
Abstract
BACKGROUND:
The ocular vestibular evoked myogenic potential is otolith-dependent and has been suggested to be a manifestation of the linear vestibulo-ocular reflex (L-VOR). A characteristic feature of the translational LVOR (t-LVOR) is its dependence on the distance of a target.
OBJECTIVE:
To assess if viewing distance affects amplitude and latency properties of the ocular vestibular evoked myogenic potential (oVEMPs).
METHODS:
Bone- and air-conducted (BC and AC) stimuli were used to evoke oVEMPs in 10 healthy subjects. BC stimuli consisted of impulsive accelerations applied at the mastoids, AFz, Oz and Iz. AC stimuli consisted of 500 Hz tones delivered unilaterally to each ear. Target distances of 40 cm (near), 190 cm (intermediate) and 340 cm (far) were used for all stimuli.
RESULTS:
The largest amplitude oVEMP was obtained from Iz and the latency for AFz was shorter than for BC stimulation at other sites. We found no significant effect of target distance on oVEMP amplitudes for any of the stimuli used. There was a small but significant effect on latency with the nearest target having a longer latency (overall 12.4 ms vs 12.0 ms for the 2 more distant sites).
CONCLUSIONS:
Previously reported differences between latencies and stimulus sites for midline BC stimulation were confirmed. Target distance had no significant effect on oVEMP amplitude, which suggests it is not modified like other components of the t-LVOR.
Keywords
Introduction
The ocular vestibular evoked myogenic potential (oVEMP) is widely used as a means of assessing the otolith system, particularly the utricle [4, 25]. It has been well established that the amplitude of the response depends to a major degree upon the amount of eye elevation. The amplitude of the response increases with ocular elevation [13], most probably reflecting a reflex enhancement due to greater pre-activation of the target muscle [24]. Definitive evidence of its origin has come from single motor unit recordings from the inferior oblique muscle [29]. The inferior oblique is the main ocular elevator when the eyes are adducted [3, 16]. The oVEMP is commonly recorded with a fixed level of upgaze, typically 20–30 degrees above the horizontal.
The effect of target distance on the oVEMP has not been explored to date. This issue is not simply a practical one, but also relates more fundamentally to the origin of the reflex. Normal head movements consist of both angular and linear components and fully compensatory eye movements must also consist of both. The oVEMP, given its origin, is likely to be a manifestation of the linear vestibulo-ocular reflex (LVOR; [23, 27]). The linear VOR itself has two forms – translational and tilt/pitch, reflecting the two methods by which the otoliths and the utricle in particular can be excited. The translational LVOR has the specific property that its gain is inversely related to the distance of the target [5, 18]. When a target is close, the correction required for a translation of the observer is relatively large, and the opposite for distant targets. Thus, the effect of the distance of the target on the reflex may indicate whether the oVEMP is likely to be related to the translational LVOR rather than compensation for tilt/pitch.
Methods
Participants
Ten healthy subjects (23±4 yrs; 6 male, 4 female) with no previous neurological, audiological or vestibular deficits were recruited from the University of New South Wales. All subjects provided written informed consent in accordance with the Declaration of Helenski and the study was approved by the local ethics committee (Human Research Ethics Committee, Northern Network, South Eastern Sydney Local Health District).
Vestibular stimulation
The BC stimulus consisted of a 3rd order gamma waveform with a 4 ms rise time which produces impulsive whole-head accelerations [28]. Customized software was used to generate the gamma waveform using a CED micro1401 (Cambridge Electronic Design, Cambridge, UK) and fed to a power amplifier (model 2718, Brüel & Kjaer P/L, Denmark). BC impulsive accelerations were delivered via a hand-held mini-shaker (model 4810, Brüel & Kjaer P/L, Denmark) with attached perspex rod at an intensity of 20 V peak, equivalent to approximately 14 N pk FL. BC stimuli were delivered perpendicular to the head at the right and left mastoids and in the midline of the skull at AFz, Oz and Iz, locations that were determined based upon the 10–20 EEG methodology. A positive phase waveform (a compressive stimulus) was used for all stimulation sites (i.e. initial movement of the perspex rod towards the head).
