
Abstract
BACKGROUND:
Vestibular compensation is the process by which the central nervous system (CNS) attempts to adapt to the loss of vestibular sensory inputs. As such, the compensation process is critically involved in the vestibular rehabilitation programs that are implemented by physical therapists for patients with vestibular disorders. One hypothesis regarding vestibular compensation, which has persisted in some of the published vestibular compensation literature and particularly on some vestibular and physical therapy websites, is the ‘cerebellar shutdown’ or ‘cerebellar clampdown’ hypothesis proposed by McCabe and Ryu in 1969. This hypothesis proposes that the cerebellum inhibits neuronal activity in the bilateral vestibular nuclei (VN) following unilateral vestibular loss (UVL), causing the VN contralateral to the UVL to be electrically silent during the early phases of vestibular compensation. Despite a wealth of evidence against this idea, it has gained traction amongst some physical therapists and has implications for vestibular rehabilitation early in the compensation process.
CONCLUSIONS:
In this paper it is argued that the ‘cerebellar shutdown’ or ‘clampdown’ hypothesis is inconsistent with well accepted neurophysiological and imaging evidence and that it is also logically flawed.
Introduction
‘Vestibular compensation’ is a term that refers to the process by which the CNS gradually adapts to the loss of the normal transmission of vestibular sensory input from the peripheral vestibular system (although it can also be used to refer to the process of recovery of less common central lesions as well). This loss of peripheral vestibular input can be unilateral or bilateral, complete or partial; it can occur suddenly (e.g., acute vestibular neuritis) or gradually over time (e.g., vestibular loss associated with aging or growth of a vestibular schwannoma) and it may be paroxysmal (e.g. Meniere’s Disease) or permanent (e.g. aminoglycoside antibiotic ototoxicity). This review focusses on a specific hypothesis for one mechanism of vestibular compensation, variously referred to as the ‘cerebellar shutdown’ or ‘cerebellar clampdown’ hypothesis. First proposed by McCabe and Ryu in 1969 [1], this hypothesis stated that early in the vestibular compensation process (e.g., 1 day) for unilateral peripheral vestibular dysfunction, when asymmetric neuronal activity exists between the vestibular nucleus (VN) on the side of the lost vestibular function, and the VN on the intact side, the cerebellum ‘shuts’ or ‘clamps’ down neuronal activity in the VN bilaterally, in order to reduce the imbalance in spontaneous neuronal activity that generates spontaneous ocular nystagmus and postural asymmetry. Despite evidence to the contrary, this hypothesis has proven to be very durable and is cited in publications (e.g., [2–5]) and on websites such as those for the Vestibular Disorders Association (VEDA) [6], Interacoustics [7] and other websites (e.g., [8, 9]). Whether or not this hypothesis is accepted as reasonable has considerable implications for the practice of vestibular rehabilitation during the early phases of recovery from unilateral vestibular loss (UVL), since it suggests that both VN will be inhibited and therefore there should be minimal vestibular function even in the VN on the intact side.
In this paper, I argue that the ‘cerebellar shutdown’ or ‘cerebellar clampdown’ hypothesis is demonstrably inconsistent with well accepted neurophysiological data that have been published since the 1980’s, that it is seriously misleading and has the potential to undermine vestibular rehabilitation therapies practiced by physical therapists. No attempt will be made to review vestibular compensation generally since many reviews have been published over the last several decades (e.g., [2, 10–21]), and these can be consulted for more detailed information on the nature of vestibular compensation and its mechanisms.
Vestibular compensation and the cerebellar shutdown/clampdown hypothesis
The cerebellar shutdown hypothesis specifically concerns the ocular motor and postural symptoms that develop following the unilateral loss of vestibular input (UVL) [1, 22], since following bilateral loss (BVL), neuronal activity in both VN will be reduced anyway. Following UVL, the most obvious symptoms comprise a high frequency, mainly horizontal, spontaneous nystagmus in light, with the quick phase directed toward the intact labyrinth, which in humans coincides with the experience of vertigo; in some mammalian species, such as rats and guinea pigs, there is often a tilt of the head and body toward the hypofunctional side (see Fig. 1). These symptoms are clearly the result of asymmetric neuronal activity between the left and right VN, as a consequence of lost input from one peripheral vestibular labyrinth (see [3, 20] for reviews). There is a large decrease in the gain (i.e., eye velocity/head velocity), and abnormalities in the phase and/or timing, of the vestibulo-ocular reflexes (VORs) for head movement toward the hypofunctional side, which causes a blurring of the visual world (i.e., oscillopsia). Many complex changes occur also in the vestibulo-spinal reflexes (VSRs) during head movement (e.g., [23–26]).
