
Abstract
BACKGROUND:
Impairment of visual fixation suppression (VS) in progressive supranuclear palsy (PSP) is not well documented.
OBJECTIVE:
To evaluate the usefulness of impaired VS of caloric nystagmus as an index for differential diagnosis between PSP and Parkinson’s disease (PD), which is often difficult, especially in the early stage.
METHODS:
Subjects comprised 26 PSP patients and 26 PD patients clinically diagnosed at Tokyo Metropolitan Neurological Hospital. We retrospectively investigated VS of caloric nystagmus, horizontal pursuit, saccades, and horizontal optokinetic nystagmus recorded on direct-current-electronystagmography, and neuroradiological findings.
RESULTS:
The median of the average VS% was 0% and 50.0% in PSP and PD patients, respectively. In PSP, VS was impaired even in the early stage of disease. We found a significant correlation between VS and velocity of saccades or maximum slow phase velocity of optokinetic nystagmus only in PSP patients. PSP patients with atrophy of the subthalamic nucleus or with decreased blood flow in the frontal lobe showed significantly more severe impairment of VS.
CONCLUSIONS:
VS may be a useful biomarker to differentiate patients with PSP from those with PD. Cerebellar networks that connect with the cerebral cortex and basal ganglia may contribute to impaired VS of caloric nystagmus in PSP.
Keywords
Introduction
Progressive supranuclear palsy (PSP) is a degenerative central nervous system disease characterized by muscle tone abnormality, postural instability, falling, pseudobulbar palsy, frontal dementia, and supranuclear ophthalmoplegia. PSP was precisely reported by Richardson, Steele, and Olsweski in 1964, and it is thus called classical Richardson-Steele syndrome (PSP-Richardson syndrome; PSP-RS) [71]. The definite diagnosis of PSP can be made at autopsy when observations reveal deposition of abnormal tau protein and neuronal loss involving the basal ganglia, superior colliculus, pons, and dentate nucleus in the cerebellum.
Diagnostic criteria for PSP include the international clinical research criteria that were proposed by the National Institute of Neurological Disorders and Stroke and PSP (NINDS-SPSP) in 1996 [51]. In NINDS-PSP criteria [51], cerebellar ataxia among the early symptoms was an exclusion criterion for clinical diagnosis of PSP, but recent reports suggest that in addition to PSP-RS, various non-classical subtypes of PSP exist, including PSP-parkinsonism [83], PSP-pure akinesia with gait freezing [12, 82], PSP-corticobasal syndrome [13, 80], PSP-frontotemporal dementia [6, 38], PSP-progressive non-fluent aphasia [8, 54], and PSP-cerebellar ataxia (PSP-C) [32, 39]. Then new diagnostic criteria for PSP was presented by the International Parkinson and Movement Disorder Society (MDS)-endorsed PSP Study Group in 2017 (MDS-PSP criteria) [29].
A wide spectrum of PSP subtypes has been described, and the clinical diagnostic criteria for PSP-C are still under discussion [29]. Pathological studies show that the distribution of abnormal tau proteins and neurodegenerative lesions in these subtypes differ from those of classical PSP, and lesions may be distributed predominantly in the cortex or the brainstem [17].
Detailed abnormal eye movements have been reported in PSP-RS. In the early stage of disease, patients usually show impairments in vertical eye movement. As the first sign of the disease, slowing of vertical saccades with a full range of vertical movement is seen, followed by impairments of optokinetic nystagmus (OKN) and vertical gaze palsy. In the advanced stage of disease, horizontal eye movements are also affected. On the other hand, the rotational vestibulo-ocular reflex (VOR) is preserved with tonic deviation of the eyes in the orbit in the direction of slow phases of nystagmus. Finally, all eye movements are impaired [48, 79]. On the other hand, patients with some subtypes of non-classic PSP do not show vertical ophthalmoplegia in the early stage of disease [62]. Some PSP patients occasionally show abnormal eye movement in the late stage of disease or not at all [15, 66]. Although some studies have reported the failure of VOR suppression in PSP [19, 78], the pathogenesis of impaired visual fixation suppression (VS) of caloric nystagmus in PSP has rarely been considered.
Depending on the type of PSP, the degree of disease progression varies, and a range of clinical presentations and eye movement disorders has been reported, making clinical diagnosis of PSP difficult, particularly in the early stages of the disease [50, 57].
With the development of new imaging technology, atrophy of the subthalamic nucleus (STN) with magnetic resonance imaging (MRI) and decreased blood flow in the frontal lobe with single photon emission computed tomography (SPECT) are seen in the early stage of disease, in addition to atrophy of the superior colliculi in PSP. These neuroradiological findings may be useful as biomarkers of PSP for the initial diagnosis.
In the present study, we investigated the impairment of VS of caloric nystagmus in PSP, and evaluated its usefulness as an index for differential diagnosis between early-stage PSP before the limitation of horizontal eye movement occurs and idiopathic Parkinson’s disease (PD). We further considered the pathogenesis of the impairment of VS.
Materials and methods
Patients and controls (Table 1)
Subjects comprised 26 patients with PSP (PSP-RS 11, PSP-PAGF 6, PSP-P 5, PSP-C 4) diagnosed with clinically probable PSP, and 26 patients with idiopathic PD (11 males, 15 females) diagnosed with clinically established or probable PD according to the Movement Disorder Society Clinical Diagnostic Criteria for Parkinson’s disease (MDS criteria) [61], in the Department of Neurology at Tokyo Metropolitan Neurological Hospital.
Characteristics in patients with PSP and PD, and normal controls
Characteristics in patients with PSP and PD, and normal controls
Mean years ± standard deviation. Fisher’s exact test, p value between *1; PSP vs. PD, *2; PSP vs. controls.
The control group, which was age-matched with the PSP group, included 26 patients (14 males, 12 females), diagnosed with spondylosis or orthostatic hypotension after neurological, neuro-otological, and neuroimaging examinations in our hospital.
PSP patients with limited horizontal eye movement and those with prior intracranial lesions such as cerebrovascular disease were excluded from the study.
We retrospectively investigated age, the duration of illness, and neuro-otological findings: gaze nystagmus, square wave jerks, horizontal smooth pursuit, visually guided saccades (saccades), horizontal OKN, cold caloric responses, and the VS test recorded on direct-current electronystagmography (DC-ENG) in patients with PSP and PD.
We also retrospectively evaluated neuroradiological findings such as brain MRI including neuromelanin imaging, and SPECT in patients with PSP and PD.
ENG recordings and analysis
All subjects were observed in terms of primary eye position, range of eye movement, and gaze nystagmus. Abnormal eye movements such as gaze palsy, spontaneous nystagmus, smooth pursuit eye movement (SPEM), visually guided reactive saccades (saccades), OKN, caloric nystagmus, and VS of caloric nystagmus were recorded by DC-ENG with a computerized analysis system (NY-50S®, RION, Tokyo, Japan).
Subjects were seated in a comfortable height-adjustable chair with their heads stabilized on a chin rest in front of the center of a hemicylindrical (Jung type) screen with a radius of 160 cm. Gaze palsy, spontaneous nystagmus, SPEM, saccades, and OKN were recorded. Direct-current recordings were performed in patients to monitor the eye position using Ag-AgCl electrodes. A visual stimulator projection system (CR-58®, NAGASHIMA Medical Instruments Co. Ltd., Tokyo, Japan) was used for the visual target. Time constants were 3.0 seconds for eye movement recordings and 0.03 seconds for eye velocity recordings.
In the gaze nystagmus test, eye movements were recorded when gazing at a visual target (red light emitting diode; LEDs) projected on a screen directly ahead and spaced 0° and 30°, right and left, and up and down. For SPEM, an LED spot was projected and moved sinusoidally in the horizontal direction with an amplitude±30° and frequency of 0.3 Hz. For horizontal saccades, a spot projected and moved in the horizontal direction with an amplitude±30° and frequency of 0.3 Hz was presented. The accuracy of saccades was evaluated as hypometria, hypermetria, or normal, and the maximum quick-phase velocity (MQPV) of saccades was quantified. For horizontal OKN, white stripes as visual stimuli moving to the left or to the right at a fixed velocity of 10°/s, up to at most 60°/s was presented by means of the projector system used by Jung. All subjects were exposed to increasing OKN velocities until OKN deteriorated at higher velocities. The maximum slow-phase velocity (MSPV) of induced left and right optokinetic nystagmus was measured, and the mean of the left and right values was calculated.
