
Abstract
The mechanisms of ion exchanges and water fluxes underlying the endolymphatic hydrops phenomenon, remain indeterminate so far. This review intends to reposition the physical environment of the endolymphatic compartment within the inner ear, as well as to recall the molecular effectors present in the membranous labyrinth and that could be at the source of the hydrops.
Prosper Menière was the first to attribute the source of the disease that now bears his name, which is characterized by paroxysmal vertigo, tinnitus and deafness, to the inner ear [29]. However, it was not until 80 years later that histopathological studies were able to correlate these functional deficits with tissue damage. The swelling of the endolymphatic compartment, which is associated with atrophy of both the cochlea and vestibule, suggests a pathophysiological mechanism centred on endolymphatic hydrops [14].
If the involvement of ion exchange and water flux between the different inner ear compartments underlies the hydrops phenomenon, the cellular and molecular mechanisms that contribute to its generation remain undetermined thus far. It is thus opportune to reposition the physical environment of the endolymphatic compartment within the inner ear and to remind the molecular effectors present in the membranous labyrinth potentially at the source of hydrops.
What is known about the endolymph
The endolymph is a liquid rich in potassium ions (K+) that is common to the cochlea and vestibular organs, as it is found in the scala media and all vestibular cavities. Due to its particular ionic composition, it is essential for the process of mechanoelectrical transduction (MET) by hair cells [15]. As such, its ionic composition must be very finely tuned. Other ionic exchanges involving Na+, Ca2 + and Cl- ions also occur in different parts of the membranous labyrinth. It can be assumed that alterations in the composition and/or hydrostatic pressure of the endolymph can affect the generation of vestibular sensory information and subsequently vestibular function.
Endolymph at the interface between two fluid exchange compartments
The endolymphatic compartment is located at the interface between two fluid exchange compartments. The first is the blood compartment, which is separated from the endolymph by the blood-labyrinthine barrier (BLB). The second is the perilymphatic compartment, which is separated from the endolymph by the membranous labyrinth wall (MLW).
The structure and properties of the BLB have been well studied in the cochlea. The stria vascularis is composed of a complex multilayer epithelium through which controlled ion and water exchanges take place [35]. These exchanges ensure the maintenance of the endolymph high K+ concentration despite continuous variations in blood composition. Significant alterations in BLB permeability have been shown to occur under conditions of infection or inflammation [44]. Bacterial infection of the middle ear, for example, is accompanied by significant bacteriolysis and the release of lipopolysaccharide, which modify BLB permeability directly or through the activation of macrophages and the release of inflammatory cytokines. Alterations in BLB permeability have also been reported in cases of sound stimulation [43].
Contrary to what is generally believed, the MLW does not behave like a long impermeable tube. In con-trast, this membrane is composed of a multilayer epithelium [38] in which different types of ion channels and membrane receptors are expressed. Although identifying molecular effectors that can effectively modify the ionic composition of the endolymph is critical, data regarding the types and precise distribution of membrane channels and receptors expressed at the MLW are scarce.
Thanks to the work of D Marcus and P Wangemann from the Kansas State University, it is known, that different types of ions are exchanged through the MLW [23]. For example, Na+, Ca2 + and Cl-, like K+, are continuously exchanged between the endolymph and perilymph. The efflux of Na+ through epithelial sodium channels was measured in extramacular epithelial cells of the saccule, Reissner’s membrane and endolymphatic sac [22]. An influx of Cl- into the endolymph also occurs under the combined influence of ATPase Na-K pumps, which allow the accumulation of K+ in epithelial cells of the MLW, and of Cl- channels, which allow an influx of Cl- into the endolymph [20, 21–30].
A significant amount of water is also transferred between the endolymphatic and perilymphatic compartments through the MLW. This water transfer takes place through aquaporins (AQs), specific water transport proteins [7], and intercellular space by ionic gradient. We now know that different types of AQs (AQ1 to AQ4) are expressed in different areas of the membranous labyrinth. They have been found in sensory cells, support cells, dark cells, Scarpa’s ganglion and the endolymphatic sac [17, 40]. AQ4 is also expressed along the cochlear duct in the inner sulcus cells, Hansen and Claudius cells [12]. As AQs open, water freely diffuses through the MLW as a function of the ion concentration gradient.