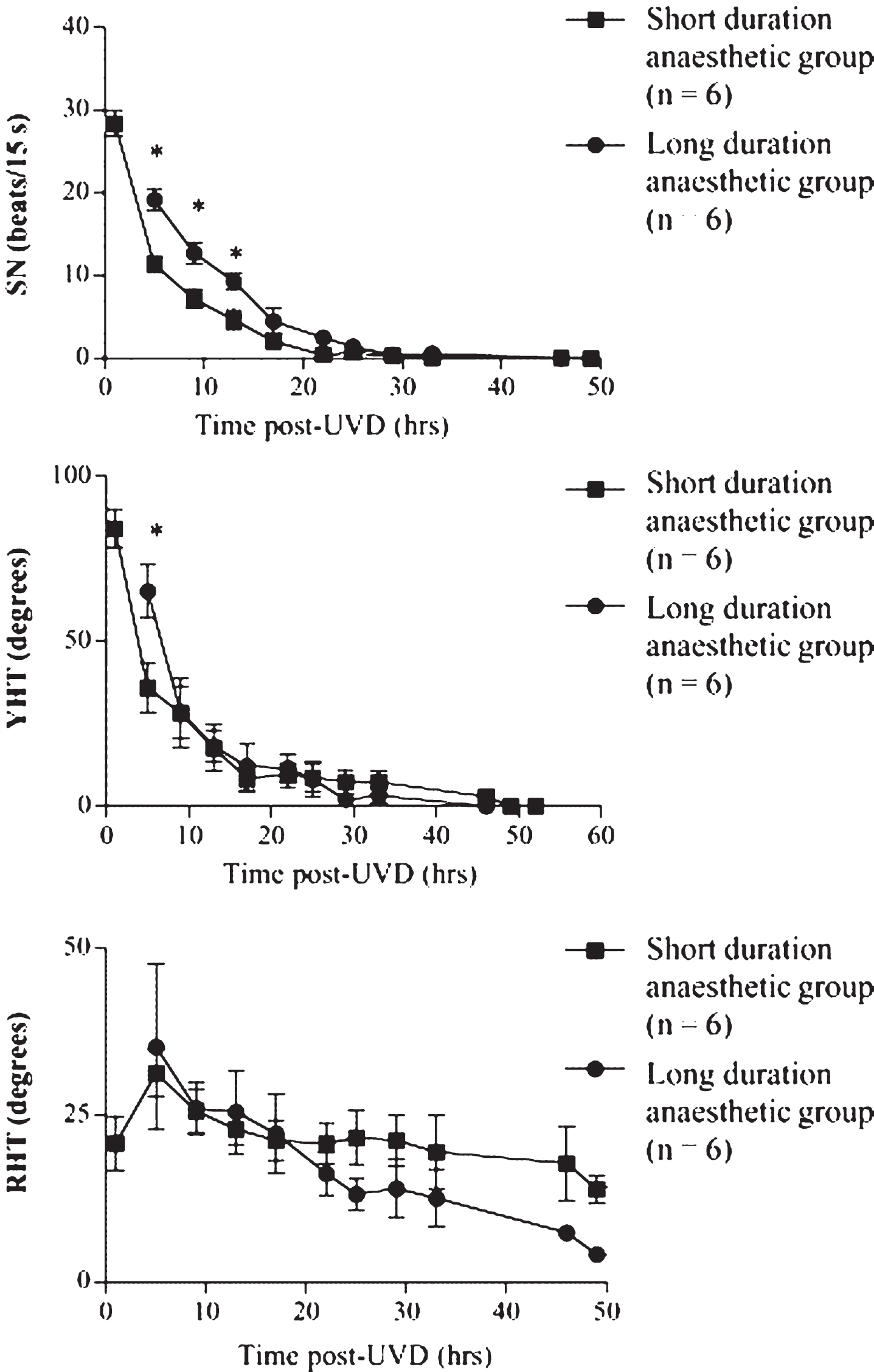
Effects of lengthening the anesthetic duration (from 25 min to 4 h, under isoflurane) on the time course of the spontaneous nystagmus (SN), yaw head tilt (YHT) and roll head tilt (RHT) compensation following unilateral vestibular deafferentation (UVD) in guinea pig. Data are expressed as mean±1 S.D. Reproduced from Gliddon et al. [50] with permission.
