
Abstract
BACKGROUND:
The gain (eye-velocity/head-velocity) of the angular vestibuloocular reflex (aVOR) during head impulses can be increased while viewing near-targets and when exposed to unilateral, incremental retinal image velocity error signals. It is not clear however, whether the tonic or phasic vestibular pathways mediate these gain increases.
OBJECTIVE:
Determine whether a shared pathway is responsible for gain enhancement between vergence and adaptation of aVOR gain in patients with unilateral vestibular hypofunction (UVH).
MATERIAL AND METHODS:
20 patients with UVH were examined for change in aVOR gain during a vergence task and after 15-minutes of ipsilesional incremental VOR adaptation (uIVA) using StableEyes (a device that controls a laser target as a function of head velocity) during horizontal passive head impulses. A 5 % aVOR gain increase was defined as the threshold for significant change.
RESULTS:
11/20 patients had >5% vergence-mediated gain increase during ipsi-lesional impulses. For uIVA, 10/20 patients had >5% ipsi-lesional gain increase. There was no correlation between the vergence-mediated gain increase and gain increase after uIVA training.
CONCLUSION:
Vergence-enhanced and uIVA training gain increases are mediated by separate mechanisms and/or vestibular pathways (tonic/phasic). The ability to increase the aVOR gain during vergence is not prognostic for successful adaptation training.
Introduction
In the last decades two main semi-circular canal vestibular afferent pathways have been identified (for review see [21]). A tonic pathway, the properties of which resemble those of regularly discharging afferents, conveys a signal corresponding to head velocity over a broad range of rotational frequencies and vel-ocities with linear response properties. A phasic pathway with properties resembling those of irregularly discharging afferents is sensitive to angular acceleration (as well as velocity) as the frequency of rotation increases. This latter pathway is highly modifiable and very sensitive to inhibitory cut-off. Experimental studies using animal models suggest that after chronic unilateral labyrinthectomy, the gain (eye velocity/head velocity) response from both pathways are increased for contra-lesional rotations, but the phasic pathway is modified to a greater degree [13, 14].
It is well known that the gain of the angular ves-tibulo-ocular reflex (aVOR) is greater than unity when viewing near targets [15]. This vergence-med-iated aVOR gain increase depends on the highly modifiable phasic pathway of the aVOR as observed in patients treated with intra-tympanic gentamicin [17]. Gentamicin preferentially damages type I peripheral vestibular hair cells and the irregular afferent pathway, thereby abolishing the vergence-mediated aVOR gain increase during high acceleration/high velocity head impulses [17]. Additionally, in patients with semicircular canal plugging surgery that spares hair cell function, the vergence-mediated aVOR gain increase is preserved putatively due to the spared irregular afferents [18] although the aVOR gain is still reduced. The main drive for the vergence-mediated gain change is thought to be a combination of binocular position and accommodation cues [3]. In addition to enhancing the aVOR gain through vergence, it has recently been shown that the aVOR gain during high acceleration head impulses can be increased using a unilateral incremental VOR gain adaptation method (uIVA) that uses an incremental retinal image velocity error signal (for review [26]). The retinal error signal is the likely main drive for aVOR gain adaptation, i.e., a drive different to that for vergence-mediated gain increase. This technique may be important as a tool to improve the ipsi-lesional aVOR gain in patients with vestibular hypofunction resulting from, for example, vestibular neuritis [1].
It is not clear which of the two afferent pathways are predominantly responsible for the gain increase during such training. If we can determine which pathway contributes to the motor learning engendered by the uIVA method, then we are likely to develop better rehabilitative interventions for patients with unilateral vestibular hypofunction. For example, it may be possible to preferentially stimulate the efferent vestibular system, which modulates afferent vestibular signals, thereby increasing the ratio and sensitivity of the irregular afferents [9, 10].
In this experiment, we ask whether there is a correlation between the two mechanisms of increasing aVOR gain (vergence and adaptation). Specifically, we seek to know whether the aVOR gain increase due to uIVA training correlates with the vergence-med-iated gain increase in patients with unilateral vestibular hypofunction (UVH) after vestibular neuritis. We hypothesise that the degree of positive correlation between these two mechanisms will reveal whether or not these two mechanisms share an origin.