Potential cause of endolymph alteration
Different factors may modulate the ionic transport in the endolymph and subsequently trigger water exchange between liquid compartments adjacent to the inner ear, eventually leading to hydrops. Dysregulation of water exchange between cerebrospinal fluid (CSF) and perilymph, inflammatory foci in the membranous labyrinth, ischaemia-induced acidification of the stria vascularis, or sound stimulation as cited above, and alterations in endolymph drainage upon dysfunction of the endolymphatic sac, are among the multiple factors that may lead to a perturbation of the inner ear ion homeostasis. Modification of the ionic transport between the inner ear liquids under inflammatory conditions has also been well documented [24, 25–38]. Endocrine fluctuations in the vascularization zone of the inner ear are also thought to perturb ion exchange in both the vestibule and cochlea, and thus contribute to the generation of hydrops. Indeed, different types of hormone receptors have been shown to be expressed in the inner ear organs. This is the case for serotonin [1, 39], histamine [32, 37], adrenergic [8, 27], vasopressin [40, 41], and oestrogen receptors [28]. These different types of hormone receptors are found in different areas of the membranous labyrinth, such as the stria vascularis, the endolymphatic sac, the sensory epithelia, and the MLW. Functional studies have also provided evidence that some of these hormones directly impact physiological parameters such as electrogenic transport [19] or endolymph volume [16]. A better understanding of the cause-and-effect relationship between ion transport and hydrops would benefit from determining the precise localization of the hormone receptors and ion channels expressed at this level and their colocalization with the water transport proteins.
Relation between endolymph hydrostatic pressure and mechano-electrical transduction
Hydrostatic pressure in the endolymph results from the equilibrium of ion concentrations on either side of the MLW, i.e., between the endolymph and perilymph, and from water exchange through this wall. To better understand the notion of hydrostatic pressure and its consequences on the physiology of the vestibule, it is necessary to return to the hydrostatic and hydrodynamic theories proposed nearly 150 years ago by Friedrich Goltz and Joseph Breuer, two famous European physiologists [5, 13] (Fig. 1). These two theories are of foremost importance for understanding the mechanisms of cupula displacement during head acceleration. The hydrodynamic theory suggests that as the endolymph is able to move within the membranous labyrinth, it actively displaces the cupula and triggers the mechanoelectrical transduction (MET) process. Conversely, due to liquid volume and semi-circular canal geometry, the hydrostatic theory refutes the hypothesis of liquid displacement inside the membranous labyrinth. In this theory, head acceleration is essentially transmitted to the hair cells through the movement of the cupula and otoconia membrane, which act as inertial masses. The weight of the water column has a limiting action on the displacement of the cupula and as such may impact the triggering characteristics of the MET. Although Josef Breuer’s hydrodynamic theory is currently favoured in the field of otoneurology, the hydrostatic theory deserves to be considered, as it could explain the different pathophysiological mechanisms underlying vestibular disorders such as BPPV and Meniere’s disease.

In the early stages of alterations in the hydrostatic pressure of the endolymph, mechanical stress may be applied to the mechanotransduction apparatus. This mechanical stress in turn modifies the frequency range of cupula displacement and alter the electrical message in vestibular nerve fibres. In the case of higher increases in pressure, it can be assumed that mechanical damage, which can be visualized today by MRI imaging of hydroptic ears, may take place. The histopathological work carried out by our peers for 150 years [9, 33] has allowed us to understand the consequences of endolymphatic hydrops in the human inner ear. At the most advanced stages of Meniere’s disease, dilation of the scala media along with rupture of Ressner’s membrane are pronounced in the cochlea. In the macula, as well as in the ampulla, significant enlargements of the endolymphatic compartment associated with degeneration of sensory epithelia have been demonstrated [14]. These structural damages were recently confirmed by MRI imaging approaches [3]. Many clinical observations have confirmed the existence of hydrops and its concomitance with Meniere’s disease. However, the impact of slight modifications of the hydrostatic pressure of the endolymph remains much less documented so far.
Physical damage is not needed to explain hydrops-related functional alterations
If we consider the movement mechanics of the cupula in the endolymphatic space, we can assume that the free movement of the cupula requires a balance of hydrostatic pressure on both sides of the ampulla [42]. Considering that the columns of liquid in the semi-circular canals on either side of the ampulla have similar masses, the cupula, which acts as an inertial mass, may be displaced to approximately the same extent on either side of the ampulla upon head acceleration (Fig. 2). A slight change in hydrostatic pressure on one side of the ampulla may be sufficient to alter the movement of the cupula and consequently the generation of the vestibular information. This is what happens in the case of dehiscence of the upper semi-circular canal (SCC), which leads to Minor syndrome. In this situation, the opening of a window in the bony wall of the upper SSC causes a change in hydrostatic pressure in the SSC [31]. This configuration, in which the hydrostatic pressure on either side of the cupula is no longer balanced, leads to alterations in vestibular information, and subsequently the generation of a vestibular syndrome. This is also what happens during surgical plugging of the SCC in cases of Meniere’s disease. Obstruction of a deficient canal reduces the working frequency of the ampulla. This operation provides therapeutic benefits, reducing acute episodes of Meniere’s disease [11].