Visual fixation suppression in caloric testing
Subjects lay on a bed with their heads raised 30° in an optically and acoustically shielded room. Caloric testing was performed by irrigation of 2 ml ice water for 20 seconds in each ear at a 10-minute interval. During the caloric test, the position of the eyes was continually observed on the monitor of the DC-ENG system. When the eye position showed a tendency toward tonic deviation from the center toward the outside, the subject was instructed to correct the eye position as far as possible to the center. When the slow phase velocity of caloric nystagmus reached a maximum in a dark room, the lights were turned on with the eyes open, and the subject was instructed to gaze at the examiner’s fingertip. The examiner’s index finger was held 50 cm above the subjects’ face as the point on which they were to fix their eyes. The original waveform and the velocity waveform were recorded and analyzed by the DC-ENG system.
Data analysis
VS of caloric-induced nystagmus was determined by measuring mean slow phase velocity during the last 10 s in darkness (a°/s) and comparing it to slow-phase velocity in light with eyes open (b°/s) (Takemori and Cohen 1974 [74]).
VS%, MQPV of Saccades, and MSPV of OKN were evaluated on both sides separately and totally analyzed. Stimulated ears with maximum induced nystagmus slow phase velocity of 10°/s or less during the caloric test were excluded from the VS value study.
Statistical analysis
The number and percentage of gaze nystagmus, square wave jerks, and each radiological finding were represented and analyzed with Fisher’s exact test between groups. The duration of illness was analyzed with the unpaired t-test. VS%, MQPV of Saccades, and MSPV of OKN were investigated statistically with the Mann-Whitney U test. Furthermore, the relationship between VS and Saccades, and VS and OKN were assessed statistically with Spearman’s rank correlation. The significance level was defined as p < 0.05. In statistical analysis, we performed statistical processing without supplementing missing values or excluding external values or outliers. Statistical analysis was carried out using the statistical software R version 3.2.4.
Results
Characteristics of the patients with PSP and PD, and controls
Table 1 shows the profile of patients. The PSP patients and control group were age matched, and hence, patients with PSP and controls were significantly older than those with PD. No significant difference was present between the average duration of illness in the patients with PSP and PD (Table 1).
Gaze nystagmus, square wave jerks, and horizontal saccadic pursuit
Gaze nystagmus, square wave jerks, and horizontal saccadic smooth pursuit were significantly observed in the patients with PSP compared to those with PD. Otherwise, no significant difference was present in horizontal hypometric saccades between the patients with PSP and those with PD (Table 2).
Neuro-otological and neuroradiological imaging findings in patients with PSP and PD
Neuro-otological and neuroradiological imaging findings in patients with PSP and PD
Number of patients/total patients (%). Fisher’s exact test, *p < 0.05, N.S.; not significant, MRI; magnetic resonance imaging, SPECT; single photon emission computed tomography.
We also evaluated patients without cerebellar involvement such as atrophy and/or decreased blood flow in the cerebellum among those with PSP and PD. The frequency of gaze nystagmus and square wave jerks was significantly higher in 6 of 19 and 7 of 19 PSP patients and in 1 of 21 and 0 of 21 patients with PD, respectively.
Table 2 also shows the radiological imaging findings in patients with PSP and PD. Atrophy of the midbrain and STN, and decreased blood flow in the frontal lobe were significantly observed in the patients with PSP compared to those with PD (Table 2).
VS of the caloric test in the patients with PSP and PD, and normal controls
Figure 1A shows a box-plot comparing VS% in the patients with PSP and PD, and normal controls. In PSP patients, VS% was significantly lower than in PD patients and normal controls (Fig. 1A).

A: Box-plot comparing VS in all patients with PSP and PD, and the control group. The median of the average VS for right- and left-induced nystagmus was 0%, 50.0%, and 60.0%, the interquartile range was 31.0%, 10.5%, and 9.5%, the minimum was –48.0%, 30.0%, and 51.5%, and the maximum was 55.0%, 72.5%, and 81.5% in patients with PSP (n = 26) and PD (n = 26), and the control group (n = 26), respectively. In PSP patients, VS% was significantly lower than in PD patients as well as in the control group (Mann-Whitney U test, **p<0.001). B: Box-plot comparing VS in the PSP and PD groups without cerebellar involvement. The median of the average VS for right- and left-induced nystagmus was 0% and 50.0%, the interquartile range was 31.0% and 12.3%, the minimum was –48.0% and 30.0%, and the maximum was 55.0% and 72.5% in the patients with PSP (n = 19) and PD (n = 21), respectively. Similarly, VS% was significantly lower in PSP patients than in PD patients (Mann-Whitney U test, **p < 0.001).
We also evaluated 19 PSP and 21 PD patients without cerebellar involvement as seen with radiological evaluation. Similarly, VS% was significantly lower in PSP patients than in PD patients (Fig. 1B).
Representative cases
Figure 2 demonstrates representative DC-ENG recordings of VS of cold caloric nystagmus following irrigation in the right ear (left) and MRI (right) in a patient with PSP (Fig. 2A) and PD (Fig. 2B). In the PSP patient, reduced VS% of caloric testing and atrophy of the midbrain and STN were seen (Fig. 2A). On the other hand, in the patient with PD, VS% was within the normal range, and the midbrain and STN were preserved (Fig. 2B).
