through the skull towards the inner ear [17, 19]. Upon reaching the inner ear, these bone-conducted vibrational waves displace the receptor hair cells, similar to that produced by natural vestibular stimulation such as angular or linear head motion. In the discussion section we elaborate on the effects of vibration on the two primary vestibular organs, the semicircular canals and the otolith organs.
More broadly, the addition of noise to sensory systems – i.e., a stimulus that does not convey information about the physical quantity of interest – has been shown to influence both sensorimotor and perceptual responses. For example, balance performance changes when electrical noise is applied to the vestibular system [36, 82], the ankle [60] and knee muscles [38], or when mechanical vibration is applied to the bottom of the feet [14, 75] and to the muscle tendons [7]. Similarly, perception of low amplitude visual, auditory and somatosensory signals is modified by the presence of external noise [13, 92]. External noise can either increase or decrease detection or recognition thresholds depending on the level of noise [1, 67]. For example, recent work found that galvanic vestibular stimulation (GVS) with optimal levels of noise lowers perceptual motion thresholds for motion along some axes [32, 50].
Vestibular perception has been studied using a self-motion perceptual direction recognition task in which subject have to report the perceived direction of threshold-level motion stimuli in the dark and their responses are fit using a cumulative Gaussian distribution function that is characterized by two parameters: threshold and bias [9, 95]. Threshold is defined as the standard deviation of the cumulative distribution and refers to the smallest motion that can be reliably perceived. Thresholds quantify the precision of vestibular perception. Bias is defined as the mean of the distribution and refers to the motion perceived on an average when exposed to null stimuli [39, 65]. Vestibular bias may arise from unequal contributions of the left and right vestibular organs or other central imbalances in the sensation and perception of different motion directions [34, 72]. Threshold and bias have been characterized for different planes of motion (e.g., translation along earth-horizontal inter-aural axis, translation along earth-vertical axis, yaw and roll rotation) and with different stimulus frequencies [85]. Studies have demonstrated that proprioceptive, visual or tactile cues from the motion platform are either absent or minimal and that vestibular cues predominate for the perception of motion direction in this kind of task [10, 85]. For example, patients having bilateral vestibular dysfunction had significantly higher perceptual thresholds when compared to subjects with intact vestibular function [85].
Despite these studies, to our knowledge, the effect of mechanical mastoid vibration on vestibular perception has not been investigated. Hence we tested the effect of vibration on vestibular perception of yaw rotations in a direction-recognition task. For the remainder of this paper, we use “vibration” to refer to continuous, bilateral, mechanical vibration of the mastoids using a random, noisy stimulus with a broad range of frequencies (1–500 Hz). These frequencies were chosen because they correspond to the behavioral bandwidth of vestibulo-ocular responses, which responds up to 50 Hz [42, 76] and even higher [79, 80] and because vibration at these frequencies have been shown to elicit nystagmus and body sway in humans [12, 90]. Recent reports have shown that perceptual vestibular thresholds are correlated with vestibular migraine [55], age [5], peripheral deficits [15, 85], medication use [22] and performance in a clinical balance test [5, 45]. Furthermore, these perceptual thresholds assay neural noise [39], which models have shown impacts dynamic processing of vestibular cues [6, 59]. Thus an improvement in vestibular perception with the use of mechanical vibration may have potential benefits for certain populations.
Methods
Ten healthy adults (M±S.D. = 34.3±12 years old) participated in the study. The participants were screened using a questionnaire to assess for any history of dizziness, vertigo, back or neck problems, neurological, cardiological or any other general physical problems. The study was approved by the institutional committee for protection of human subjects (CPHS) at the University of Houston and the Institutional review board at Massachusetts Eye and Ear Infirmary (MEEI). All the participants provided informed consent prior to their participation and all study procedures were conducted in accordance with the Helsinki declaration.
All the participants’ yaw rotation vestibular perception thresholds and biases were tested under two conditions, with vibration and without vibration. The order of the two conditions was randomized across the participants. Yaw rotation was chosen because it primarily stimulates the semicircular canals, and vibration has been shown to activate semicircular canal neuronal units [91].