Methods
Subjects
Twenty patients (13 men, 7 women) with UVH due to vestibular neuritis were tested. Thirteen patients had left UVH and n = 7 patients had right UVH. In 50% of patients (n = 10), only the superior and lateral canals were involved, whereas the other patients had reduced aVOR gains in all three ipsi-lesional semicircular canals as per clinical video-oculography measures (ICS System, GN Otometrics, Denmark). In the present study, only horizontal head rotation (i.e. predominantly horizontal semicircular canal) was studied. The average age of the patients was 50.45±8.04 years (range 34–64 years). The average time after the acute vestibular loss was 19.00±14.45 months (range 6 months - 60 months). In eight cases, 20 months or more passed since the acute phase of the vestibular neuritis. The criteria for diagnosis of vestibular neuritis, and inclusion into the study, were the presence of typical clinical signs such as spontaneous nystagmus, increased nystagmus velocity during gaze in the direction of the fast phase, VOR gain less than 0.8 on the involved side, and a normal contralateral VOR gain [2, 27]. Additional inclusion criteria were normal hearing (hearing tests showing no deviation of hearing threshold relative to age-matched controls) and no middle ear pathology (via eardrum examination). Exclusion criteria were gaze evoked nystagmus and skew deviation.
The study was approved by the institutional review board of the Petz Aladar Hospital, Györ, Hungary (PAMOK Hospital Protocol number 76-1-13/2018). The protocol was in accordance with the ethical standards laid down in the 1964/2013 (7th revision) Declaration of Helsinki for research involving human subjects and the investigators obtained written informed consent from each participant.
Recording system
Head and eye rotations were measured using a monocular video oculography recording system, EyeSeeCam system (Interacoustics, Denmark). The EyeSeeCam system has two camera mounting locations, allowing either the right or left eye to be rec-orded. Horizontal aVOR gain measurements were performed during convergence using the monocular video technique as described in [6].
Experimental protocol
Each subject was seated 1.5m in front of a featureless white wall during far-viewing head impulses. First, the dominant eye was determined using the unconscious sighting task [23]. During the experiment the dominant eye was measured during baseline gain and post training gain testing (explained below), whereas both eyes (not simultaneously) were measured for vergence gain testing (explained below) as per Figtree et al. [6] (Fig. 1). Subjects always used both eyes to view the target during testing and training.Two targets, one far (1.5 m) and one near (15 cm), were placed directly in front of each eye (separated by the subject’s IPD) and positioned at eye level. While looking at either near target, one eye was straight-ahead (close to primary position) and the other eye was adducting. Only the adducting eye was recorded with video-oculography. Subjects were as-ked to close their adducting eye and verify that the line-of-sight for their straight-ahead eye intersected both the near and far targets on the side of that eye, i.e. left near and left far targets when the left eye was straight-ahead and vice versa. This target alignment process was performed between each head impulse [3]. At the beginning of each trial an in-vivo calibration was performed by recording eye position while subjects fixated the far-targets for ten seconds each. These fixation points were used to determine the reference (zero) position of the adducting eye during near-viewing. Passive far- and near-viewing horizontal aVOR gains during head impulses were recorded as separate trials consisting of 10 head impulses in each direction. The vergence-mediated gain enhancement was considered significant when aVOR gain was ≥5% of far-target viewing gain.