There is general agreement that the initial effect of the loss of vestibular input to the VN on the hypofunctional side, is a decrease in the spontaneous activity of those neurons receiving excitatory input from the vestibular nerve (e.g., ‘type I’ neurons in the case of the medial vestibular nucleus (MVN)), although it must be noted that much of the evidence relates to either the MVN or the lateral vestibular nucleus (LVN) and there is less information about the superior and inferior vestibular nuclei (SVN and IVN, respectively) (see [12] for a review). A large number of single neuron recording studies have been conducted in animals and have reported similar results for type I neurons in the MVN (e.g., [17, 27–42]). Similar studies have been published for the LVN (e.g., [43]). This loss of spontaneous neuronal activity is a consequence of reduced excitatory synaptic input from the ipsilateral vestibular nerve as well as increased inhibition from the contralateral MVN, through inhibitory commissural projections, or excitatory commissural projections to inhibitory type II neurons which synapse on type I neurons on their own side [44]. At the same time that the ipsilateral MVN undergoes a large decrease in spontaneous neuronal activity, the contralateral MVN exhibits a dramatic increase, due to release from commissural inhibition normally exerted upon it by type I neurons in the VN on the hypofunctional side. Thus, there is an asymmetry in spontaneous neuronal activity between the ipsilateral and contralateral MVNs (see Fig. 2). However, the LVN does not engage in commissural inhibition to the same extent as type I MVN neurons and therefore changes in the neuronal activity there are due mainly to the loss of ipsilateral excitatory drive from the vestibular nerve (see [12, 45] for reviews). Nonetheless, 2-deoxyglucose (2-DG) studies have still reported an overall asymmetry in activity between the LVNs on the hypofunctional and intact sides [46–49]. After BVL, both VNs undergo a decrease in spontaneous neuronal activity; however, it is less dramatic because both VNs are equally affected by the loss of excitatory input from their ipsilateral vestibular nerve and the impact on the commissural inhibitory pathways is similar bilaterally (e.g., [36]). As a result, the VORs as well as the VSRs are eliminated completely in the case of complete BVL (see [3, 20] for reviews).
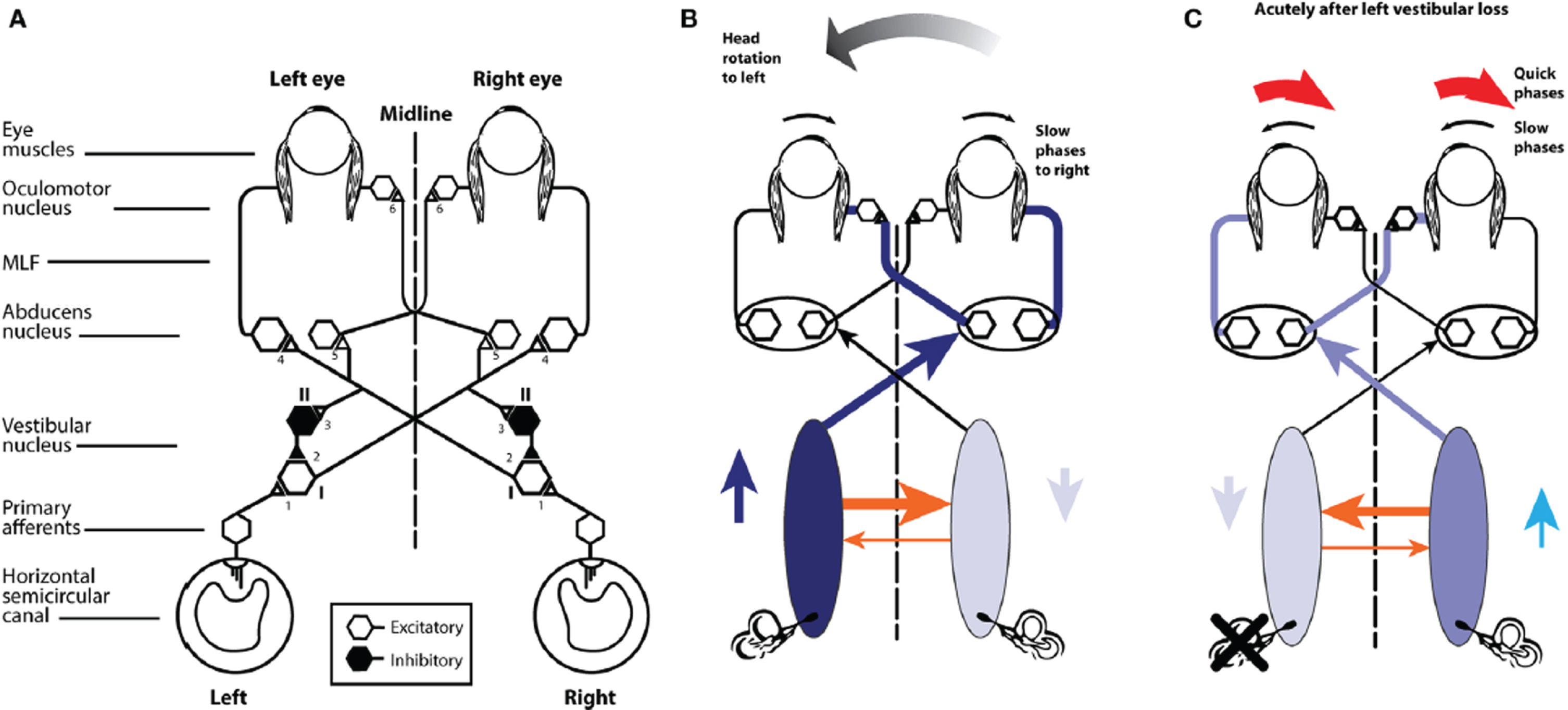