The AC stimulus consisted of unshaped 2 ms, 500 Hz sinusoidal tone bursts at an intensity of 138 dB peak sound pressure level (pSPL), equivalent to 110 dB LAeq. These were generated using customised software, a CED power1401, a power amplifier and delivered unilaterally to each ear via calibrated headphones (THD-49, Telephonics Corp., Farmingdale, NY, USA). All AC recordings were made using an initial condensation polarity. Both AC and BC stimuli were delivered at a rate of 5/s.
oVEMP recordings
Surface electrodes (Cleartrace 1700-030, Conmed Corp., NY, USA) were used to record extraocular potentials. Recording electrodes positioned over the zygoma between the centre of the eye and the outer canthus (see [9] for mediolateral (ML) recording location details). Reference electrodes were positioned 2 cm below them. An earth electrode was placed near the suprasternal notch. Electrode impedance was kept below 5kΩ. Gaze was fixed at 30° above the horizontal and fixation points were located at three target distances; 40 cm (near), 190 cm (intermediate) and 340 cm (far). EMG signals were amplified (50,000x), filtered (5 Hz –1 kHz) and sampled (10 kHz) using a CED Power1401 and recorded using Signal software (version 6, Cambridge Electronic Design, Cambridge, UK).
Subjects were seated upright during oVEMP recordings and instructed to direct their gaze at either the near, intermediate or far viewing distance fixation points. Head position was not fixed but care was taken to ensure the subjects’ heads remained in a neutral position for all recordings. The order of stimulus presentation and fixation points was randomised. Recordings were performed from 10 ms before to 60 ms after stimulus onset for 100 individual trials and averaged. Negative extraocular potentials at the active electrodes are shown as upward deflections. Response amplitudes were corrected for any DC offset which was measured over the pre-stimulus interval.
Statistical analysis
The initial biphasic peaks were measured for oVEMP amplitude while the initial peak was used for latency measurements. Preliminary ANOVA was conducted using eye (right and left), stimulus site (mastoid, AFz, Oz, Iz and ear for AC stimulation) and viewing distance (near, intermediate and far). As there was no overall effect of eye for amplitude and latency measures this factor was collapsed and averaged. Subsequent analyses therefore used a 2-way ANOVA design with stimulus site and viewing distance as factors. Separate ANOVAs were conducted for amplitude and latency measures. Post-hoc t-tests were used to investigate main effects with the significance threshold set at P = 0.01 due to the number of comparisons. Values in the text are reported as mean±S.D and in the figure as mean±S.E.M.
Results