Vibratory stimuli
Vibration was applied bilaterally [12] on the mastoid using two small circular-shaped tactors (C-2 tactor, Engineering Acoustics, Inc.) having a diameter of 1.17 inches and a thickness of 0.3 inches. The tactors were attached to the mastoids using two-sided tape and Velcro straps. The tactors were controlled using custom designed software to generate longitudinal vibrations perpendicular to their surface. These vibrations were band-passed (1–500 Hz, colored noise) and had maximum amplitude of 0.635 mm [20, 21]. As no data exists in the literature relating vibratory noise to human vestibular perception, we chose a random, noisy stimulus with a broad range of frequencies to maximize the chance of eliciting a response.
Motion stimuli
Motion stimuli consisted of single cycles of sinusoidal angular acceleration about the earth-vertical axis at a frequency of 1 Hz (Fig. 1). This motion and frequency is known to primarily stimulate the semicircular canals. The stimuli were generated using a motion platform having six degrees of freedom (MOOG CSA Engineering, Mountain View CA, Model 6DOF2000E). The instantaneous angular acceleration of the motion stimulus can be represented by the expression a (t) = A sin(2πft), where A is the amplitude of acceleration and f is the frequency or the inverse of the period of the stimulus (T = 1/f). The frequency of the motion stimulus was fixed at 1 second for all trials, which concentrates most energy around 1 Hz [5]. The expressions defining velocity and displacement can be obtained by integrating acceleration and velocity respectively, velocity v (t) = A/(2πf) [1 - cos(2πft)] and displacement Δp (t) = A/(2πf) [t - 1/(2πf) sin(2πft)].
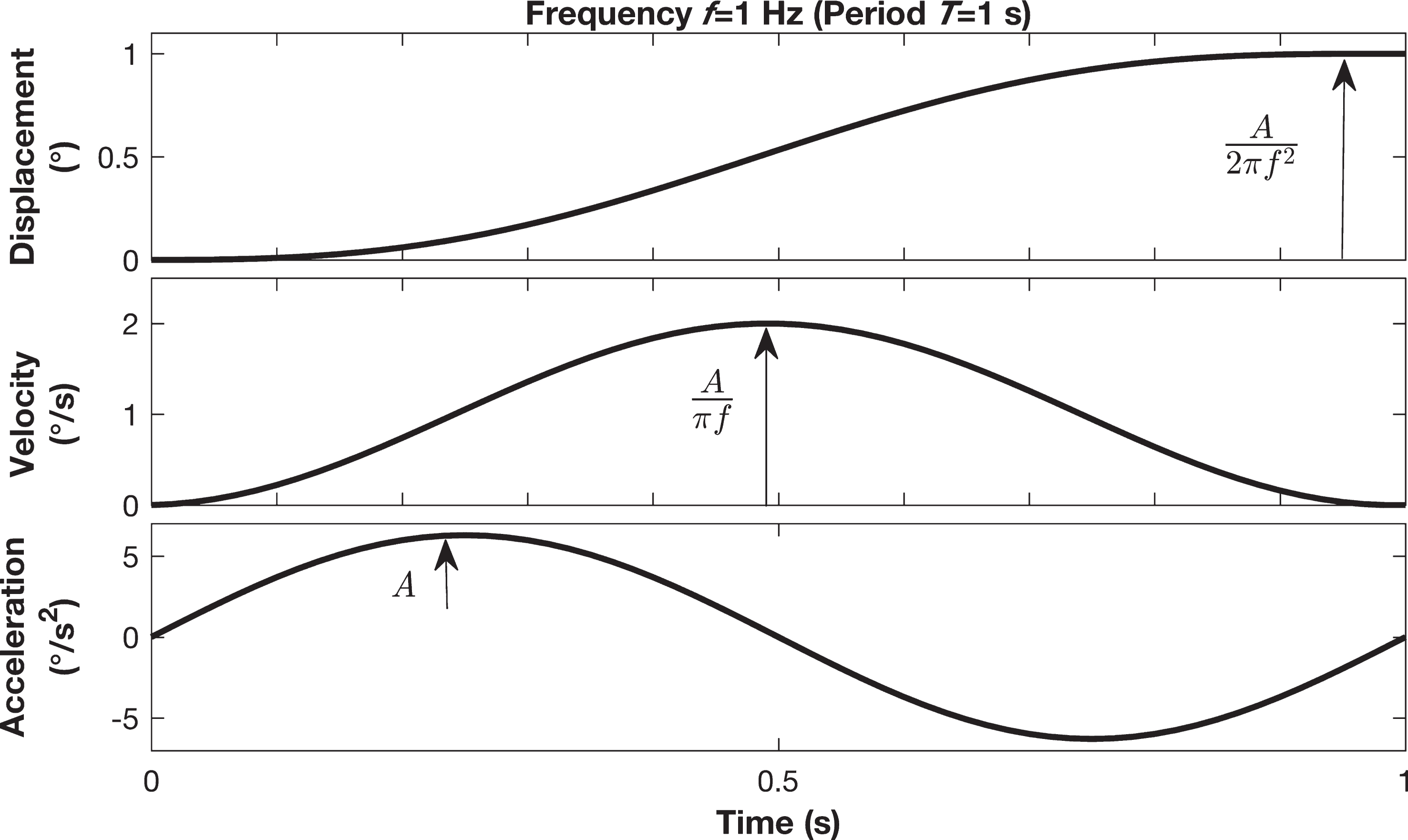
An example of one cycle of sinusoidal acceleration of yaw rotation. This corresponds to cosine bell velocity and sigmoidal displacement profiles at 1 Hz motion with a displacement of 1 degree.