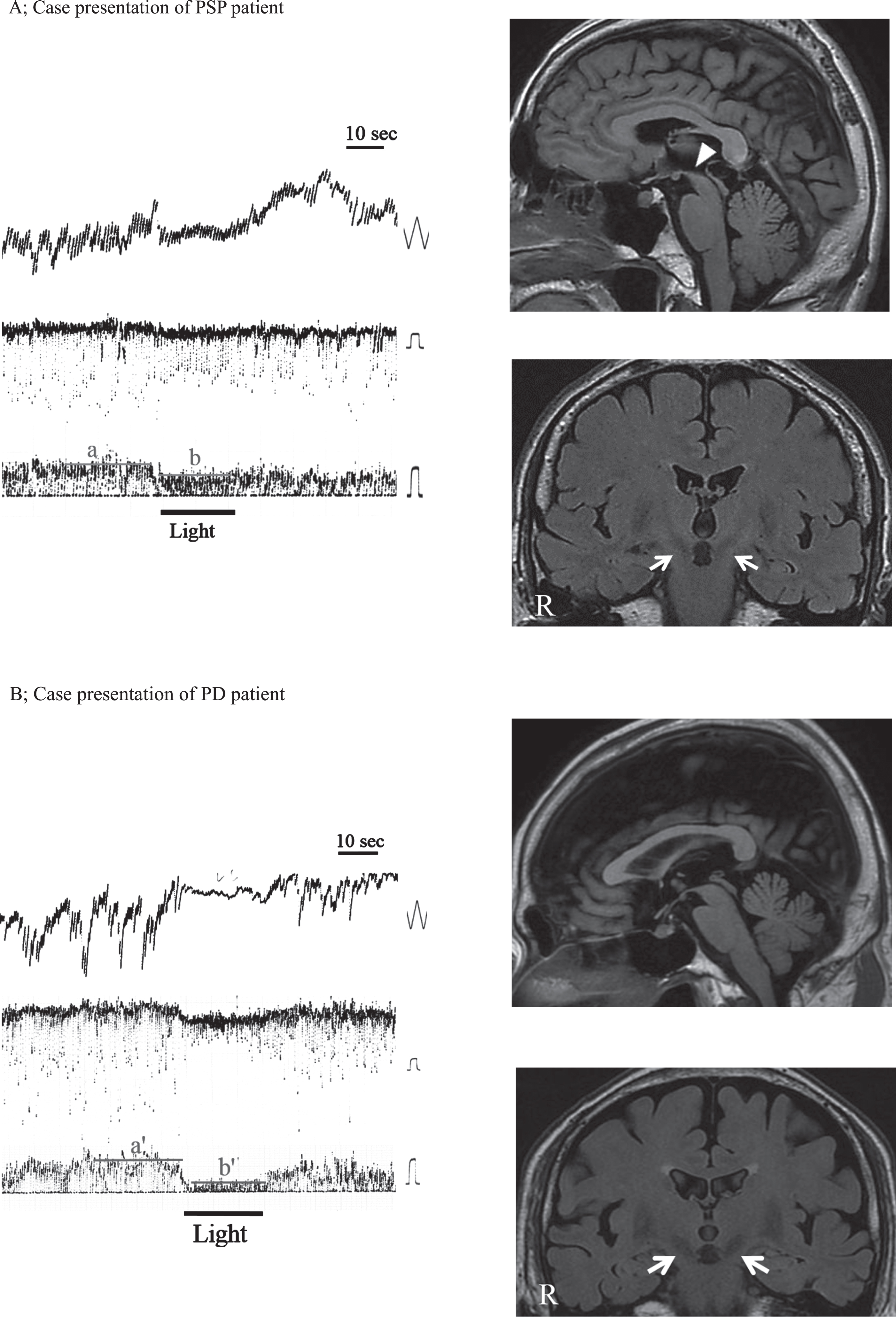
Representative VS recordings and brain MRI in PSP and PD patients. Left: DC-ENG recordings in darkness; Upper: eye-movement recordings, time constant 3.0 seconds, calibration 10 degrees; Middle: the velocity of eye movement, time constant 0.03 seconds, calibration 20 degrees/second; Bottom: slow phase velocity of eye movement, calibration 20 degrees/second, bar: fixation suppression in light. Right: brain MRI; Upper: sagittal plane, Bottom: frontal plane. A: PSP patient, 70-year-old male. a: mean slow phase velocity of caloric nystagmus in darkness, b; mean slow phase velocity of caloric nystagmus in light with eyes fixed on a target. He showed decreased VS% in light. VS% = a –b/a = 20%. MRI revealed atrophy of the midbrain (arrowhead) and reduced signal intensity in the STN (arrows) as well as in the SNc and LC. B: PD patient, 73-year-old male. a’: mean slow phase velocity of caloric nystagmus in darkness, b’; mean slow phase velocity of caloric nystagmus in light with eyes fixed on a target. VS% was within normal range (>40%). VS% = a’ –b’/a’ = 75%. The midbrain and STN (arrows) were preserved.
We investigated the relationship between VS and the duration of illness. We found a weak relationship between VS% and the duration of illness in patients with PD. On the other hand, in PSP, even in the early stage of disease, VS was impaired. The longer the duration of illness, the lower the VS%. The induced nystagmus was even more augmented when the light was turned on in the room. We found a significantly strong relationship between VS% and the duration of illness in PSP patients (Fig. 3).
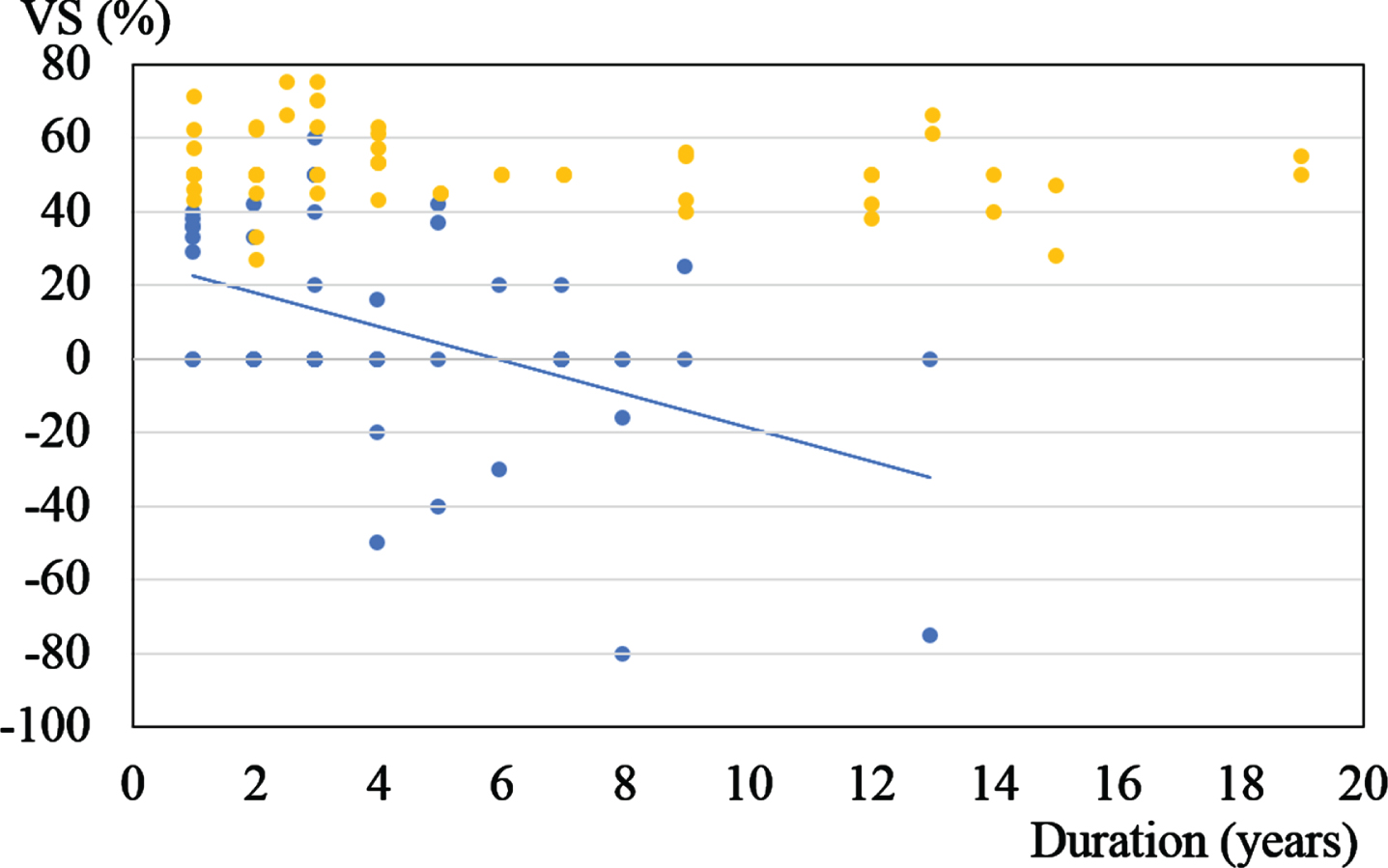
Relationship between VS and the duration of illness. Blue dots: PSP (n = 51 ears, Spearman’s rank correlation, ρ= –0.4, p = 0.03). Yellow dots: PD (n = 52 ears, Spearman’s rank correlation, ρ= –0.2, p = 0.3). In PSP, even in the early stage of disease, VS was impaired. We found a significantly strong relationship between VS% and the duration of illness in PSP patients.
We also evaluated the relationship between VS% and atrophy of the STN in 50 patients with PSP and PD who underwent neuromelanin imaging MRI. We found a significant difference in the VS% between the patients with and without atrophy of the STN (Fig. 4).
