
Abstract
BACKGROUND:
Eurycoma longifolia (EL) is a well-known aphrodisiac herb for men. Recently, the crude extract of EL was reported to possess anti-osteoporotic activities.
OBJECTIVE:
This study aims to determine the bone protective effects of the standardized quassinoid-rich EL extract in testosterone-deficient rat model.
METHODS:
Ninety-six intact male Sprague-Dawley rats were randomized into baseline, sham, orchidectomized, and chemically castrated groups. Chemical castration was performed via subcutaneous injection of degarelix at 2 mg/kg. The orchidectomized and degarelix-induced rats were administered with vehicle, intramuscularly injected with testosterone once a week, or orally supplemented with EL extract at doses of 25 mg/kg, 50 mg/kg or 100 mg/kg daily for 10 weeks. Bone mass, microarchitecture and strength were analyzed by dual-energy x-ray absorptiometry (DEXA), micro-CT and three-point bending test.
RESULTS:
Whole body bone mineral density and femoral bone mineral content significantly increased in testosterone groups (p < 0.05). Micro-CT analysis revealed that trabecular bone volume, number, separation and connectivity density were significantly improved by testosterone administration. However, the structural model index was only improved in degarelix group supplemented with 100 mg/kg EL extract (P < 0.05). The improvement of cortical thickness by EL extract was similar to that of testosterone groups (p < 0.05). Biomechanically, EL extract supplementation was able to improve stiffness, strain and modulus of elasticity in degarelix-induced groups, while stress parameter was significantly improved in orchidectomized groups (p < 0.05).
CONCLUSION:
Quassinoid-rich EL extract enables to protect against bone loss due to testosterone deficiency. The protective effect on cortical thickness and biomechanical parameters is comparable to testosterone group.
Introduction
The incidence of osteoporosis in men is often overlooked even though it is recognized as a debilitating condition that cause a significant morbidity and mortality in older men [1]. Although the prevalence of osteoporotic fracture is lower in men than in women, the mortality rate following hip fracture has been reported to be higher in men [2, 3]. Osteoporotic fractures may not only compromise the quality of life of the patients but also cause an increase in global economic and societal burden [4, 5].
Late-onset hypogonadism (LOH) or hypogonadism in older men, hypogonadotropic hypogonadism and patients receiving androgen deprivation therapy (ADT) are among the populations that are prone to comorbid testosterone deficiency syndrome and osteoporosis [6]. Gonadotropin deficiency in congenital or acquired hypogonadotropic hypogonadism leads to low serum testosterone levels and is associated with low bone mass. Men receiving ADT for the treatment of advanced prostate cancer had elevated bone turnover markers and reduced bone mineral density [7]. ADT is also associated with a 2-fold increase incidence of osteoporosis and a 50% increase in fracture incidence [8].
Although not as abrupt as menopause, androgen deficiency in males due to chemical or surgical castration, or an age-related declined in androgens level adversely affected the skeleton [9]. It has long been known that sex steroids are the chief hormones for the regulation of skeletal growth and maintenance in both genders [10, 11]. The loss of pro-apoptotic effects on osteoclast and anti-apoptotic effects on osteoblast and osteocytes in sex-steroids deficiency, is associated with high rate of bone remodeling, resulting in progressive loss of bone mass and strength [9].
Animal model for osteoporosis is crucial for the evaluation of the efficacy and safety of interventions intended to prevent and/or reverse bone fragility at the preclinical level [12]. Orchidectomized rat has been used extensively as a model for male osteoporosis due to testosterone deficiency. Alternatively, bone loss can be induced in intact experimental animals with the use of pharmaceutical agents such as gonadotropin-releasing hormone (GnRH) agonists, estrogen receptor antagonists and aromatase inhibitors [13]. Earlier studies have demonstrated that GnRH agonist and estrogen receptor antagonist can also induce bone loss similar to ovariectomy in female rats [14, 15]. However, studies on the use of these pharmaceutical agents in inducing bone loss in male rats are still lacking. To the best of our knowledge, this is the first study that utilized GnRH antagonist, degarelix as an agent to induce bone loss in intact male rats. Degarelix is a new class of GnRH antagonist that has been used as an ADT for the treatment and management of advanced prostate cancer [16]. In the present study, surgical orchidectomy and chemical castration with degarelix were adopted as models of androgen-deficient osteoporosis for the evaluation of the potential anti-osteoporotic agent.