(A) Schematic view of the brainstem showing the basic projections underlying the horizontal VOR. (B) A depiction of the neural activity in the VOR network during a head turn to the left, eliciting a compensatory eye movement response to the right. Increased activation is shown by the darker blue and the thicker orange shows the increased commissural inhibition from the activated vestibular nucleus. (C) With head stationary, a unilateral vestibular loss (left here) elicits an imbalance in neural activity between the two vestibular nuclei. The absence of primary vestibular input means that the left vestibular nucleus has reduced activity (light blue), which in turn generates a reduction in commissural inhibition to the right vestibular nucleus (thin orange line), allowing the cells in the right nucleus to fire at a higher firing rate, resulting in the slow phase of vestibular nystagmus to the left and quick phase to the right (red arrows). From Halmagyi et al. [52] with permission.
Behavioral symptoms such as spontaneous nystagmus and head tilt related to UVL, tend to dissipate rapidly during the process of vestibular compensation, over a time course of days to a week, depending on the species [50] (see Fig. 1). By contrast, the response of the VORs and VSRs to head movement does not recover as rapidly or completely; for example, head impulse testing has indicated persistent deficits in the VOR for rotations to the hypofunctional side (see [3, 15] for reviews). After BVL, there is no longer any vestibular input and therefore, by definition, there can be no pure VORs or VSRs and any reflex response to head movement must derive from some form of sensory substitution [15].
During vestibular compensation for UVL, there is a gradual recovery of spontaneous neuronal activity in the VN on the hypofunctional side. In a seminal study, Precht et al. [27] were the first to report this recovery of spontaneous neuronal activity in the VN ipsilateral to the UVL (in cats). Most of the following studies also reported that there was a regeneration of the spontaneous activity in the ipsilateral VN to a greater or lesser extent, although the specific results varied according to whether, and what kind, of anesthesia was used while recording the neuronal activity. Nonetheless, dynamic responses of ipsilateral VN neurons to head movements remained below normal (e.g., [17, 52]; see [13] for a review of the literature before 1995). Mapping studies using 2-DG are consistent with the recovery of the spontaneous activity reported in single neuron recording studies [46–49]. However, there appears to be a dissociation between the exact time course of behavioral recovery and the restoration of the spontaneous neuronal activity in the ipsilateral VN [34], and therefore other factors may be involved in the rapid compensation of spontaneous nystagmus and head tilt following UVL. Some recent studies even suggest than the efferent vestibular system may be involved [53], although the vestibular nerve afferents on the intact side following UVL are unlikely to contribute substantially to vestibular compensation [54].