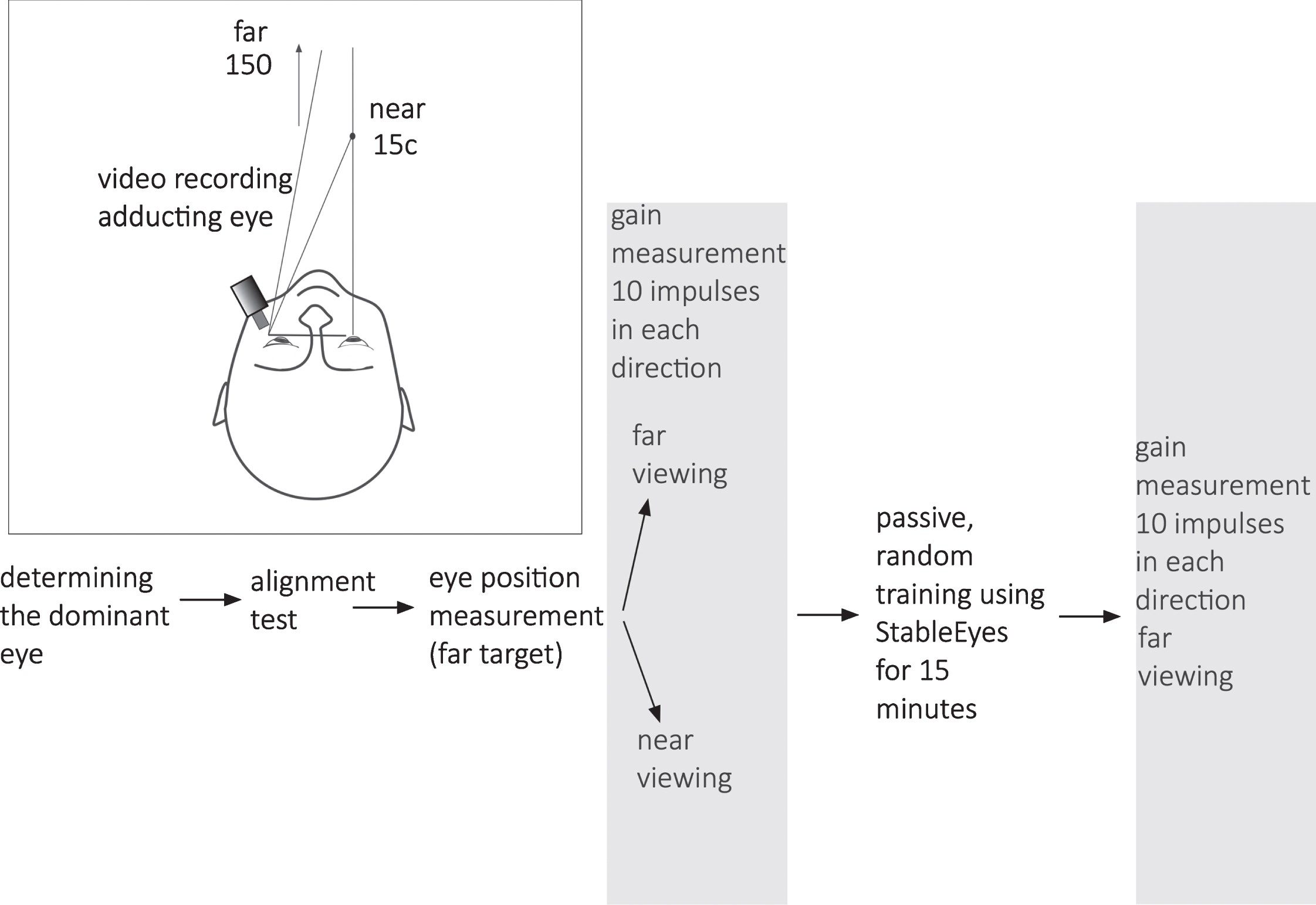
Experimental setup and data collection sequence.
Each subject then performed 15 minutes of uIVA passive training [5, 29] using StableEyes [28]. Eye movements were viewed by the operator, but not recorded to file during training. The training consisted of horizontal passive head impulses delivered by one of two experienced operators (operators alternated between subjects). Head impulses were unpredictable in timing and direction and delivered randomly left and right. During each head impulse, the subject’s task was to visually follow a moving laser target. Apart from the laser target, training was performed in darkness. The movement of the laser target was set depending on the desired gain demand. For example, if the desired gain demand was 1.5 the laser moved in the opposite direction to the head, but at 50% of head velocity, requiring an aVOR gain of 1.5 to stabilize the laser target image on the retina. During adaptation training the gain-demand towards the ipsi-lesional side increased by 0.1 every 90 seconds, starting from the subject’s pre-training far-viewing aVOR gain (rounded up to the nearest 0.1, as reported by EyeSeeCam). Towards the contra-lesional side the gain-demand was always 1.0. Based on prior work, gain increases of 3-4% are typical for rotations towards the side not being trained and are often statistically insignificant [22], thus uIVA gain adaptation ≥5% was considered as the threshold for adaptation to have occurred as a result of training. A sub-analysis was performed comparing patients below the 5% adaptation threshold with those on or above the threshold. If there is a relationship between vergence-mediated gain increase and VOR gain adaptation, then the vergence-mediated gain increase should be different between these two groups.
Immediately following uIVA training, the far viewing aVOR gain in response to passive head impulses (10 in each direction) were collected as previously described.
Calibrated eye and head data were digitally filtered with a 50-tap zero-phase low-pass FIR filter with a bandwidth of 50 Hz. Horizontal angular eye position was differentiated and the onset of each head impulse was calculated by fitting horizontal angular head velocity magnitude to a polynomial curve versus time. The point where the magnitude of the fitted curve was greater than 2 % of the curve’s peak magnitude (typically this threshold was 4 °/s) was defined as the impulse onset. Only head impulses with peak magnitude between 150°/s and 300 °/s were included in the analysis. Traces with saccades occurring inside a window starting 100 ms before impulse onset and ending at impulse peak magnitude (typically 100 ms after onset) were also removed. Head velocity traces that did not fit the velocity criteria and eye traces containing blinks and other artefacts were removed, along with their corresponding eye and head traces. The instantaneous aVOR gain was calculated as the magnitude of eye velocity divided by head velocity. The impulse aVOR gain was calculated as the median of the instantaneous aVOR gains calculated during the 30 ms period (at 220 Hz this corresponds to six instantaneous gain values) immediately prior to impulse peak-velocity magnitude.