The experimental procedures mimicked those used in earlier studies [10, 85]. All participants were tested in four blocks of 200 motion trials per block. Subjects received mastoid vibration on two of the four blocks. Thus there were a total of 400 trials per condition. Each block lasted for about 15 minutes. The expected coefficient of variation for the threshold estimates is roughly 10% [46].
Participants sat upright on a chair placed on the MOOG platform in a completely dark room that eliminated the availability of any visual cues. The participant’s body was secured to the chair using a five-point harness. Additionally, the participants’ head was held firmly with the help of a helmet attached to the chair to prevent any movements of the head as the chair rotated. To minimize haptic cues caused by air motion, all the skin surfaces of the participants were fully covered (long sleeves, gloves and socks). External noise and other auditory cues were also reduced by using noise-cancelling earplugs and by playing white noise.
The direction of the motion was randomized across trials. In each motion trial participants were rotated either to the left or the right. They were instructed to indicate the perceived direction of motion by pushing either the left or the right button. For trials they were unsure of the direction, they were asked to make their best guess. Each motion stimuli was preceded by a “warning” tone and followed by a tone of higher pitch to mark the end of the trial. The subjects’ responses to motion stimuli were recorded using a data acquisition system, which was used offline for data analysis.
As in previous studies, a three-down, one-up staircase adaptive algorithm was used to set the magnitude of the motion stimulus [9, 85]. “Three down” means that when the participant correctly reported the direction of three consecutive rotational trials, the velocity of the motion stimuli was reduced. “One up” means that when the participant incorrectly reported the direction of one trial the velocity of the stimuli was increased. We used the standard parameter estimation sequential testing algorithm (PEST) to determine the step size [84]. Testing for all participants started with large magnitude of rotations well above the typical yaw rotation threshold (40/s). The participants were given a mandatory break between each block during which the lights were switched on and they were allowed to get up from the chair and stretch.
Threshold and bias determination
Threshold and bias were determined separately for the two conditions (with and without vibration) by fitting a Gaussian cumulative distribution function to the data using a generalized linear model and probit link function [63, 85]. Specifically, the function call b=brglmfit(ω,y,’binomial’,’link’,’probit’) in Matlab 2016b (The Mathworks, MA, USA) was used to obtain the fits, where ω is the vector containing amplitude of motion stimulus and y is the vector containing the participants’ direction responses to the motion stimulus [11]. The output of this function contains elements that relate to the underlying Gaussian probability distribution, and were used to calculate threshold and bias. Threshold was defined as the standard deviation of the distribution and equal to the stimulus at which the subject correctly responds 84% of the times [40, 65]. Bias was defined as the mean of the distribution, located halfway between the lower and upper bounds of the psychometric function and equal to the stimulus that the subject correctly perceives as rightward 50% of the times. The sign of the bias denotes the direction along which the individual’s perception is biased [65]. For example a bias equal to –1.30/s denotes that on an average the individual perceives a rotation of magnitude equal to 1.30/s towards the left whenever exposed to null stimuli.