Despite the significant advances in pharmacological options, conventional therapies for the treatment of osteoporosis remained unsatisfactory due to the issues with patient compliance, adverse effects, cost and long-term efficacy [17]. The antiresorptive therapies including bisphosphonates, teriparatide and hormone replacement therapy are the most commonly prescribed class of drugs but are not free of side effects [17, 18]. Testosterone replacement therapy has been associated with cardiovascular disease, benign prostatic hyperplasia and is not recommended for patient with underlying or high risk factors of cancer [19]. Therefore, the search for effective, low-cost alternative modalities with fewer side effects is highly desirable.
Eurycoma longifolia (EL) or popularly known as ‘Tongkat Ali’ in Malaysia is well-reputed among the natives of South East Asia for its aphrodisiac potential [20]. The bioactive compounds quassinoids, which is unique to this Simaroubaceae family, displayed various medicinal potential including antimalarial, antipyretic, antimicrobial and anticancer activities [21–24]. The root extract of this plant was recently reported to possess anti-osteoporotic activities. Earlier studies in orchidectomized male rats have demonstrated that the crude extract of this plant was able to prevent bone calcium loss [25, 26]. However, the bone protective potential of EL extract was not prominent in bone morphometric studies [27, 28]. These studies have used crude aqueous extract, which contain glycosaponin and eurypeptide in higher percentage than the quassinoids concentration. In light of discovering the true potential of EL, the present study was carried out to evaluate the effects of standardized quassinoid-rich EL extract in both intact and surgically castrated male rats. The standardized quassinoid-rich EL extract contains higher quassinoid concentration of which more than 10 folds of eurycomanone and its analogues were isolated [29]. Hence, we anticipated that this standardized extract might provide better protection against osteoporosis induced by testosterone deficiency in both surgically and chemically castrated male rats.
Material and methods
Animals and treatments
Ninety-six intact-male Sprague-Dawley rats at 3-month-old of age were obtained from the Laboratory Animal Resource Unit, Faculty of Medicine, Universiti Kebangsaan Malaysia (UKM). They were allowed to acclimate to an environmentally controlled room (12-h light/dark cycle, room temperature) in an animal care facility for one week. The rats were housed in plastic cages and were given free access to standard pellet diet and water. Experimental protocols concerning the use of laboratory animals were reviewed and approved by the UKM Animal Ethics Committee (FP/FAR/2015/NAZRUN/25-MAR./665-MAR.-2015-DEC.-2017).
At the beginning of the study, rats weighing between 300-350 g were randomized into four main groups, which were the baseline, sham-operated (SHAM), surgically- and chemically-castrated groups. Surgical and chemical castration was used as the model of androgen-deficient osteoporosis. The baseline group was sacrificed at the beginning of the study without undergoing any surgical treatment or intervention. The rats in the SHAM group underwent similar surgical procedure stress but the testes were not removed. Surgical castration was performed by bilateral orchidectomy through a small incision at the tip of the scrotum. The tunic was opened and the cauda epididymis, vas deferens, spermatic blood vessels and testes were exteriorized and removed. Meanwhile chemical castration was performed by subcutaneous injection of degarelix around the scapular region at the dose of 2 mg/kg twice. Hormonal changes following surgical testes removal and chemical castration by degarelix lead to testosterone deficiency, which results in accelerated bone loss and the development of osteoporosis.
The first injection of degarelix was done at the beginning of the study while the second injection was performed at 6 weeks from the first injection. The surgically castrated groups were subdivided into orchidectomized control (ORX), orchidectomized and given testosterone at 7 mg/kg (ORX + TES), orchidectomized and given standardized EL extracts at 25 mg/kg (ORX + EL25), 50 mg/kg (ORX + EL50) and 100 mg/kg (ORX + EL100). The chemically castrated groups were subdivided into degarelix-induced control (DGX), degarelix-induced and given testosterone at 7 mg/kg (DGX + TES), degarelix-induced and given standardized EL extracts at 25 mg/kg (DGX + EL25), 50 mg/kg (DGX + EL50) and 100 mg/kg (DGX + EL100). The treatment was initiated a week after orchidectomy and degarelix injection. Testosterone enanthate was given intramuscularly once a week while the standardized EL extracts were given daily for 10 weeks via oral gavages.