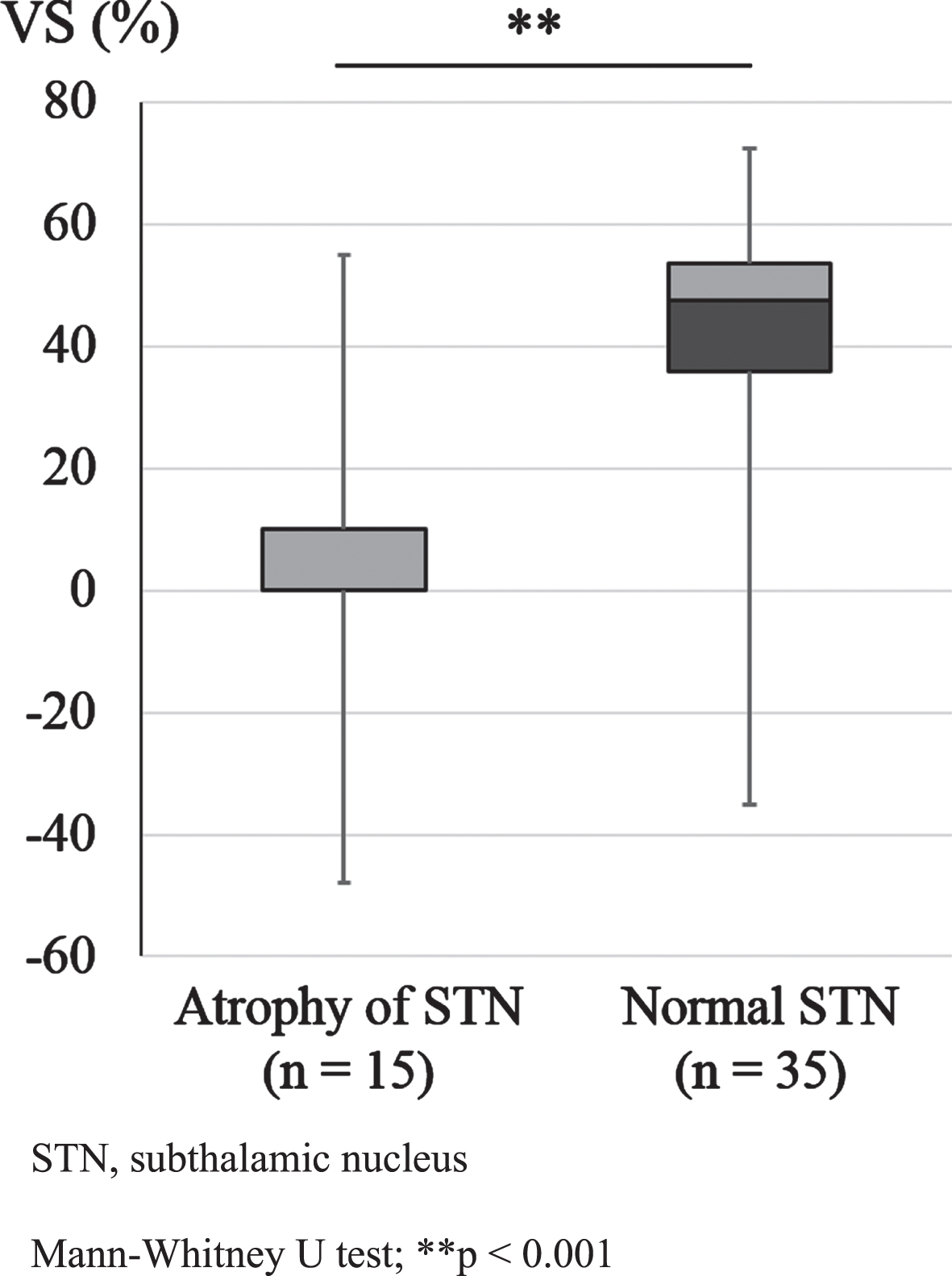
Relationship between VS and atrophy of the STN in all subjects. STN, subthalamic nucleus. The median of the average VS for atrophy of the STN (n = 15) and the normal STN group (n = 35) was 0% and 47.5%, the interquartile range was 10.0% and 17.5%, the minimum was –48.0% and –35.0%, and the maximum was 55.0% and 72.5%, respectively, in the patients who underwent neuromelanin MRI (n = 50). We found a significant difference in VS% between cases with and without atrophy of the STN for all subjects (Mann-Whitney U test; ** p < 0.001).
We investigated the relationship between VS% and blood flow in the frontal lobe in all 44 patients with PSP and PD who underwent both MRI and SPECT. Figure 5 shows the box-plot for VS% of patients with (31 patients) and without (13 patients) findings of decreased blood flow in the frontal lobe. We found significant differences in VS% between patients with decreased and normal blood flow in the frontal lobe.
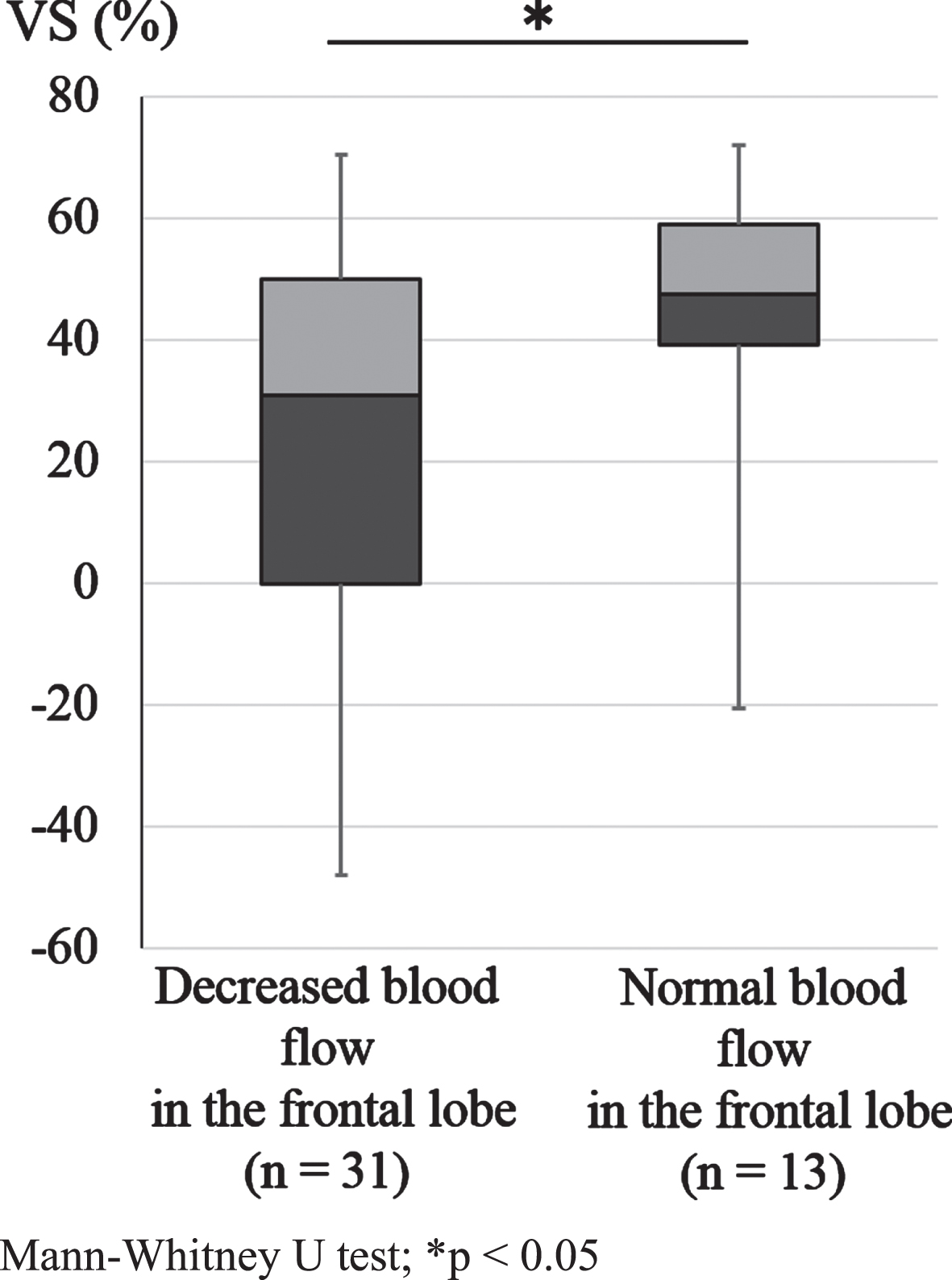
Relationship between VS and blood flow in the frontal lobe in all subjects. The median of the average VS for decreased (n = 31) and normal blood flow in the frontal lobe (n = 13) was 31.0% and 48.0%, the interquartile range was 50.0% and 19.8%, the minimum was –48.0% and –20.0%, and the maximum was 70.5% and 72.5%, respectively, in the patients who underwent SPECT (n = 44). We found a significant difference in VS% between decreased and normal blood flow in the frontal lobe (Mann-Whitney U test, *p < 0.05).