Statistical analysis
All statistical tests were performed using SPSS V.20 (IBM Corp, Somers, NY). As in earlier studies, a log transformation was applied on the thresholds before performing any statistical tests [37, 85]. Normality tests were performed before running any analyses. The distribution of both the biases and log-transformed thresholds of the two conditions were found to not vary significantly from normal.
Statistical testing was done using two approaches. First a paired t-test was used to compare thresholds and bias measured with and without vibration. Since a t-test can mask an effect when there is a substantial inter-subject variability in effect size, we also used a linear regression to determine the relationship between results obtained with and without vibration. A regression slope not significantly different from 1 was interpreted as a case in which there was no evidence of a difference in the measure between the two conditions across subjects. On the other hand, a slope significantly less than 1 was interpreted as a significant reduction in the measure with vibration from that without vibration. The slopes of the regression analyses were then tested using a one-sample t-test to determine if they were significantly different from a mean slope of 1.
Results
Figure 2 shows perceptual responses and psychometric function fits to those responses for one subject. The dashed-dotted line indicates the level where the subject perceived 50% of the motions to be leftward and 50% of the motions to be rightward, which is referred to as the vestibular bias. The dashed line in the figure indicates the level where the subject perceived 84% of the motions to be rightward. The one-standard-deviation threshold is the difference in stimulus amplitude between the 50% and 84% levels (i.e., the 84% level after adjusting for vestibular bias). For this subject, bias was larger without vibration (Fig. 2A, dashed-dotted line) compared to with vibration (Fig. 2B, dashed-dotted line). The change in threshold for this subject was minimal.

Example of typical subject’s responses fit with Gaussian cumulative distribution psychometric functions (A) without vibration; B) with vibration. The position of the circles represents the fraction rightward out of all responses at given amplitude and the size of the circle indicates the total number of responses. In some cases, responses for stimuli within smaller intervals were pooled for display only.
Figure 3A shows the relationship between thresholds measured with and without vibration, while Fig. 3B shows the relationship between biases measured with and without vibration. The paired t-test showed no significant differences in threshold or bias (p > 0.05) measured with and without vibration.

A) Relationship between thresholds with and without vibration. B) Relationship between bias with and without vibration. The dashed black line represents the regression line and the circular dots represent each subject’s experimentally-measured thresholds and biases. The solid line (unity line) represents a line with slope equal to 1. The area enclosed within the shaded grey region represents the 95% confidence interval of the regression line. The data point on the far left of figure B represents the visually identified outlier. The dot pointed by the arrow represents the data of a subject who had difficulty in performing the task. Both the data points were used in all analyses since they were objectively not found to be outliers.
A t-test can mask an effect when there is a substantial inter-subject variability in effect size. Thus, we also examined the effect of vibration on threshold using a linear regression between the log-transformed thresholds measured without vibration with those measured with vibration, which yielded a slope of 1.026 (S.E. =±0.065, Fig. 3A, dashed line). This slope was not significantly different from a slope of 1 (p = 0.162). This confirms that vibration had little effect on threshold. In contrast, when we examined the effect of vibration on bias using a linear regression between the biases with and without vibration, it yielded a slope of 0.4064 (S.E. =±0.109, Fig. 3B, dashed line), which was found to be significantly less than a slope of 1 (p < 0.001).
This result suggests that bias was significantly reduced in the presence of vibration when compared to that without vibration, but vibration had little effect on threshold. However, visual inspection of bias results indicated that one of the subjects might have been an outlier that could have disproportionately affected the regression slope; we now describe analyses that showed that this was not the case and that the subject had a minimal impact on the results. To objectively determine if there were outliers, we calculated the z-scores (standardized residuals) and Cook’s distance. To find outliers using z-scores, the residuals of the data during model fitting were determined and standardized to a normal distribution, and any standardized residual with an absolute value >3.29 was flagged as an outlier [30]. Cook’s distance measures the overall influence of a particular case over the parameters of the model, and any data with a Cook’s distance >1 was flagged as an outlier [30]. None of the data points qualified as an outlier. Despite these results, we conducted a separate regression analysis without the visually-identified outlier to ensure that our result was not an artifact. Removal of the outlier changed the slope by a small amount (slope = 0.331, S.E. =±0.210). In addition, we observed that some subjects had biases with different polarities with and without vibration. Since the interpretation of the effect of these subjects on the regression slope, as well as the physiological origins, are unclear, we also repeated analyses after excluding the three subjects whose biases switched polarity with vibration. This resulted in regression slope of 0.421 (S.E. =±0.119) with the outlier included and slope of 0.380 (S.E. =±0.240) without the outlier. These additional statistical tests provide further support that vibration changes bias by a factor of roughly 0.4.