Taking into account all the structural characteristics of the vestibular sensors mentioned above, as well as their mechanical properties within the membra-nous labyrinth, it can be assumed that local perturbations in the ionic composition of the endolymph (e.g. hormonal alterations mentioned above), associated with structural specificities of the SCC (e.g. local reduction or partial obstruction of the SCC lumen), may lead to alterations in the generation of the vestibular information unilaterally, and subsequently conduct to vestibular syndromes such as encountered in BPPV and Meniere’s disease. The formation of domains of altered hydrostatic pressure in the SCC would thus modify the displacement of the inertial masses, alter the generation of vestibular sensory information, and trigger a vertiginous syndrome. The reality of the formation of such domains of altered hydrostatic pressure remains to be confirmed. Nevertheless, the physical phenomena that govern ion and fluid exchange between the CSF and perilymph and the endolymph do not prohibit such a process.
What do the experimental hydrops models bring us?
Various animal models of endolymphatic hydrops have been developed over the last 30 years, with diverse outputs (for review [34]). The injection of artificial endolymph into the scala media, aiming at increasing the endolymph volume, induced physiologic changes (e.g. changes in endocochlear potential) linked to the pressure changes due to the injection, rather than from a sustained increase of the endolymph volume. Combined surgical ablation of the endolymphatic sac and vasopressin injection, aiming at inducing endolymphatic hydrops, exacerbated vestibular symptoms in a guinea pig model [18]. Conversely, the surgical approach to the endolymphatic sac and the cochlear aqueduct in the guinea pig did not resulted in the generation of vertigo [2]. In the same way, the administration of aldosterone, which is thought to act on the reabsorption of Na+ and Cl- and in turn on water retention, or the administration of vasopressin, which acts on aquaporins, do not have a significant effect on the generation of vertigo in animals [26].
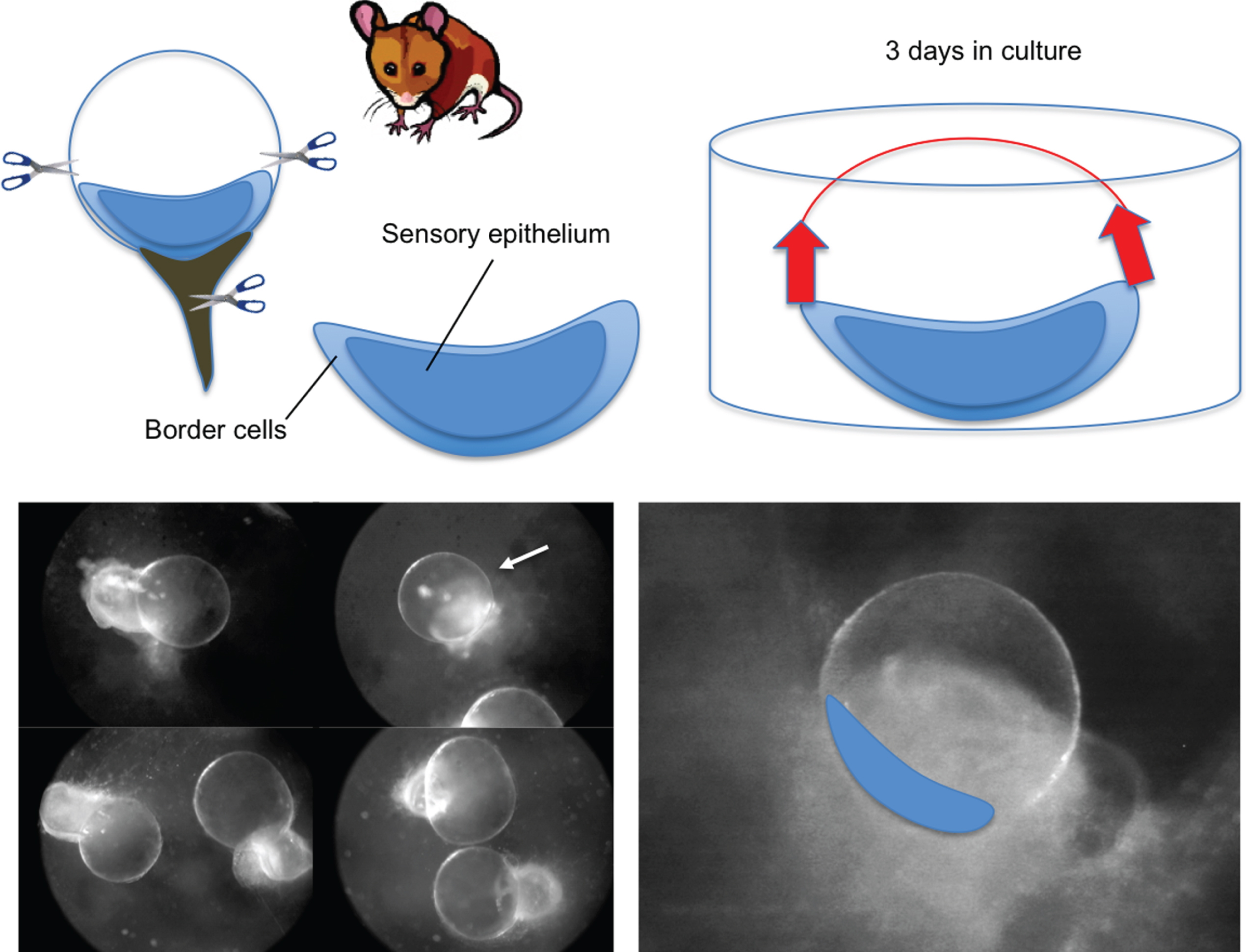
Other models have allowed a better understanding of the dynamics of ion exchange in the endolymph, its correlation with water movement and control of the hydrostatic pressure of the endolymph. This is the case for the endolymphatic cysts formed following the removal of the vestibular macula in rodents [4, 10]. In this model, the utricle is removed from young rats or mice and cultured for several days. After two to three days in culture, the border cells display regenerative properties that allow rebuilding of the membranous labyrinth. When performed at neonatal stages in rats and mice, this operation leads to the formation of a significant number of cysts. After three days in culture, the cysts are fully reformed. Measurements of the K+ concentration in the endolymph using ion-selective electrodes have shown that the mean concentration of 80 mM K+ is reached after four days in culture [44].