We compared the MQPV of horizontal saccades in both groups. The median of the mean MQPV was 281.0°/s in 26 PSP patients. The median of the mean MQPV was 301.3°/s in 26 patients with PD. We found significant slowing of saccades in PSP patients compared with patients with PD (Mann-Whitney U test, p = 0.005).
Moreover, we investigated the relationship between the MQPV of horizontal saccades and VS. We found a significant correlation between VS and MQPV of horizontal saccades in the patients with PSP. On the other hand, no significant correlation was present between VS% and the maximum velocity of saccades in the patients with PD (Fig. 6).
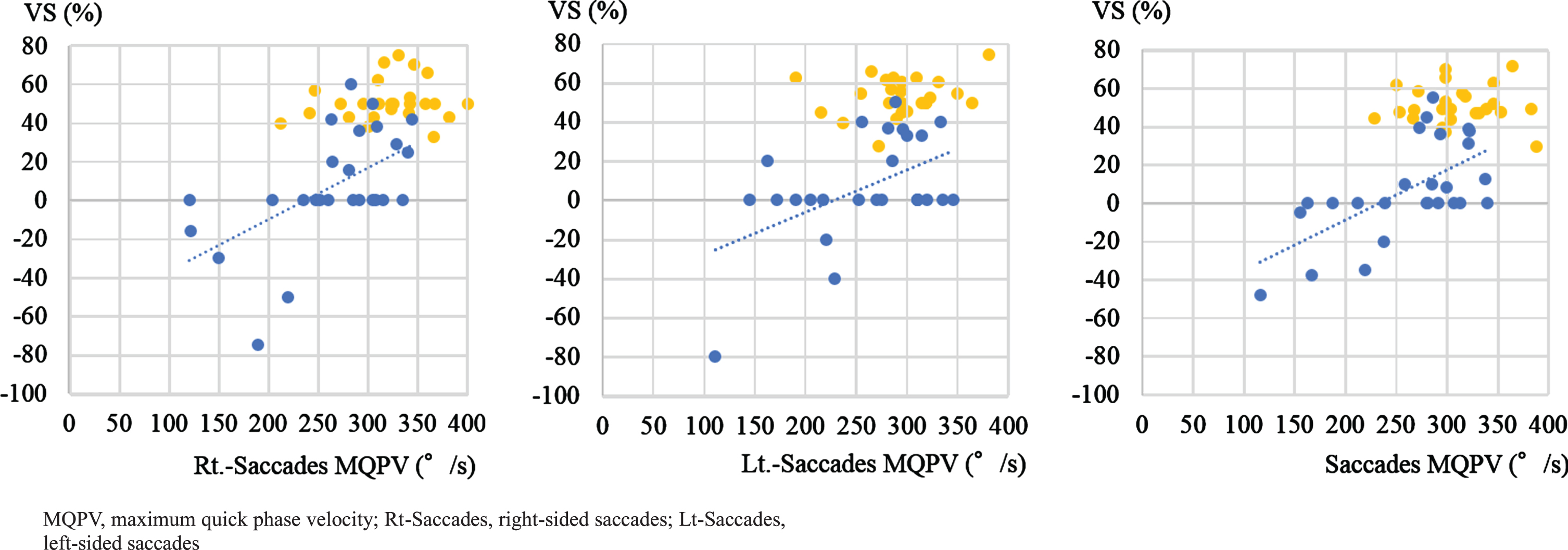
Correlations between VS and the maximum velocity of horizontal saccades. Blue dots: PSP (left: Rt.-saccades MQPV, n = 25, Spearman’s rank correlation, ρ= 0.6, p = 0.003; middle: Lt.-saccades MQPV, n = 26, Spearman’s rank correlation, ρ= 0.3, p = 0.1; right: mean saccade MQPV, n = 26, Spearman’s rank correlation, ρ= 0.6, p = 0.002). Yellow dots: PD (left: Rt.-saccades MQPV, n = 26, Spearman’s rank correlation, ρ= 0.2, p = 0.4; middle: Lt.-saccades MQPV, n = 26, Spearman’s rank correlation, ρ= –0.004, p = 1.0; right: mean saccade MQPV, n = 26, Spearman’s rank correlation, ρ= 0.1, p = 0.8). In PSP patients only, we found a significant correlation between VS% and MQPV of saccades.
We investigated the MSPV of horizontal OKN in both groups. The median of the mean MSPV was 27.0°/s in 26 PSP patients. The median of the mean MSPV was 43.5°/s in 26 patients with PD. In PSP patients, the MSPV was significantly lower than in patients with PD (Mann-Whitney U test, p < 0.001). These data indicate that the involvement of the lower brainstem is more severe in patients with PSP than with PD.
The correlation between VS% and the MSPV of OKN was also evaluated. In PSP patients, we found a significant correlation between VS% and the MSPV of OKN. Hence, patients with PD did not show any significant correlation between VS% and the MSPV of OKN (Fig. 7).
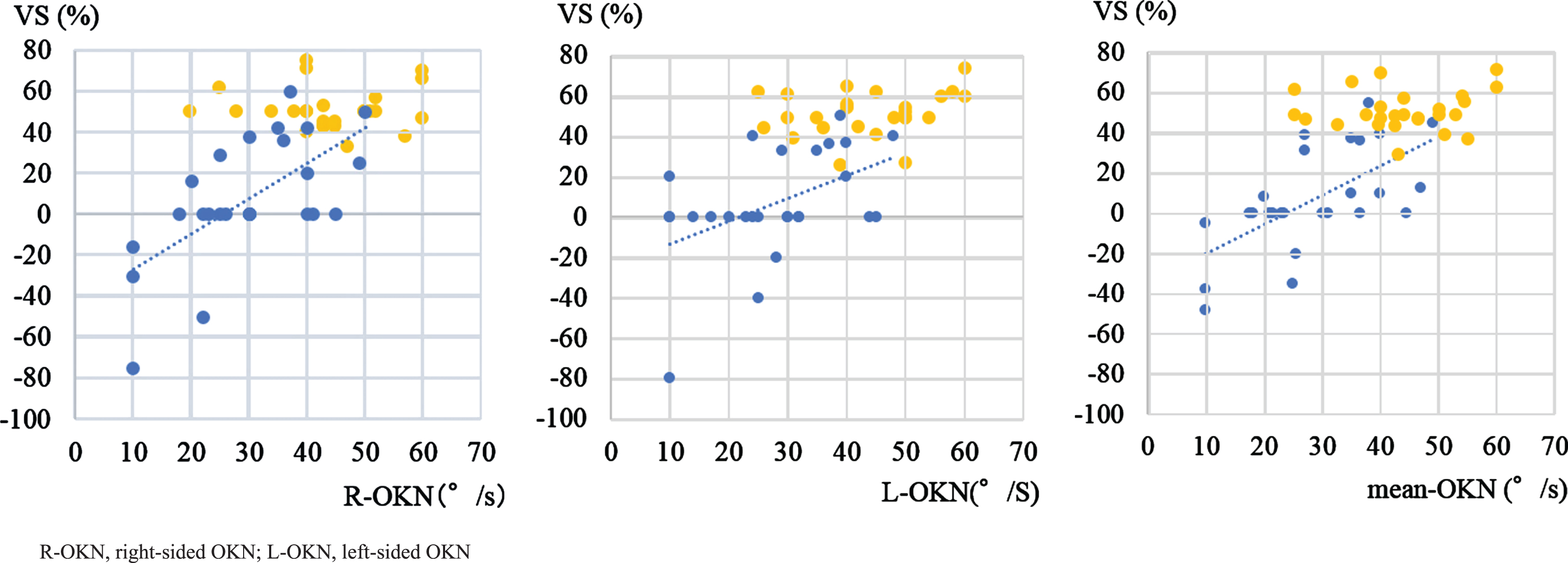
Correlations between VS and the maximum slow phase velocity of OKN. Blue dots: PSP (left: R-OKN MSPV, n = 25, Spearman’s rank correlation, ρ= 0.6, p = 0.001; middle: L-OKN MSPV, n = 26, Spearman’s rank correlation, ρ= 0.4, p = 0.02; right: mean-OKN MSPV, n = 26, Spearman’s rank correlation, ρ= 0.7, p < 0.001). Yellow dots: PD (left: R-OKN MSPV, n = 25, Spearman’s rank correlation, ρ= –0.1, p = 0.7; middle: L-OKN MSPV, n = 26, Spearman’s rank correlation, ρ= 0.3, p = 0.2; right: mean-OKN MSPV, n = 26, Spearman’s rank correlation, ρ= 0.1, p = 0.6). We found a significant correlation between VS% and MSPV of OKN in the PSP group.