At the end of the treatment period, the rats were euthanized with ether and blood was collected from the orbital sinus. The serum was extracted by centrifugation (3,000 rpm×10 min) and stored at –80°C until biochemical analyses were performed. The right femora were dissected out and wrapped in gauze, which was soaked in phosphate buffered saline (PBS) and stored at –20°C for μCT and biomechanical strength evaluation.
Dual energy x-ray absorptiometry (DXA) Analyses
In vivo whole-body and femoral DXA scans were performed using DXA equipped with regional high-resolution scan software for small animal (Hologic QDR series, Hologic, USA). The same operator scanned all animals at baseline and after 10 weeks of experimental period. Before performing in vivo scan, the rats were anesthetized with Ketapex:Xylazil (1:1). Bone mineral area (BMA), bone mineral content (BMC) and bone mineral density (BMD) of the whole body and femur were evaluated. Before measurements were made, a tissue calibration scan was performed with the Hologic phantom for the small animal. Total BMD was calculated as bone mineral content/area.
Micro-CT analyses
The right femora that have been stored at -80°c were thawed to room temperature prior bone scanning. Femora were scanned by μCT (Skyscan 1176, Bruker, Belgium) at a pixel size of 18μm. Scanning conditions, the x-ray tube potential of 65 kV and 380μA, rotation step of 0.5 degrees were kept identical for all the tests. The volume of interest (VOI) was set to 200 slices and the region of interest (ROI) selected was at the metaphyseal region located approximately 1 mm below the distal growth plate, extended towards the proximal end of the femur. The VOI was assessed for structural parameters including trabecular bone volume (BV/TV), trabecular number (Tb/N), trabecular separation (Tb.S), trabecular thickness (Tb.Th), structural model index (SMI), and connectivity density. Cortical bone of the femur was evaluated by acquiring 200 slices of VOI located approximately 9 mm below the distal growth plate. Cortical parameters included total cross-sectional area inside the periosteal envelope (Tt.Ar), cortical area (Ct.Ar), cortical area fraction (Ct.Ar/Tt.ar) and average cortical thickness (Ct.Th). The analyses of trabecular and cortical bone parameters were performed using CTAn software (SkyScan CTAn, Bruker microCT, Belgium).
Bone mechanical testing
Biomechanical properties of femoral bone were evaluated using three-point bending test method (Shimadzu, Autograph AGS-X 500 N, Japan) controlled by proprietary software (Trapezium Version 1.4.2, Shimadzu). The femurs were cleaned from soft tissues and the total length and width of the midshaft were measured with digital caliper. The right femur from each rat was placed centred on an adjustable two-point block jig spaced at 5 mm apart, with a load sensor attached to a bending punch on the crosshead. Force was applied at a rate of 10 mm/min to the mid-shaft of the femur diaphysis until a break was determined by measuring a reduction in force. The testing parameter recorded included both extrinsic and intrinsic properties. Whole-bone mechanical properties were determined by the extrinsic parameters (force and displacement) while the measurement of the bone material were determined by intrinsic parameters, including stress, strain and Young modulus of elasticity. The slope of the stress-strain curve represents the Young modulus of elasticity or the elastic modulus. The elastic modulus is an indicator of intrinsic stiffness or bone’s ability to resist deformation.
Statistical analysis
Statistical analysis was performed using Statistical Package for the Social Sciences (SPSS, version 23.0, Chicago, USA). Distribution of the data was assessed using Shapiro-wilk test. Analysis of variance (ANOVA) followed by either Tukey’s or Dunnett’s T3 post hoc test was used to detect between-group differences in the variables of interest. P-values less than 0.05 was considered as statistically significant. All data were expressed as mean ± standard error of the mean (SEM).
Results
Before the start of the experiments, total BMA, BMC and BMD of the whole body and femur did not differ significantly in all groups (Table 1). Compared with baseline, total BMA, BMC and BMD were increased in all groups, but the increase was lower in orchidectomized and degarelix-induced groups. Following 10 weeks of experimental period, both orchidectomy and degarelix administration resulted in a significant decrease in whole body BMC, BMD and femoral BMD (p < 0.05) when compared to SHAM group. Femoral BMC was reduced significantly in ORX group (p < 0.05) but the changes were not significant in DGX group. Treatment with testosterone was able to prevent the reduction in whole body BMD and femoral BMC in orchidectomized groups (p < 0.05) but no significant changes were found in orchidectomized and degarelix-induced rats supplemented with EL extract.