Grand mean recordings shown in Fig. 1A demonstrate the similarity in oVEMP waveforms across the three viewing distances (far, intermediate and near) and five stimulus sites (mastoids, AFz, Oz, Iz and AC stimulation). Overall mean oVEMP amplitudes by stimulus site were 8.2±3.9μV (mastoid), 7.3±3.1μV (AFz), 11.8±5.0μV (Oz), 13.6±5.4μV (Iz) and 4.5±1.6μV (AC stimulation) with Iz producing the largest peak-to-peak amplitudes (main effect of stimulus site; F (4,36) = 12.1, P < 0.001, Fig. 1B top row). Amplitudes for Iz stimulation were significantly larger than for AFz (t (9) = 3.5, P = 0.006), mastoid (t (9) = 3.3, P = 0.009) and AC stimulation (t (9) = 5.9, P < 0.001). Overall, viewing distance did not affect oVEMP amplitudes (main effect of viewing distance; F (2,18) = 0.9, P = 0.423) nor were there any differences between stimulus sites (interaction between viewing distance and stimulus sites; F (8,72) = 0.6, P = 0.741). For amplitudes, no individual subject values were in excess of 2 SDs of the mean across the viewing distances and stimulus sites.
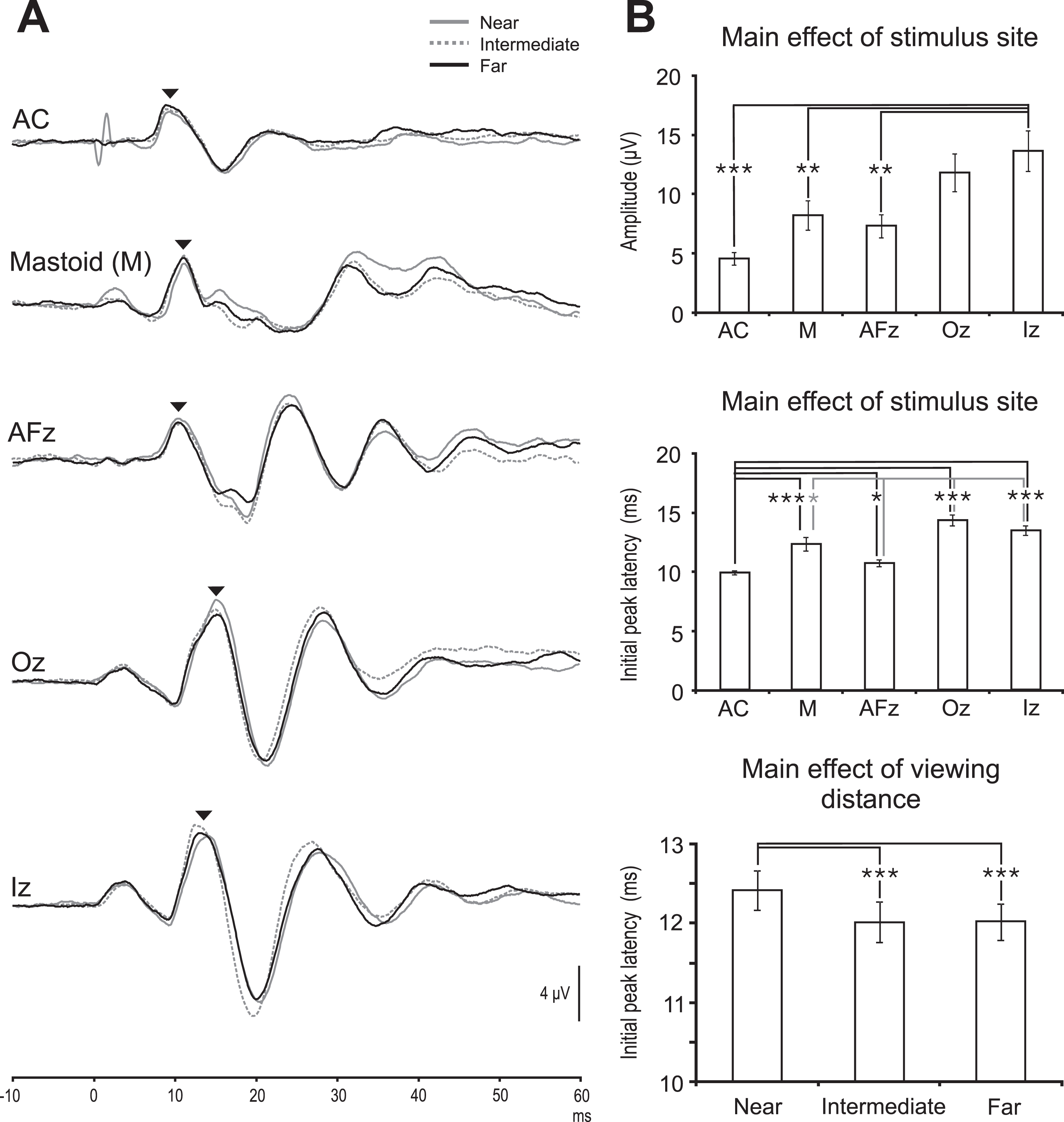
(A) Grand mean oVEMP recordings from the left eye to AC 500 Hz stimulation and BC impulses across the three viewing distances (near: grey lines, intermediate: grey dashed lines and far: black lines). Black arrows indicate the initial peak of the oVEMP response for each of the five traces. BC stimulation at the mastoids (M), AFz, Oz and Iz as well as AC 500 Hz delivered at the ears showed similar oVEMP waveforms across the three viewing distances. All stimuli were given at 10 ms after the trace onset and stimulus artefact is visible in several of the traces before the main response. (B) (Top) oVEMP amplitudes for Iz stimulation were significantly larger than for AC and BC at the mastoid as well as at AFz. (Middle) latency comparisons for AC (dark line) and AFz (grey line). AC had significantly shorter latency than for all BC sites except AFz. AFz latency in turn was significantly shorter than for Oz and Iz. (Bottom) the main effect of viewing distance on initial peak latencies demonstrated later responses overall for the near target. ***P = or<0.001, **P < 0.01, *P < 0.05.
For latency, mean initial peak values overall were 12.3±1.8 ms (mastoid), 10.7±0.9 ms (AFz), 14.3±1.4 ms (Oz), 13.5±1.3 ms (Iz) and 9.9±0.5 ms (AC stimulation) and differed significantly between the sites (main effect of stimulus site; F (4,36) = 25.2, P < 0.001, Fig. 1B middle row). AC latencies were significantly earlier than for Oz (t (9) = 11.6, P < 0.001), Iz (t (9) = 7.2, P < 0.001) and mastoid stimulation (t (9) = 4.7, P = 0.001) and showed a trend of being earlier when compared to AFz stimulation (t (9) = 2.7, P = 0.02). Latencies for AFz stimulation were earlier for than for Oz (t (9) = 8.0, P < 0.001) and Iz stimulation (t (9) = 7.2, P < 0.001) and tended to be earlier when compared to mastoid stimulation (t (9) = 2.5, P = 0.03).