Impairment of fixation suppression of caloric nystagmus
Only a few limited reports have been published about the impairment of VS on the caloric test in patients with PSP. A review noted that impairment of VS on caloric tests is found in the early stage of PSP [78]. Duvoisin (1994) uncovered that failure of VOR suppression is common in PSP but is not found in PD [19]. Okuma et al. (1989) [59] and Nawata et al. (1997) [20] reported the impairment of VS in three of four patients and nine of 10 patients with PSP, respectively. Furukawa et al. (1998) described that although no VS abnormalities were seen in any of seven cases of PD, 11 cases of PSP all showed impairment of VS [78].
In the present study, the results showed significantly lower VS% in the PSP group than the PD group (Fig. 1A), which is consistent with the reports from prior studies. Moreover, examination of the relationship between disease duration and the VS value showed that VS was lower in PSP than PD at the early stage of disease, and that the tendency toward lower values in PSP was exacerbated as disease duration became longer (Fig. 2).
These results suggest that findings of VS impairment in the caloric test may be a useful biomarker for differential diagnosis of PSP and PD, regardless of disease subtype or stage.
The pathogenesis of the impairment of VS in PSP (1): the midbrain?
The midbrain including the superior colliculus should be considered as a site of pathogenesis of the impairment of VS in the caloric test in PSP; this impairment is hardly ever seen in PD. Atrophy of the midbrain tegmentum is a cranial MRI finding characteristic of PSP [13, 58]. In the present study as well, atrophy of the midbrain tegmentum was found in all cases in the PSP group and in none in the PD group, a statistically significant finding that is consistent with prior reports.
The centers for saccadic eye movements and visual fixation are distributed in the superior colliculus. The saccades pathways include pathways formed by projections from the frontal lobe that reach the middle layer of the caudal region of the superior colliculus in the midbrain, either directly or indirectly via the basal ganglia, and ultimately, projections to the brainstem premotor neurons that generate horizontal saccades.
Visual fixation, on the other hand, involves the frontal eye field, the visual fixation neurons of the superior colliculus, and omnipause neurons (OPN) in the nucleus raphe interpositus in the midline region of the pons. Inhibitory projections from the OPN to the horizontal system burst neurons continually inhibit saccades, thus maintaining visual fixation. The centers for saccadic eye movements and visual fixation are distributed in the superior colliculus of the midbrain tegmentum, and this is the site of the system that drives the opposing functions of visual fixation and saccade generation through the mutual inhibition loop of OPN and inhibitory burst neurons [69]. The quick phase pathway of the horizontal VOR system and the horizontal saccades system pathway are similar [73]. Furthermore, some reports indicated that the superior colliculus may contribute to the control of smooth pursuit as well as saccades [46, 52]. Therefore, in the case of visual fixation suppression during caloric nystagmus, the superior colliculus may play a role to suppress caloric nystagmus.
Loss of VS was constantly observed in seven of nine cats after creation of a superior colliculus lesion [42]. On the other hand, nine patients with midbrain lesions showed normal VS [76]. It may thus remain a challenge to explain VS abnormalities in PSP due to PSP-specific midbrain tegmental disorders solely.
The pathogenesis of the impairment of VS in PSP (2): the cerebellum?
Visual fixation suppression in caloric nystagmus and the cerebellum
It has been implicated that cerebellar lesions: the nodulus and flocculus [74, 77], and the y group which receives afferents from the flocculus and paraflocculus Purkinje cells [84], are involved in the impairment of VS of caloric nystagmus.
In the caloric test, cold water is irrigated into the external auditory canal on one side, and the temperature change in the lateral semicircular canal creates a convective current in the endolymph, causing deviation of the cupula [3]. This results in depolarization of sensory hair cells, with increased spike frequency of the vestibular primary nerve and increased activity of vestibular nucleus cells, which project to the horizontal eye movement neurons, causing slow co-ocular deviation to the ipsilateral side (slow phase), followed by quick eye movement to the contralateral side (quick phase). This is the phenomenon of caloric nystagmus (vestibular nystagmus). At the same time, the vestibular primary nerve sends out a side branch to the cerebellar flocculus, and Purkinje cells in the flocculus suppress the vestibular nucleus, thus regulating the gain (output/input) of VOR [31]. The cerebellar flocculus also receives visual information from the retina and plays an important role in controlling eye movement with the result of VOR [49, 64].
Involvement of the cerebellum in PSP
In PSP, the core pathological lesions are in the basal ganglia and brainstem, with the globus pallidus, STN, and substantia nigra being most impaired [16, 81]. Cases with early marked cerebellar ataxia were excluded from the NINDS-SPSP clinical research criteria for PSP [51].
However, the presence of cerebellar disorders in four of nine patients with PSP had been described already in the first report of PSP [72]. PSP-C [4, 39] has been known as one of an atypical PSP subtype that is very rare in Europe, the United States, and Canada [34, 83] and accounting for 14% of cases in Japan [36]. Moreover, cerebellar lesions are described not only in PSP-C but also in other subtypes of PSP patients. Atrophy of the cerebellar dentate nucleus and superior cerebellar peduncle occurs in addition to the globus pallidus, STN, and substantia nigra lesions that are inevitably present in PSP [39]. A study using transcranial magnetic stimulation, which is a noninvasive technique to stimulate the human brain, also suggested cerebellar dysfunction in patients with PSP without clinical cerebellar signs [70].
Based on these factors, cerebellar dysfunction may be present in PSP, although clinical cerebellar ataxia may be rarely detected in patients with PSP.
In the present study, in the PSP group, we identified statistically significant findings of horizontal saccadic pursuit, which is generally seen with dysfunction of the dentate nucleus of the cerebellum flocculus. Gaze nystagmus and square wave jerks, which could be due to dysfunction of the cerebellum [40], were seen more frequently in the PSP group than in the PD group. Those findings suggest the presence of a functional disorder of the cerebellum in a broad spectrum of PSP patients but not in PD patients.
Impairment of VS and cerebellar dysfunction
To exclude the influence of the primary involvement of the cerebellum in the impairment of VS, we observed 19 and 21 patients with PSP and PD, respectively, without cerebellar involvement such as atrophy of the cerebellum and/or decreased blood flow in the cerebellum according to neuroradiological imaging findings. Surprisingly, the results were similar to those in the examination of the entire PSP group. Even when cases with no cerebellar change in the neuroimaging study were examined, the PSP group still had a significantly lower VS% and higher presence of gaze nystagmus, square wave jerks, and saccadic pursuit than the PD group (Fig. 1B). In the PSP group with no apparent cerebellar disorders as seen with imaging, a greater functional disorder of the cerebellum flocculus may be present compared to the PD group. This means that even in cases of PSP without cerebellar involvement neuroradiologically, impairment of VS on the caloric test may be related to a functional disorder of the cerebellum.