In an effort to explain the rapid decrease in behavioral symptoms such as spontaneous nystagmus (see Fig. 1) and head tilt following UVL, McCabe and Ryu [1, 22] proposed the ‘cerebellar shutdown hypothesis’, which is sometimes re-named the ‘cerebellar clampdown hypothesis’. Simply put, the hypothesis is that the cerebellum shuts down the asymmetric spontaneous neuronal activity between the ipsilateral and contralateral VNs, at approximately 1 day following UVL, in order to rebalance the asymmetric spontaneous neuronal activity between the two sides, which is the cause of spontaneous nystagmus and head tilt. This hypothesis which was based on single neuron recordings made in the cat VN by McCabe and Ryu [1, 22], has persisted despite evidence against it, and has re-emerged in clinical contexts as a potential explanation for the different phases of recovery that patients undergo following UVL (e.g., [6–9]).
The cerebellar shutdown hypothesis states that the cerebellum, presumably through its descending inhibitory projections to the VN, inhibits spontaneous neuronal activity in the bilateral VN at approximately 1 day following UVL [1, 22]. The VN ipsilateral to the UVL would be expected to have lower than normal spontaneous activity at this time; however, many single neuron recording studies have recorded in the contralateral VN immediately or up to 24 h following UVL, and none of them has observed electrical silence in that nucleus [31, 55]. Some of these studies have been performed with the cerebellum partially removed (e.g., [31]); however, others have been conducted with the cerebellum intact [33, 54]. Similar results have been reported in alert animals [36].
The case for the cerebellar shutdown or clampdown hypothesis
The primary evidence on which the cerebellar shutdown hypothesis is based is a series of single neuron recording experiments in spinally transected cats with the cerebellum intact. McCabe and Ryu [1] and McCabe et al. [22] recorded at several time points following UVL and reported that at 1 and 2 days post-lesion, “there was no activity (electrical silence) in either the ipsilateral or contralateral MVN” ([22]; p. 385). However, when they removed the cerebellum, they observed an increase in activity in the contralateral MVN. Hence, they concluded that the cerebellum must be “shutting down” activity in the contralateral MVN in an attempt to re-balance the asymmetry in neuronal activity between the bilateral VN.
Given that the cerebellum has a close relationship with the VN, it is not surprising that many studies have documented neurochemical and morphological changes in the cerebellum following UVL and BVL (e.g., [56–74]). Lesions of the flocculus [75–77], and the nodulus and uvula [78], have been reported to retard vestibular compensation, although other studies are inconsistent [79]. There has been conflicting evidence regarding the effects of lesions of the anterior and posterior vermis, and the fastigial nuclei, as well (see [12] for a review). A number of more recent studies suggest that the cerebellum is necessary for normal vestibular compensation of the VORs and VSRs (e.g., [80, 81]). However, while these various studies indicate an important role for the cerebellum in vestibular compensation in general, none of them directly supports the cerebellar shut down hypothesis, because it is not necessary for the cerebellum to shut down activity in the bilateral VN during the early phases of compensation in order to make an important contribution to the compensation process.
The case against the cerebellar shutdown or clampdown hypothesis
A number of studies have demonstrated vigorous neuronal activity in the contralateral MVN within 1 day following UVL; however, some of these recorded immediately after the UVL (e.g., [29, 82]) and many of them involved the removal of the midline cerebellum (e.g., [30] (albino rats); [28, 32]), which could have prevented a cerebellar shutdown. Therefore, the only neurophysiological studies relevant to test the cerebellar shutdown hypothesis are those that kept the cerebellum intact and included neuronal recording in the contralateral VN during the first 24 hrs., but not immediately, following UVL.
Hamann and Lannou [30] studied the responses of VN neurons in the pigmented rat with cerebellum intact during the first 48 hrs. following UVL. The animals were anaesthetized with pentobarbital. Recording in the VN on the side contralateral to the lesion, they found that the spontaneous neuronal activity of type I neurons was not significantly different to animals with intact labyrinths, indicating no evidence of a cerebellar shutdown.