Whole body and femur BMA, BMC and BMD in experimental rats
Whole body and femur BMA, BMC and BMD in experimental rats
The data is presented as the mean and standard error of the mean. *Significant difference compared to baseline group; aSignificant difference compared to SHAM group; bsignificant difference compared to ORX group; csignificant difference compared to ORX + TES group.
At 10 weeks of orchidectomy and chemical castration with degarelix, femoral metaphyseal trabecular bone quality was decreased as indicated by a significant reduction in trabecular bone volume and trabecular number, and a significant increase in trabecular separation when compared with SHAM group (p < 0.05). A significant reduction in connectivity density was observed in ORX group while the increase in SMI was found in DGX group when compared to SHAM group (p < 0.05). There were no statistical differences in trabecular thickness by the end of the experimental period in all groups. Treatment with testosterone prevented the degenerative changes at the distal femur as indicated by the improvement in the trabecular number, trabecular separation and connectivity density in both orchidectomized and degarelix-induced groups. While, improvement in trabecular bone volume with testosterone was only observed in orchidectomized group (p < 0.05). Trabecular bone volume, trabecular number and connectivity density of distal femur in EL-treated groups showed trends towards improvement when compared with respective negative control groups, but did not reached the level of statistical significance. Only degarelix-induced group treated with 100 mg/kg EL extract exhibited a significantly lower SMI when compared to DGX group (Fig. 1). Representative reconstructed 3-D images generated from μCT scan of the femur metaphysis showed that the degenerative changes were prevented by EL extract but at a lower efficacy than that of testosterone (Fig. 2).
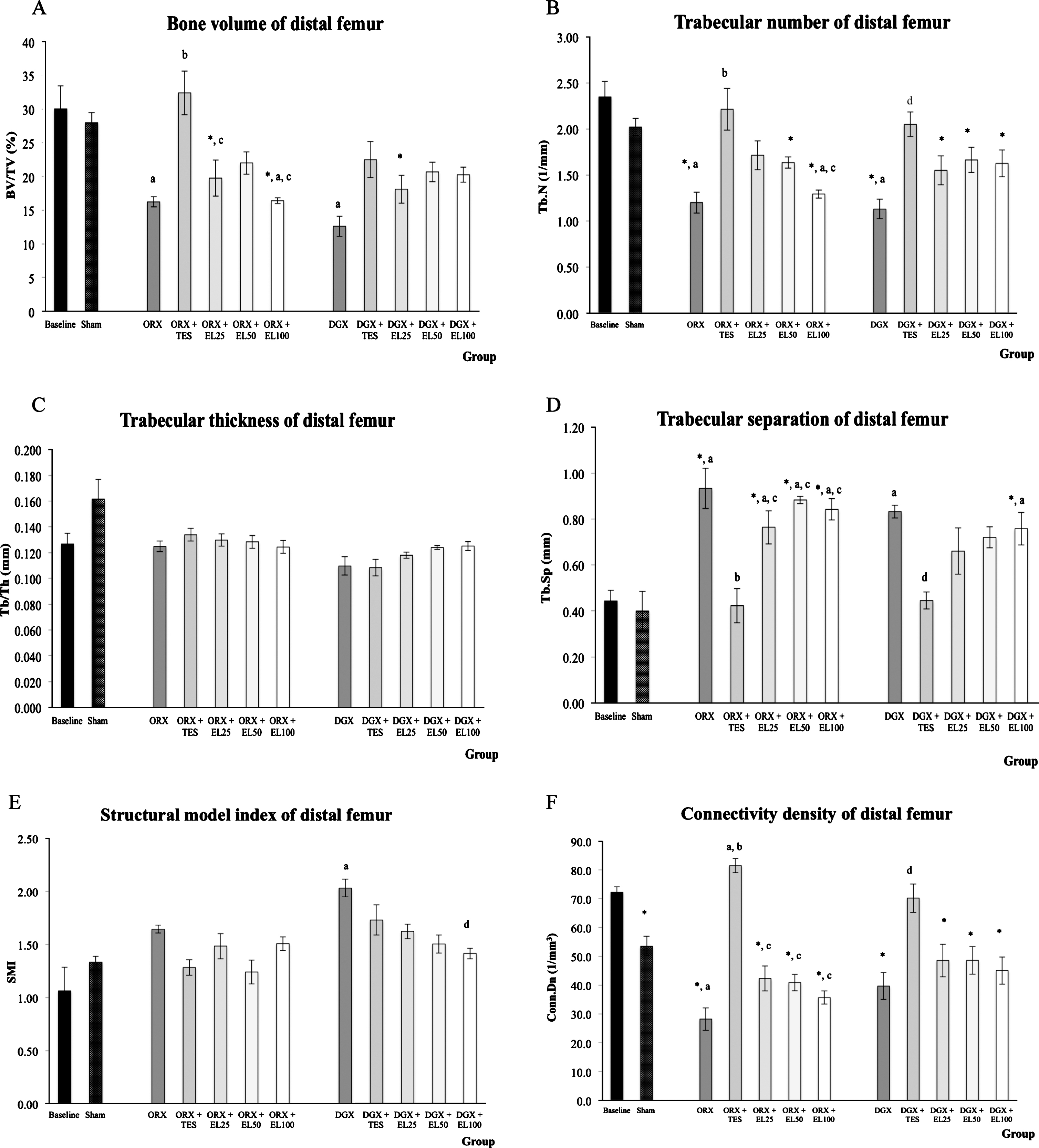
Trabecular bone structural indices in distal femur metaphysis. The bar charts (A-F) show trabecular bone structural indices at the distal femur metaphysis evaluated using μCT. The data is presented as the mean and standard error of the mean. *Significant difference compared to baseline group; aSignificant difference compared to SHAM group; bsignificant difference compared to ORX group; csignificant difference compared to ORX + TES group; dsignificant difference compared to DGX group.

Representative three dimensional micro-CT images of trabecular microarchitecture of distal femur metaphysis. The degenerative changes in trabecular structure due to orchidectomy and degarelix administration were prevented by EL extract but at a lower efficacy than that of testosterone.