The convergence angle measured immediately prior to each head impulse was calculated as the mean magnitude of the horizontal adducting eye position in the 100 ms window starting 150 ms before head impulse onset. This convergence angle (during near-viewing) was calibrated by subtracting from the adducting eye the horizontal eye position (for that eye) when looking at the far target in front of it. The relevant eye fixation was identified manually, and a time window (which excluded blinks and other artefacts, minimum 2 seconds) was selected over which to calculate the mean horizontal eye position.
Power analysis revealed that eight subjects would be sufficient to detect a difference of 5% in aVOR gain (Mean 1 = 0.6, Mean 2 = 0.63, SD = 0.03, Power = 0.82). Statistics were completed using IBM SPSS Statistics for Windows V.26 software. The significance of a gain increase was calculated using a linear mixed model (LMM) with repeated measures. The independent factors were: state (‘baseline’, ‘vergence’,’training’), side (‘ipsi-lesional’, ‘contra-lesional’), and type (‘adapter’, ‘non-adapter’). The dependent variable was VOR gain. Tukey HSD was used to correct for multiple pairwise comparisons. The Chi-square test was used to compare proportions and linear regression was used to measure correlation between two variables.
Results
There was a significant difference in gains bet-ween baseline and vergence (near viewing) states (LMM: F1,36 = 20.432, p < 0.001) and between sides (LMM: F1,36 = 68.612, p < 0.001). In addition, there were significant interactions between side and type (LMM: F1,36 = 4.618, p < 0.05), suggesting that th-ere was a difference in gain between adapters and non-adapters for ipsi-lesional rotations; and an inte-raction between state, side and type suggesting that that difference between groups for ipsi-lesional rotations became larger during vergence (LMM: F1,36 =6.424, p < 0.02). We observed the same significant differences and interactions between baseline and post training (state: LMM, F1,36 = 20.79, p < 0.001; sides: LMM, F1,36 = 58.8, p < 0.001; state*type: LMM, F1,36 = 4.175, p < 0.05; side*state*type: LMM, F1,36 = 14.048, p < 0.002). When comparing the gain increase due to vergence (i.e., vergence gain minus baseline gain) with the gain increase due to training (post training minus baseline), there was no difference (LMM, F1,36 = 2.455, p = 0.126). However, there was a significant interaction between side and type (side*type: LMM, F1,36 = 12.35, p < 0.001), suggesting that there was a difference in ipsi-lesional gain increases due to vergence and training between adapters and non-adapters.
aVOR gain (baseline)
The mean vHIT aVOR gain (±SD) for all patients during ipsi-lesional and contra-lesional yaw head rotations during far viewing were 0.47±0.2 (n = 193 impulses) and 0.98±0.11 (n = 143 impulses) respectively. During near viewing, the vergence angle was on average 14.49±4.23 degrees during vHIT towards the ipsi-lesional side, and 15.95±5.87 degrees towards the contra-lesional side.
aVOR gain enhancement (vergence and uIVA)
During near viewing, the average vHIT gain inc-reased by 15% to 0.542±0.24 (mean±SD) during ipsi-lesional yaw head rotations; for contra-lesional yaw head rotations the aVOR increased 10% to 1.08±0.14 (mean±SD).
After uIVA training, the mean vHIT aVOR gain increased by 14% to 0.536±0.24 during ipsi-lesional rotations and by 6% for contra‐lesional rotations (1.04±0.11), (Table 1, Fig. 2).