This high K+ concentration, which is maintained over a period of approximately two weeks, results from the spontaneous accumulation of K+ in the endolymphatic compartment. It is accompanied by the swelling of cysts as a consequence of water in-flux through the MLW. Although the hydrostatic pressure inside the cysts was not measured in this study, the pharmacological blockade of K+ influx and efflux demonstrated that K+ secretion by dark cells is highly efficient. Indeed, at a saturating concentration of ouabain, a specific blocker of the Na+-K+ ATPase pump expressed at the base of the dark cells, the K+ concentration in the cysts drops by 50% in 25 minutes. This observation is noteworthy considering that a 1-millimolar change in the K+ concentration in the vicinity of transduction channels, which are located in the hair bundle of vestibular sensory cells, is sufficient to significantly alter vestibular sensory information [3]. The pharmacological blockade of other effectors also expressed in dark cells, such as the Na-K-Cl type-1 cotransporter (NKCC1) using micromolar concentrations of bumetanide, also induces a very significant decrease in the endocystic K+ concentration within 15 minutes. It should also be noted that the blockade of K+ effluxes through transduction channels has a significant impact on the endocystic concentration of K+. In the presence of gentamicin or amikacin, two antibiotics administered in the case of middle ear infections, or in the presence of FM1-43 or gadolinium in the culture bath medium, a 30% increase in the endocystic K+ concentration is observed after 60 minutes. These data directly confirm the K+ efflux through hair cells. They also provide details on the likely ototoxic mechanisms of aminoglycosides. The development of vestibule explants that preserve synaptic contacts with vestibular nerve fibres may eventually lead to a better understanding of how the alteration of the K+ cycle or the changes in the hydrostatic pressure of the endolymph can affect the excitability of the vestibular nerve. Mathematical modelling of cupula movements as a function of hydrostatic pressure changes will also need to be undertaken.
Conclusion
Different studies need to be undertaken in the near future to better understand both the mechanisms of the development of endolymphatic hydrops and its consequences on vestibular pathophysiology. Identifying the molecular effectors expressed in the membranous labyrinth and understanding their possible involvement in the hydrops phenomenon is a priority. It is also important to better visualize hydrops in humans to understand its characteristics, sequence of development, and correlation with the manifestation of the different symptoms of vertigo. The identification of biomarkers of the various types and stages of Meniere’s disease is also required to develop targeted and effective countermeasures.
Conflict of interest
The author declares no competing financial interests.