In a direct test of the cerebellar shutdown hypothesis, Curthoys et al. [55] demonstrated that the spontaneous neuronal activity in the contralateral MVN underwent a dramatic increase immediately following UVL, returning toward normal values by approximately 12 hs. Removal of the midline cerebellum further increased the spontaneous activity but with the cerebellum intact there was still an increase in firing rate following UVL. These experiments were conducted in guinea pigs anaesthetized with pentobarbital, fentanyl and droperidol, and in 2 animals a spinal transection was performed to allow them to recover from surgical anesthesia and be maintained on local anesthesia. A total of 612 neurons were recorded in the contralateral MVN before, immediately after, and at 12 hrs. following UVL, with the cerebellum intact, and 429 neurons were recorded with the midline cerebellum removed. Figure 3 shows the time course of the changes in spontaneous neuronal activity for each condition. In order to confirm that the presence of neuronal activity in the contralateral MVN was in no way attributable to the anesthetics used in the main part of the study, spinal transection was employed in 2 animals, and the results were similar.

Spontaneous neuronal activity for type I neurons in the contralateral MVN in anaesthetized guinea pigs with the cerebellum intact or the midline cerebellum removed in labyrinthine intact animals (‘before’), animals immediately after and up to 12 hs following a UVL, and animals >12 hs following a UVL. Columns show means±95% confidence interval. Redrawn from Curthoys et al. [55] with permission.
Newlands and Perachio [33] performed a similar study in the gerbil following UVL. The animals were anaesthetized with a short-acting barbiturate anesthetic (methohexital sodium) and then decerebrated. Recordings were made in the contralateral MVN from 2 hrs. following the UVL. They observed vigorous type I neuronal activity in the contralateral MVN, with an average firing rate of 44.6±25.5 spikes/sec.
Furthermore, in alert guinea pigs with cerebellum intact, Ris and Godaux [36] recorded from neurons in the contralateral SVN, MVN, LVN and IVN, at 1, 6, 24, 48 h and 1 week following UVL. Consistent with the previous studies, they observed spontaneous neuronal activity at all time points, including resting rates above 40 spikes/sec for type I neurons (see Fig. 4). This was incontrovertible evidence against the cerebellar shutdown hypothesis, because the presence of spontaneous neuronal activity in the contralateral VN could not be dismissed as an artifact of anesthesia, removal of part of the cerebellum or even a restriction of neuronal recording to the MVN.
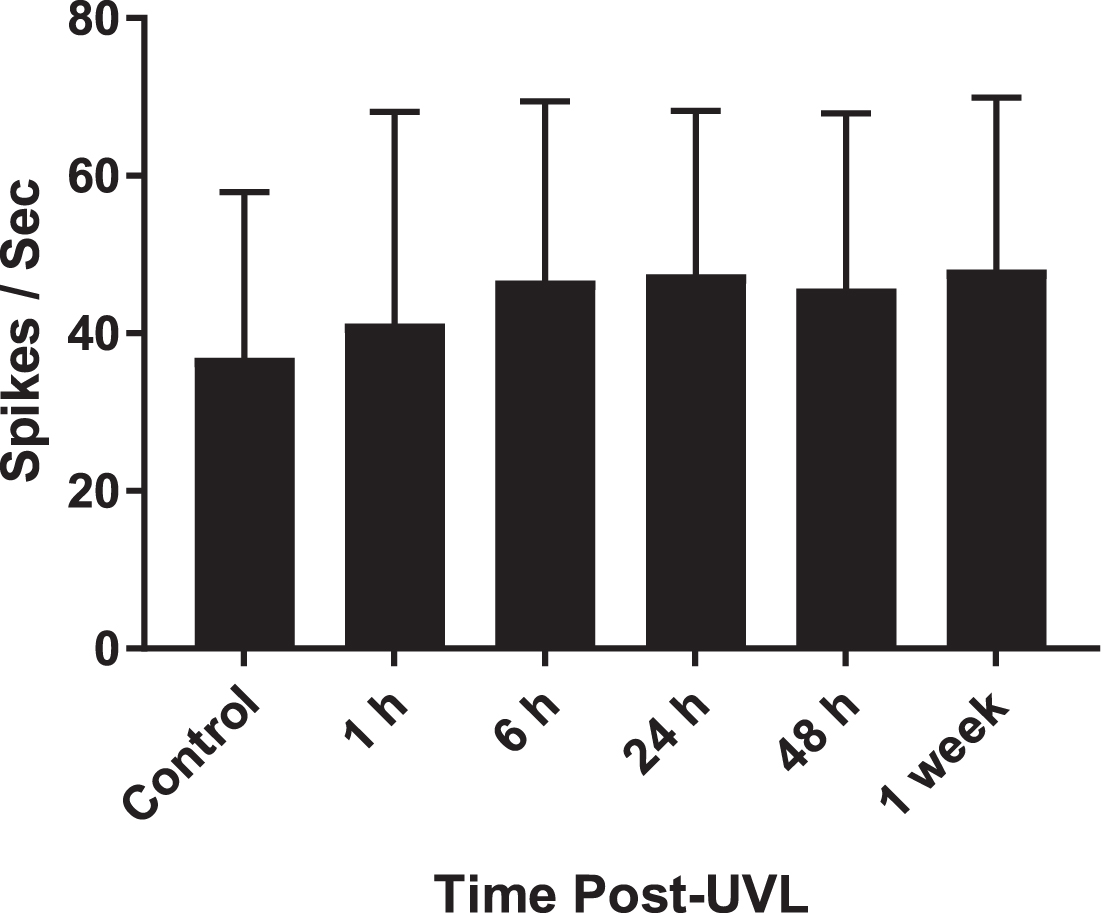