Mean (±SD) vergence and uIVA aVOR gain increase in all subjects
Mean (±SD) vergence and uIVA aVOR gain increase in all subjects
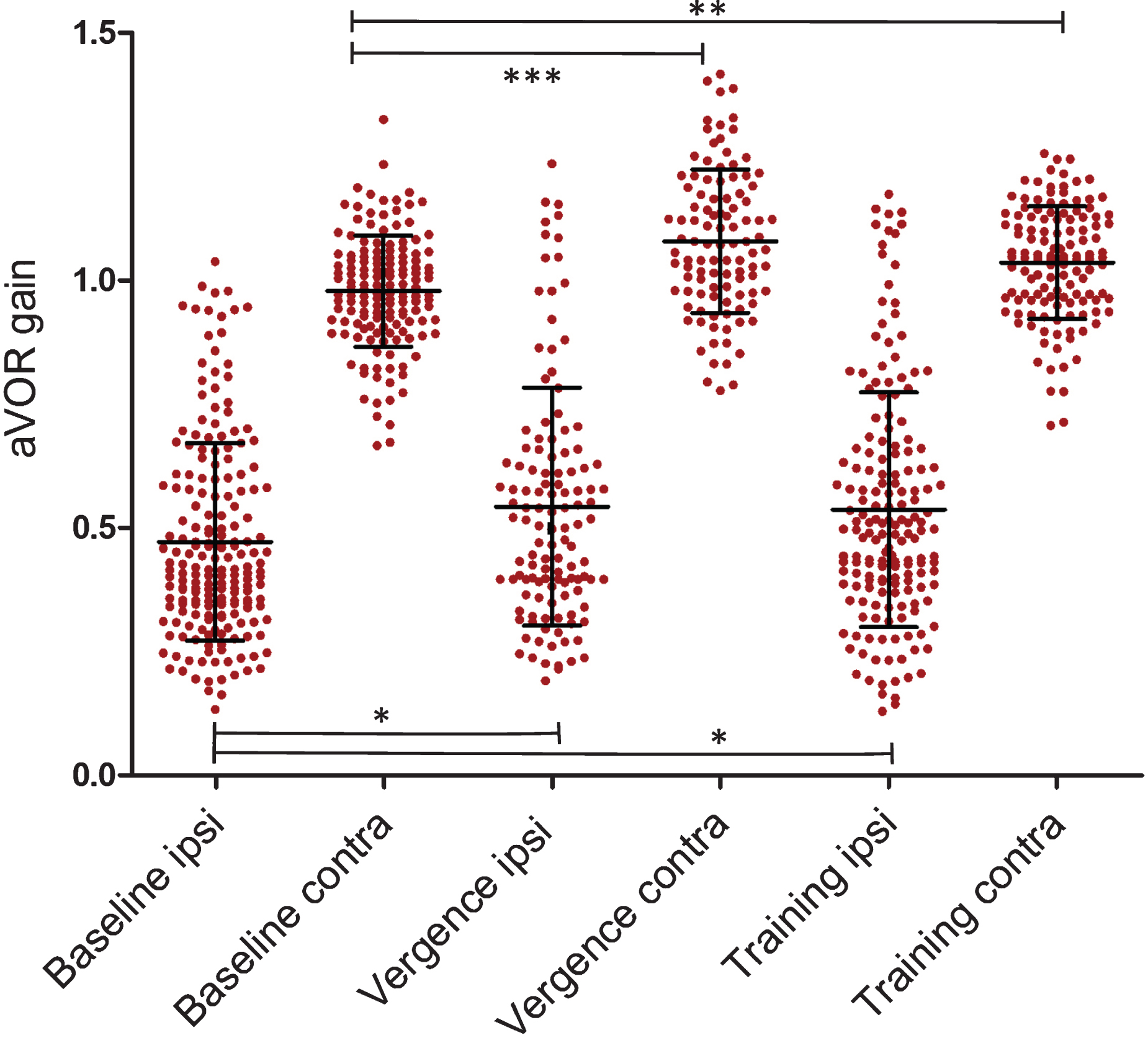
Average aVOR gain values of individual head impulses (±SD) in the different settings in all patients (20 patients). Significant differences are shown between the data groups collected on the same side, *= p < 0.05; **= p < 0.01; ***= p < 0.001. Numbers of individual head impulses (filled circles) are listed in the text.
Patients were divided in two groups based on their response to uIVA: those with ≥5 % gain increase (10/20 patients, ‘adapting’ group), and those with <5 % (10/20, ‘non-adapting’ group). There was no difference in the time of acute vestibular loss between ‘adapting’ (17.400±17.582 months; range 6 to 60 months) and ‘non-adapting’ (21.400±12.258; range 7 to 46) groups (F1,19 = 0.348, p = 0.562). The ipsi- lesional baseline (far viewing) gain values were significantly different between the two groups with ‘adapting’ group gain 0.534±0.212 versus ‘non-ada-pting’ group gain 0.416±0.181 (F1,192 = 17.47, p <0.001). In the adapting group, there were significant ipsi-lesional changes during vergence from the baseline value of 0.512±0.071 to 0.632±0.071 (t-test, HSD = 5.53, p < 0.01); and after adaptation training an increase to 0.64±0.071 (t test, HSD = 6.66, p < 0.01), but no changes on the contra-lesional side (Fig. 3). In the non-adapting group, there were significant contra-lesional changes during vergence from the baseline value of 0.938±0.032 to 1.132±0.033 (t test, HSD = 9.41, p < 0.001), and after adap-tation training an increase to 1.039±0.033 (t test, HSD = 5.28), but no changes to the ipsi-lesional side.