VS impairment and the STN in PSP
As Fig. 3 shows, in PSP, VS was impaired, even in the early stage of disease. However, can the cause of VS abnormalities from the early stage of the disease be explained only as direct cerebellar dysfunction, even when radiologically invisible? From the early stage of the disease, degeneration would start with involvement of the STN and substantia nigra in PSP, but no STN disorder is present in PD [26]. Albers reported evidence of oxidative stress in the STN that was invisible in the cerebellum in PSP [1]. Another group reported an autopsy of a case of early-stage PSP in which the degeneration started from the substantia nigra and STN [67].
Neuromelanin images obtained using 3-Tesla MRI high-resolution fluid-attenuated inversion-recovery coronal slices have allowed visualization of melanin-containing neurons in the substantia nigra pars compacta (SNc), the locus coeruleus (LC), and the STN. Reduced signal intensity has been reported in the SNc and LC in both sporadic PD and PSP patients [41, 68], whereas atrophy of the STN, which is one of the most severely affected nuclei in PSP [16, 81], has been reported as a finding specific to PSP. Also in the examination of neuroimaging in the present study, atrophy of the STN was observed more frequently in the patients with PSP than in those with PD (Table 2), a result that is consistent with prior reports.
STN in the cerebral basal ganglia for the eye movement loop
The STN is a nucleus of the cerebral basal ganglia and has anatomical connections with both low-level oculomotor nuclei through nigro-collicular projections and high-level frontal and prefrontal areas [47, 65]. Therefore, the STN is an important hub that regulates inhibitory control related to visuo-oculomotor action selection.
The cerebral basal ganglia control somatic motor output by acting as a behavior selection switch, receiving input from the cerebral cortex and giving out commands to execute the appropriate actions at the appropriate times. During voluntary eye movement, projections from the cerebral cortex to the basal ganglia control three pathways. In the direct pathway, direct inhibitory output is present from the striatum to the substantia nigra pars reticulata (SNr). In the indirect pathway, inhibitory stimuli are sent from the striatum to the STN via the outer segment of the globus pallidus, and excitatory output is then sent from the STN to the SNr [2]. In the hyperdirect pathway, excitatory output is sent from the cerebral cortex to the SNr via direct projections to the STN [56]. The SNr suppresses the superior colliculus, so when the eyes are at rest, the SNr is active. As the superior colliculus drives the brainstem reticular formation saccade generator (the paramedian pontine reticular formation; PPRF), horizontal saccades are thus controlled. The direct, indirect, and hyperdirect pathways form an oculomotor loop that goes through the thalamus to the cerebral cortex. This loop is involved in regulation of eye movement and sends inhibit-disinhibit-inhibit signals that result in temporal and spatial effects on the activity of the thalamus and cerebrum, finally resulting in necessary eye movements with the correct timing and suppression of unnecessary movements. Through the STN, the hyperdirect and indirect pathways suppress movements that are unnecessary to the movements that have been induced, and they fulfill the role of clarifying the start and finish of the necessary movements that have been induced by the direct pathway.
Namely, the STN receives projections from the frontal lobe (hyper-direct pathway) and communication from the basal ganglia (indirect pathway) and has output pathways to regions including the midbrain superior colliculus, the pedunculopontine tegmental nucleus, and the cerebellar nucleus. Saccade-related neurons have been found in the STN [53], indicating involvement of the STN in eye movements.
STN function in PSP and PD
STN activity is quite different between PSP and PD. A decrease in dopamine occurs in PD due to degeneration or loss of dopaminergic neurons in the SNc of the basal ganglia. However, excitatory input to direct pathway neurons and inhibitory input to indirect pathway neurons are impaired, causing an increase in STN activity, which thus increases the activity of the internal segment of the globus pallidus. In PSP, on the other hand, although degeneration or loss of the substantia nigra of the basal ganglia is present in the same way as in PD, unlike PD, extensive neurodegeneration of cortical and subcortical nuclei and a clear disorder of the STN are present [28, 72].
In the present study, VS abnormalities were significantly associated with PSP from the early stage of the disease (Fig. 3), and a relationship was found between STN atrophy and the VS value (Fig. 4) in PSP but not PD. These results therefore suggest the possibility of a relationship between STN atrophy and PSP.
Decreased blood flow in the frontal lobe in PSP
In a positron emission tomography study, decreased blood flow and oxygen consumption in the frontal lobe and decreased cerebral metabolism in the frontal lobe (hypofrontality) and the caudate have been identified in PSP [14, 60]. Further, Goffinet et al. and Blin et al. described decreased cerebral metabolism in the basal ganglia, thalamus, and cerebellum in addition to the frontal lobe [7, 22]. SPECT shows decreased blood flow in the frontal lobe, parietal lobe, and basal ganglia [35], as well as in the caudate [43].
Our results are consistent with the reports of decreased blood flow in the frontal lobe in PSP of these reports. Also, our statistical examination of the relationship between the VS value and imaging findings in the overall PSP and PD groups suggests that the decreased blood flow in the frontal lobe, including the frontal eye field (FEF), was statistically related to the VS value in the caloric test.
Frontal lobe and fixation or pursuit
The frontal lobe FEF has visual fixation cells that continuously fire during fixation and stop firing during saccades [33], but the FEF is also involved in pursuit as well as in saccades. The FEF generates the movement commands that cause saccades and pursuit and transmits them via the brainstem to the brainstem premotor neuron groups involved in both types of eye movements. The FEF directly activates gaze centers. Some of these commands are sent via the pontine nucleus or the nucleus reticularis tegmenti pontis to the cerebellar hemisphere or the cerebellar vermis. Some are sent via the superior colliculus to the vermis and further project to the cerebellar vermis dorsal cortex and the fastigial oculomotor region (FOR) or the PPRF.
Hypometria of horizontal saccades is seen with dysfunction of the FOR of the cerebellar vermis, which controls the amplitude of saccades. In the present study, however, no significant difference in the frequency of hypometria was found between the PSP and PD groups, and no association was found between the VS abnormalities on the caloric test in PSP and cerebellar vermis FOR disorder.
VS and OKN or saccades
In the present study, the MSPV of OKN and the MQPV of horizontal saccades were significantly lower in the PSP group than in the PD group. Further, the VS value in the PSP group showed a relatively high, statistically significant association with decreased MQPV of horizontal saccades and decreased MSPV of OKN (Figs. 6, 7).
The caudate in the striatum is activated by optokinetic simulation [18]. Projections from the cerebral cortex to the striatum (direct and indirect pathways), or to STN (hyper-direct pathway), control saccades via SNr which suppresses the superior colliculus (SC). SC drives the brainstem saccade generator, PPRF (Fig. 8).