This study examined the effect of mastoid vibration on perceptual yaw rotation threshold and bias in healthy adults. The results show that vibration had little effect on yaw rotation threshold. There was some evidence that vibration reduced perceptual bias, although the results should be interpreted with caution. Analysis using linear regression found that there was a statistically significant reduction in bias, while analysis using paired t-test found no statistically significant change. Future testing with a larger sample size could potentially account for inter-subject variability and confirm the results obtained in this study. We note that t-tests may not detect an effect when there is substantial intersubject variability in effect size.
Evidence suggests that mastoid vibration broadly stimulates the vestibular organs, including the semicircular canals (SCC) and otolith organs, and that the relative activation of the two organs may depend on the specific stimuli. For example, E.D. Young et al. [91] recorded single unit activity of vestibular nerve fibers from adult squirrel monkeys in response to vibration applied to the frontal bone and found that SCC neuronal units are more sensitive to vibration than otolithic units and specifically most sensitive in the frequency range of 200–400 Hz. On the other hand, I.S. Curthoys et al. [18] reported that sinusoidal vibration (200–1,500 Hz) and short impulsive stimuli result in preferential activation of otolithic neurons as opposed to SCC neurons in guinea pigs. It has been proposed that the preferential tuning of otolithic afferents to rapid and sudden changes in accelerations facilitates the activation of these afferents to single clicks of vibratory stimulus [26]. Thus, our stimuli, consisting of frequencies in the range of 1–500 Hz, likely activated both the otolith organs and SCC. However, given the interspecies differences, there is a possibility that the otolith organs are more affected by vibration than the SCC. Future experiments could aim to compare these effects by testing translational perceptual thresholds (e.g., interaural) in addition to rotation. Emphasizing the importance of testing both, a recent study using electrical (galvanic) stochastic vestibular stimulation [32] found that while translation perceptual thresholds decreased in the presence of noise, SCC thresholds were unaffected.
We do not know the mechanism by which vibration could reduce vestibular bias. In fact, little is known about the physiologic basis for perceptual bias [34, 65]. It is speculated that perceptual vestibular bias is related to the imbalances in the peripheral or central vestibular function or a combination of both [72]. Given that vibration was applied anatomically close to the vestibular organs, it certainly is likely that peripheral factors play an important role in altering perceptual bias in this study although that does not rule out additional central contributions. One of the causes for peripheral imbalance might be the presence of structural asymmetries. For example, the orientation of SCC pairs is reported to be slightly different in right and left ears in certain mammals [4]. Similarly, asymmetries in the size and mass of otoconia have been found in invertebrates and these asymmetries at the peripheral level can have a direct effect on vestibular functions [3, 58]. So, it could be possible that the structural asymmetries in the orientation of the SCC affects the fluid dynamics of the endolymph differently on the two sides, which in turn causes imbalance in the displacement of the hair cells and the activation of the vestibular neurons, resulting in perceptual bias. Bone conducted vibration is proposed to activate the vestibular receptors by modulating the fluid pressure [62] and producing pressure waves that travel through the inner ear and displace the hair bundles [17]. It is possible that these fluid pressure waves somehow corrected for the differences in the fluid dynamics of the SCCs produced by the structural asymmetries and thus reduced perceptual bias in the current study.
We modeled six scenarios with different assumptions to determine how vestibular bias may arise from unequal neural activity on the left and right sides [65, 72], including additive (Fig. 4A-D) and multiplicative (Fig. 4E-H) asymmetries. We assumed that: a) perceived motion is determined by averaging bilateral activity, with the polarity of the left ear reversed, mimicking the standard push-pull SCC configuration in which the brain effectively inverts one signal during bilateral integration; b) perceptual bias arises from non-zero average activity with zero motion; c) neurons respond linearly with amplitude and direction, consistent with experimental data for small stimuli [25]; and d) perceptual asymmetries are not centrally compensated, since this is inconsistent with perceptual experimental results, despite evidence of central compensation in motor reflexes.