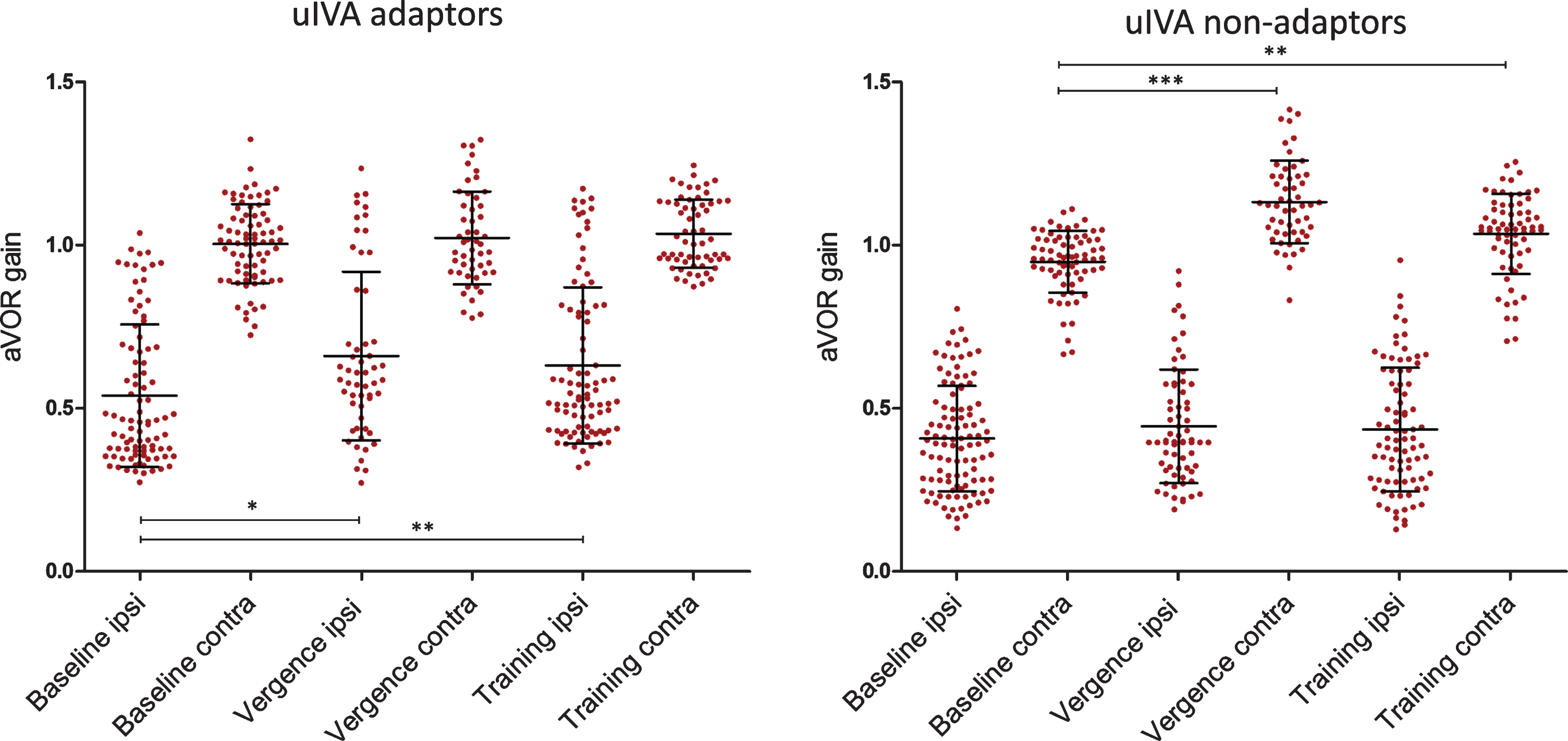
Average aVOR gain values of individual head impulses (±SD) in two separate patient groups. Left panel (A): in this group after training the ipsilateral aVOR gain increased (10 patients ‘adapted’ successfully). Right panel (B): in this group the patients could not increase the aVOR gain due to training (10 patients did not ‘adapt’).Numbers of individual head impulses (filled circles) are listed in the text.
Although 10 patients had≥5 % ipsi-lesional gain increase as a result of adaptation training and 11 patients had≥5 % ipsi-lesional vergence-mediated gain increase, the two groups only partially over-lapped. Chi-square analysis suggests no significant difference in the ratio of those with vergence-med-iated gain increase between adapting (70%) and non-adapting (40%) groups (p = 0.178; Table 2).