Initial peaks were later for the near viewing distance target (main effect of viewing distance; F (2,18) = 8.6, P = 0.002, Fig. 1B bottom row). Initial peak latencies for near viewing (12.4±0.8 ms) were significantly later than for the intermediate (12.0±0.8 ms) and far (12.0±0.7 ms) targets (t (9) = 4.8 and 5.0, P < 0.001 for both comparisons). The effect of viewing distance on initial peak latencies did not differ between stimulus sites (interaction between viewing distance and stimulus site; F (8,72) = 1.2, P = 0.330). In individual subjects, latency values exceeded the mean by 2 SDs in one subject for AFz stimulation and, in another subject, for Oz, Iz and AC stimulation.
The otolith organs detect linear acceleration, both translational acceleration and changes in the direction of gravity induced by tilt [1]. The otolith organs, specifically the utricle, are believed to be the origin of the irregularly firing afferents which evoke the oVEMP [6, 12]. Potentially the oVEMP could be part of either of the two otolithic reflexes described – the linear VOR and the tilt/pitch VOR [23, 27]. Initially it was suggested that the “bowing” of the head laterally might be the mechanism by which stimulation at AFz and Fz might excite the otoliths [15]. However, there are a number of problems with this proposal when attempting to explain the pattern of excitation seen when a BC stimulus is applied at different sites in the midline [11]. In this study [11] using different midline locations, neither linear acceleration changes nor the imposed pitch could explain the range of responses recorded, particularly the changes occurring around AFz/Fz. In this region, the response abruptly reversed in polarity with no specific change in linear acceleration in any direction [10]. The polarity of the responses at Nz and Iz however broadly fitted with what might be expected for the translational VOR for an elevated midline target: a compressive stimulus at Iz caused initial excitation and the same stimulus at Nz caused initial inhibition. Our present results confirm some of the other unusual aspects of the midline stimulation sites noted by Govender and Colebatch [11] and others [19]. These include the disproportionately short latency occurring for AFz/Fz stimuli compared with other sites of BC stimulation, a finding which led Govender and Colebatch [11] to propose that there may be a direct action on the hair cells at this site. In addition, we confirmed that the response at Iz was the largest obtained, suggesting it would be a suitable site for eliciting oVEMPs.
Paige and Seidman [21] summarised experiments in the monkey regarding the linear VOR. They noted that ocular torsion could occur with purely translational accelerations, particularly if these were slow and that these movements were not compensatory for the applied acceleration. Torsion could be seen with interaural and vertical (pitch) responses with naso-occipital accelerations, responses which were thought to represent the tilt-LVOR, correcting for apparent head rotations with respect to gravity. Angelaki and McHenry [2] showed that the LVOR in monkeys could start around 10 ms for the nearest targets. A robust short latency LVOR can be seen in humans with both lateral (heave) and antero-posterior (surge) linear accelerations [22, 26].
Despite the evidence for the translational LVOR in humans, we found no significant change in amplitude with changing target distance. It may be that the irregularly-firing otolith afferents which are mainly excited [7] are not subject to this modulation. Some support for this interpretation comes from the observation by Lempert et al. [17] that the utricular afferents mediating the tLVOR respond only to acceleration directed ipsilaterally. In contrast, we have found similar oVEMP responses, albeit inverted, in response to both directions of lateral acceleration [14]. Possibly the oVEMP pathway is too direct to receive distance-related modulation and, like the cervical VEMP (cVEMP), is mainly controlled by the level of target muscle tonic activation. There was a small but significant change in latency, becoming slightly later for the nearest target. In the absence of any amplitude change, this latency change for the initial peak may be due to slight changes in the length of the muscle occurring with increased vergence. Minken and Van Gisbergen [20] reported that there was only a small change (about 1 deg., their Fig. 1) in ocular torsion with vergence from 120 to 20 cm. Previous evidence suggests that the size of the oVEMP response is determined, like the cervical VEMP, by tonic activity of the muscle of origin, the inferior oblique [24]. Given the role of the inferior oblique in ocular torsion one might conclude that, given the lack of change in torsion, there is no change in tonic activation in the inferior oblique and therefore no effect of antero-posterior target position would be expected. However, eye elevation also causes no change in ocular torsion [30] while being a strong determinant of the amplitude of the oVEMP [13]. Simply assessing the degree of ocular torsion ignores possible co-contraction of the oblique muscles which has been shown to occur and which would mean greater tonic activation than suggested by the degree of ocular torsion [8].
In contrast to gaze angle, target distance makes little difference over a wide range. We have found no evidence for the oVEMP being modulated by target distance as expected for the t-LVOR. The practical consequence of our findings is that, for oVEMP amplitude, only the angle of gaze needs to be controlled over a wide range of target distances, for a given site and mode of stimulation. Latency is also strongly affected by mode and stimulation site but can be prolonged by using very near targets.