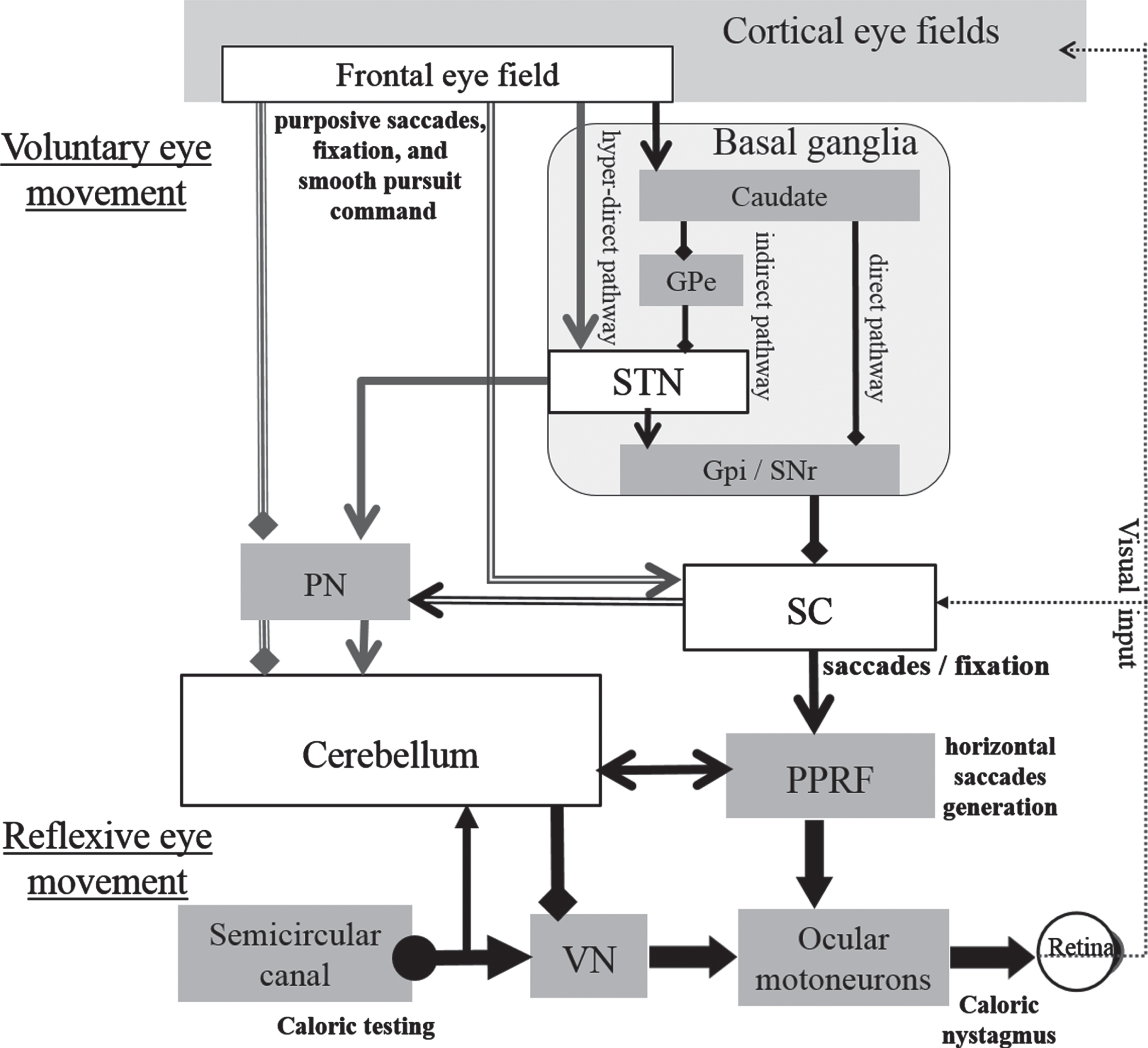
Simplified outline of projections from cortical eye fields to superior colliculus via basal ganglia and to cerebellum as higher-level voluntary control of visual fixation suppression for caloric nystagmus as a result of vestibulo-ocular reflex. From basal ganglia or cerebellum, the projection path leading to the cortical eye fields via the thalamus are omitted. In the caloric test, the temperature change in the semicircular canal creates a convective current in the endolymph with increased spike frequency of the vestibular primary nerve and increased activity of vestibular nucleus cells, which project to horizontal motoneurons, causing caloric nystagmus. At the same time, the vestibular primary nerve sends out a side branch to the cerebellar flocculus, which suppresses the vestibular nucleus. During voluntary eye movement such as fixation, projections from the cerebral cortex to the striatum control three pathways, which form an oculomotor loop that goes through the thalamus to the cerebral cortex. The SNr suppresses the superior colliculus, and the superior colliculus drives the brainstem horizontal saccade generator, PPRF. The centers for saccadic eye movements and visual fixation are distributed in different parts of the superior colliculus. Visual fixation involves the frontal eye field and the visual fixation neurons of the superior colliculus. Inhibitory projections from the OPN to the horizontal system burst neurons continually inhibit saccades, thus maintaining visual fixation. Although degeneration or loss of the substantia nigra of the basal ganglia is seen in PSP as well as in PD, extensive neurodegeneration of cortical and subcortical nuclei and a clear disorder of the STN are seen only in PSP. Axons from the frontal lobe and STN projecting to the cerebellum could play an important role with the superior colliculus during fixation suppression in light for caloric nystagmus induced as a result of VOR. Gpe, Globus pallidus externa; SNc, Substantia nigra pars compacta; Gpi, Globus pallidus interna; STN, Subthalamic nucleus; SNr, Substantia nigra pars reticulata; SC, Superior colliculus; PN, Pontine nuclei; PPRF, Paramedian pontine reticular formation; VN, Vestibular nucleus; VOR, Vestibule-ocular reflex. The pathways concerned with the frontal eye field, STN, superior colliculus, and cerebellum are emphasized. ←; stimulation, ♦—; inhibition.
Those results are consistent with a relationship among the frontal lobe, STN, SC and PPRF dysfunction for the impairment of VS in PSP.
Previous reports have described the evidence that the smooth pursuit system is involved in fixation suppression of vestibular nystagmus [11, 24]. However, we speculate that both the quick and slow phases of ocular movement are relevant to the impairment of VS in PSP.
In humans, following vestibular stimuli, neuroimaging showed high activation in the putamen, cerebellum including the uvula, nodulus, flocculus, and paraflocculus, vestibular nuclei [10], both cerebellar hemispheres [5], as well as cortical areas such as the inferior parietal lobule.
Little has been described about the relationship between VS abnormalities in the caloric test and higher centers. Takemori et al. reported VS abnormalities on caloric testing caused by only inferior parietal lobule lesions in the cerebrum [75]. Naito et al. reported that during visual fixation, in addition to activation of the visual cortex, cerebral activation was seen in the inferior parietal lobule on the side of quick phase nystagmus, the posterior insula, and the cerebellar vermis and hemisphere [55]. Gardner et al. reported that patients with PSP show significant connectivity disruptions within the network of neurons connected to the dorsal midbrain, including the brainstem, cerebellum, diencephalon, basal ganglia, and cerebral cortical areas [21]. The present results support the possible involvement of the effects of the higher center in VOR suppression disorder, which is in line with prior reports.
The existence of a pathway linking the dentate nucleus of the cerebellum, thalamus, and striatum has recently been shown [9, 30], and the STN and cerebellum, dentate nucleus, thalamus, and striatum form a network of neural circuits.
When these results are taken together, impairment of VS abnormalities on the caloric test in PSP clinically indicates not only direct cerebellar malfunction, but also suggests the existence of effects involving the cerebellar networks with basal ganglia such as the STN–pontine nucleus–cerebellar cortex–thalamus–striatum communication, and the cerebral cortex–basal ganglia loop, which is a neural pathway with a major role in brain function for voluntary movement and cognition. To put it differently, the involvement of extensive neural networks of the cerebellum, basal ganglia, and cerebrum is suggested as a higher central mechanism controlling the vestibule-ocular reflex under the tentorium to avoid a perceptual visual-vestibular mismatch (visual-vestibular interaction).
The number of cases in the present study was small, and the statistical study was uneven. Further work in the future with a greater number of cases is needed to investigate midbrain tegmentum and STN atrophy and decreased frontal lobe blood flow as mechanisms for VS abnormalities in PSP, by studying how these relate to VS abnormalities among the various complex neural pathways as the disease progresses.
In conclusion, impairment of VS on the caloric test may be a very useful, cost-effective marker for differential diagnosis between PD and PSP. VS disturbance upon caloric stimulation observed from the early stages of PSP cannot be entirely explained by a direct disturbance of the cerebellum alone, and more extensive disorders may therefore be involved, such as breakdown of the basal ganglia–cerebellum connections and/or the cerebrum–basal ganglia loop caused by atrophy of the STN, decreased frontal lobe blood flow, and/or impairment in the midbrain superior colliculus. We suggest that cerebellar networks with the cerebral cortex and basal ganglia may contribute to impairment of VS of caloric nystagmus in PSP.
Footnotes
Acknowledgments
The authors thank Mrs. Chieko Imai and Mrs. Midori Isa for their excellent technical support with the electronystagmography recording.
Funding
The authors have no financial conflicts of interest relevant to this article to disclose.
Competing interests
The authors declare no competing interests.
Supplementary materials
The subjects’ consent was obtained according to the Declaration of Helsinki, and the study was approved by the ethical committee of Tokyo Metropolitan Neurological Hospital, Tokyo, Japan (TS-H29-006).