Chi-square analysis of the test results: p = 0.178 (‘adapting’ group: patients, who could increase gain due to training)
Correlation analysis revealed poor correlation between gain increases due to uIVA training and near-viewing (Pearson Correlation=–0.262, slope co-efficient p = 0.464, R = 0.262) (Fig. 4).
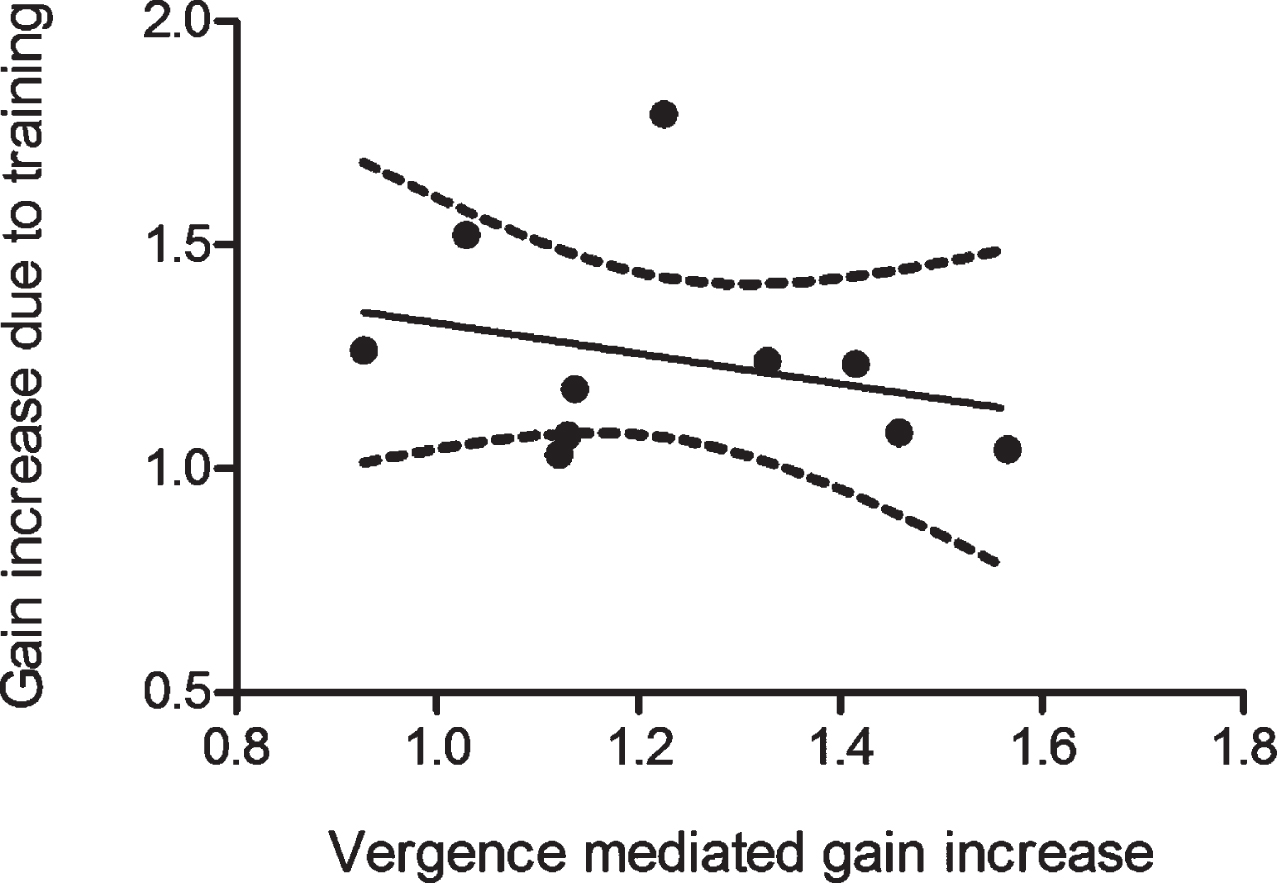
There is a low correlation of the average relative gain increases due to near-viewing and uIVA training during ipsi-lesional impulses in the n = 10 patients showing at least 5% aVOR gain increase. 95 % confidence interval of the best fitting line is shown.
Our data show a poor correlation between the vergence-mediated aVOR gain increase and the gain increase as a result of adaptation training, suggesting that the two rely on different mechanisms and or have unique origins.
Half of our patients with UVH had successful aVOR adaptation after passive uIVA training, and 70% of those had increased aVOR gain during vergence. For the other half of patients, only 40% had increased aVOR gain during vergence. However, this difference (40% vs 70%) was not significant. A limitation of this sub-analysis of the data was the small sample size, i.e., only n = 10 subjects per group; perhaps a larger sample size would have revealed a significant difference.