Spontaneous neuronal activity for neurons in the contralateral VN in alert guinea pigs with the cerebellum intact. Columns show means±1 SD. Samples sizes are: 261, 105, 105, 109, 98 and 90 neurons, respectively. Redrawn from Table 2 in Ris and Godaux [36] with permission.
Studies of the bilateral VN following UVL, using 2-DG, PET or MRI, also do not support the cerebellum shutdown hypothesis [46–49, 83–85]. They demonstrate asymmetric activity between the bilateral VN during the first 24 hrs. following UVL.
Allum et al. [86] estimated that approximately 60% of the VOR improvement following UVL is mediated by central compensation processes, which reduce the asymmetry in spontaneous neuronal activity between the ipsilateral and contralateral VNs. If the cerebellum were to shut down the activity in the bilateral VNs following UVL, then it seems logical that there would be no spontaneous nystagmus or head tilt during this time (see Fig. 1 for evidence), because the state of the bilateral VNs would be more similar to a BVL.
There is overwhelming evidence against McCabe and Ryu’s ‘cerebellar shutdown/clampdown hypothesis’ of vestibular compensation and very little evidence in support of it. The direct evidence supporting it are two related studies by the authors in spinally transected cats, the results of which have never been replicated. The direct evidence against it consists of 4 single neuron recording studies by different researchers, using guinea pigs, rats or gerbils, who obtained similar results in anaesthetized, decerebrate or alert animals with the cerebellum intact. Numerous 2-DG, PET and MRI studies are all consistent with the latter evidence. Furthermore, the hypothesis is not logical: if the cerebellum did shut down activity in the bilateral VN within 1 day following UVL, there would be no spontaneous nystagmus or head tilt, because there would be no asymmetry in activity between the bilateral VNs. This is clearly not observed at this stage of vestibular compensation following UVL (see Fig. 1; see [12] for a review). The cerebellar shutdown/clampdown hypothesis is inconsistent with neurophysiological evidence and it is potentially misleading to physical therapists, because it suggests that early in the process of vestibular compensation, the brain has undergone neurophysiological changes to ease the asymmetry in neuronal activity in the VNs caused by UVL, when in fact this is a time when UVL symptoms may be at their worst. This may lead to errors in suggested vestibular rehabilitation programs for a patient with UVL at this stage of recovery and is potentially dangerous. I propose, therefore, that the cerebellar shutdown/clampdown hypothesis of vestibular compensation should now be considered mainly of historical interest.
Footnotes
Acknowledgments
I would like to thank Dr. Michael Maslin and Professor Ian Curthoys for useful discussions.