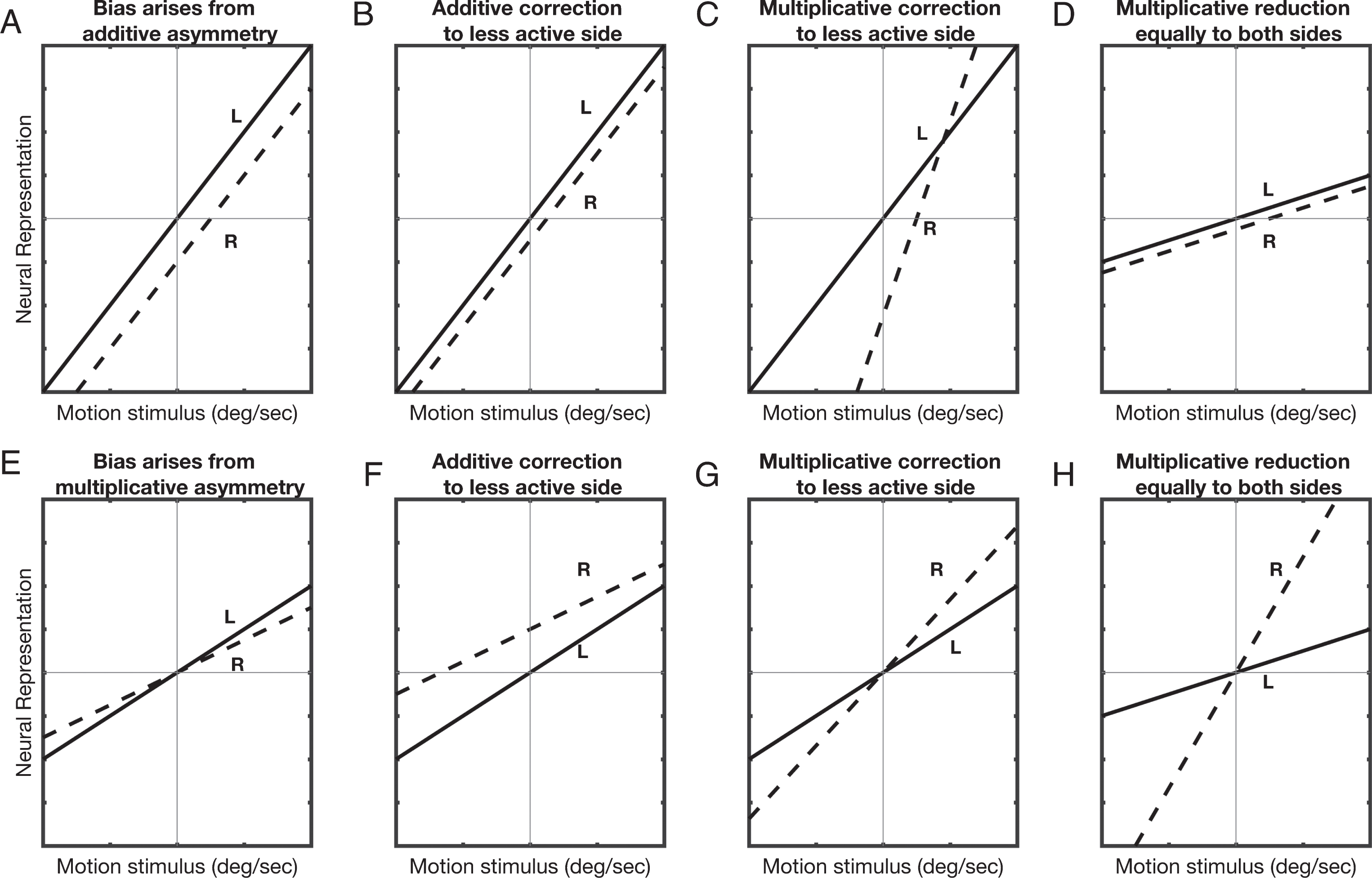
A) The neural representations of the left (solid line) and right (dashed line) semicircular canals as a function of motion stimulus amplitude for the additive asymmetry case. The neural activity of the right side is assumed to be less than the left side by a constant amount. B,C,D) As described in the text, three scenarios are considered for how the activity shown in (A) could be modified by vibration. E) In the multiplicative case, the sensitivity of the right side is assumed to be lesser than the left side. F,G,H) As described in the text, three scenarios are considered for how the activity shown in (D) could be modified by vibration.