There is strong evidence that the vergence-med-iated gain increase depends on gating of the irregular (phasic) afferent signal modulated by eye position [17, 18]. In contrast, Ushio et al. [17] speculated that regular (tonic) afferents play the dominant role during gain adaptation training towards the ipsi-lesional side. In monkeys with unilateral labyrinthectomy exposed to repeated transient (high acceleration) rotations towards the ipsi-lesional side while wearing ma-gnifying lenses, the gain significantly increased [15]. The authors suggested this gain increase was mediated by the regular afferent signal coming from the contra-lesional side because the ipsi-lesional afferent signal was abolished due to the labyrinthectomy, and the irregular afferent signal from the contra-lesional side was reduced due to inhibitory cut-off. Our present findings in humans with unilateral peripheral lesions support this hypothesis. The main difference between the primate and present study is that in our subjects there was some remaining vestibular function for ipsilesional rotations, i.e., the gain was reduced by ∼50% but not abolished. Taken together, the present data can be interpreted in one of two ways. One interpretation is that unlike the vergence-mediated gain increase, which depends on the irregular (phasic) signal, regular (tonic) afferents play the more important role in adaptation training. Another interpretation is that vestibular neuritis preferentially affects the irregular afferents that project to the vestibulocerebellum, which have been shown to be important for VOR adaptation via visual-vestibular training [12]. Primary vestibular regular afferents have smaller diameters than irregular afferents and run closer to the outer side of the vestibular nerve (for review see [8]). If vestibular neuritis particularly affected larger diameter more centrally located nerve fibers, then a preferential effect is possible [11]. One way to test this second explanation would be to repeat the study in patients with hypofunction due to canal plugging because with that lesion the afferents themselves would not be affected. Another potential approach would be to determine if the vergence gain is affected by adaptation training. If so, then that would argue against the second explanation.
About half of our patients had a significant ver-gence-mediated aVOR gain increase during head im-pulses to their pathological side. A similar number of patients had a significant aVOR gain increase due to ipsi-lesional passive impulse uIVA training. The percentage of participants that did not adapt (∼50%) was higher than prior uIVA studies that used active head rotation training. The passive applied head rotation prevents efferent copy strategies to en-hance the VOR gain [4], thereby making adaptation more difficult to passive motion. Additionally, in our experience, nearly one-third of patients have some di-fficulty learning how to perform the training task - which when coupled with passive head rotation, likely impaired some of the motor learning. Evidence for this include patients having difficulty tracking the target, and instead blinking during the head impulse or moving their eyes in the direction of the passive head rotation. Many of these responses impede uIVA training though these are strategies patients recruit to ensure function in daily life, and avoid the retinal image slip signal [24]. When we considered only ‘adapting’ patients, the gain increase was similar to data gathered in other uIVA adaptation experiments [20, 25].
There was no difference in the time of acute ves-tibular loss between ‘adapting’ and ‘non-adapting’ groups. However, in the ‘non-adapting’ group, there were more patients with profound ipsi-lesional dysfunction with 20% lower overall mean baseline gain compared to the ‘adapting’ group, which might explain why they had more difficulty performing the training protocol, i.e., the 0.1 gain demand increment during training was relatively larger for the ‘non-adapting’ group making tracking the target harder for them. Only in the ‘adapting’ group was there a sig-nificant vergence-mediated gain increase during ipsi-lesional impulses suggesting a remaining contribution of type I receptor cells and their irregularly discharging afferents [17]. Only in the ‘non-adapting’ group were there significant contra-lesional gain changes, probably because patients found it harder to adhere to the protocol during ipsi-lesional impulses due to their hypofunction, but performed the task better for contra-lesional rotations. Therefore, if their starting contra-lesional VOR gains were below unity, which is typical in patients with UVH, then their aVOR gain was brought closer to unity due to the training gain demand of x1 towards that side.
Conclusion
Our data support that some patients with UVH do retain the ability to enhance their aVOR gain through vergence and incremental passive adaptation, but there is little evidence that these are mediated by the same pathways and mechanism.
Conflicts of interest
The authors report no conflict of interest
Footnotes
Acknowledgments
AAM was supported by The Garnett Passe and Rodney Williams Memorial Foundation Senior Principal Research Fellowship in Otorhinolaryngology
MCS is supported by Department of Defense Awards W81XWH-15-1-0442 and W8lXWH-l7-CTRR-CTA
BB is supported by the grants RTO005 and SF06 of the Karl Landsteiner Private University of Health Sciences