We assume a constant right-left difference in the additive case (Fig. 4A-D). As motion velocity becomes more positive, neural activity increases for the right SCC (dashed line) and decreases for the left SCC (solid line; polarity reversed for display purposes). Figure 4B shows the predicted changes in responses if bilateral vibration increased activity by a constant amount only on the side with less baseline activity (i.e., the right side), resulting in average activity that is close to veridical and therefore consistent with our results. Despite evidence that vibration increases activity, we cannot explain how bilateral vibration would affect the less active side more. If both sides were equally affected (not shown), the two curves move in opposite directions by equal amounts, thus keeping the average the same. When correction occurs via an increase in sensitivity on the side with less baseline activity (Fig. 4C), i.e., a multiplicative correction, perceptual bias reduces at some amplitudes but increases overall. When correction occurs via equal bilateral reductions in sensitivity (Fig. 4D), perceptual bias is reduced. While this case is more plausible because vibration has bilateral effects, past experimental work found that vibration causes excitation rather than inhibition. We assume different right and left sensitivities for the multiplicative case (Fig. 4E). Since this predicts zero perceptual bias at zero motion amplitude, it does not match experimental results. Figure 4F,G,H show the same correction models as for the additive case. In summary, Fig. 4B, D are consistent with our results, although not previous experimental evidence.
Although the vibratory stimulus used in the current study was meant to stimulate the vestibular organs peripherally, the tactors that were used to generate the stimulus are generally used to provide tactile stimulation [20, 73–75]. Although unlikely, it is possible that the vibration did not have a direct effect on the vestibular organs and the changes in bias that we observed are a consequence of tactile stimulation caused by vibration. Many behavioral studies have shown that vestibular-somatosensory interaction have important effects on perception, which are mainly due to anatomical overlap of neural sites that are involved in the processing of vestibular and somatosensory inputs. For example, improvement in perception of faint somatosensory stimuli was reported in the presence of caloric [27], galvanic [28] or natural vestibular stimulation (whole body rotations) [29, 31]. The improvement in somatosensory perception in the presence of vestibular stimulation has been suggested to occur due to convergence of vestibular and somatosensory signals at bimodal neurons, where one sensory signal modulates the gain or synaptic connection in the afferent pathway of the other sensory signal. Several neurons that respond to both vestibular and somatosensory inputs have been identified by neuroimaging studies in certain regions of the brain [8, 94]. Taken together, these findings suggest the possibility that changes in vestibular bias produced by vibration in this study could result from vibration (tactile stimulus) induced central modulation of the activity of the bimodal neuron that is responsible for compensating for any peripheral imbalances.
A few methodological limitations in this study are worth noting. Firstly, the broad vibration frequency range (1–500 Hz) used is known to stimulate a number of sensory organs that are proximate to the mastoid, many of which have known contributions to spatial orientation or multisensory interactions with vestibular cues. For example, the posterior and superior SCCs or otolith organs could have been activated, and although they are relatively insensitive to the yaw stimulus used, they interact in central spatial orientation mechanisms [2, 66]. Also, one of the cervical muscles (sternocleidomastoid muscle) could have been activated, which is known to affect postural responses [61], and thus may also play a role in self-motion perception. Furthermore, the auditory system was likely activated [24], and multisensory interactions occur between the auditory and vestibular systems in spatial processing [71, 89]. In fact, auditory noise has been shown to change the processing of cross-modality cues [56]. Finally, cutaneous mechanoreceptors in the skin could have been activated [14, 75]. Additionally, the small sample size and limited vibratory and motion characteristics tested (yaw rotation, 1 Hz) are a few other limitations. Nonetheless, our statistical analyses, including the exclusion of artifacts from potential outliers, increase confidence in our conclusions. We expect that this investigation will provide the basis for the design of future experiments to address more specific questions using mechanical vibration like the most effective frequency range and intensity of vibration, and comparison of vibration effects on direction recognition thresholds of other motion planes (such as inter-aural translation or roll tilt).
Footnotes
Acknowledgments
This research was supported by NIH/NIDCD R03-DC013635. The authors thank Dr. Daniel M. Merfeld, the Director of the JVPL for providing us access to the MOOG device and Dr. Beom-Chan Lee for designing the software to precisely control tactor activation.