Analysis of cortical bone at femoral diaphysis demonstrated that both orchidectomy and degarelix administration resulted in the reduction of total cross-sectional area inside the periosteal envelope, cortical bone area and cortical thickness when compared to SHAM group (p < 0.05). No alteration was found in the cortical area fraction in response to ORX and degarelix administration at the end of 10 weeks of experimental period. Testosterone treatment to orchidectomized group significantly prevented the reduction in total cross-sectional area inside the periosteal envelope, cortical bone area and cortical thickness parameters but the changes in degarelix-induced group treated with testosterone did not reach statistical significance. Interestingly, EL supplementation at the dose of 50 and 100 mg/kg was able to prevent the adverse changes to the cortical thickness in both orchidectomized and degarelix-induced groups. However, supplementation with 25 mg/kg of EL extract was only able to significantly improved cortical thickness in orchidectomized group (Fig. 3). Although EL supplementation did not cause any significant impact on total cross-sectional area inside the periosteal envelope and cortical bone area, its effect on cortical thickness was similar to that of testosterone.

Cortical bone structural indices in femur diaphysis. The bar charts (A-D) show cortical bone structural indices at the femur metaphysis evaluated using μCT. The data is presented as the mean and standard error of the mean. *Significant difference compared to baseline group; aSignificant difference compared to SHAM group; bsignificant difference compared to ORX group; csignificant difference compared to ORX + TES group; dsignificant difference compared to DGX group.
Three-point biomechanical testing of the femur mid-shaft revealed a significant reduction in cortical strength in both ORX and DGX group as indicated by the decrease in maximum force, stress, strain and stiffness value (p < 0.05), but the changes in Young Modulus of elasticity did not differ significantly when compared to SHAM group. Maximum displacement value was not statistically different in all experimental groups at 10 weeks of treatment. Treatment with testosterone was able to prevent the reduction in maximum force value in orchidectomized group and stiffness value in both orchidectomized and degarelix-induced groups (p < 0.05). Testosterone treatment also prevented the reduction in maximum stress and Young modulus of elasticity value in degarelix-induced group when compared to DGX group (p < 0.05). Compared to ORX group, supplementation of EL extract to the orchidectomized rats at the dose of 50 and 100 mg/kg significantly prevented the reduction in maximum stress value (p < 0.05). Supplementation of EL extract at the dose of 25 mg/kg resulted in a significantly higher maximum stress and stiffness value when compared to DGX group but Young Modulus of elasticity was significantly higher in degarelix-induced group supplemented with 100 mg/kg EL extract (Fig. 4).
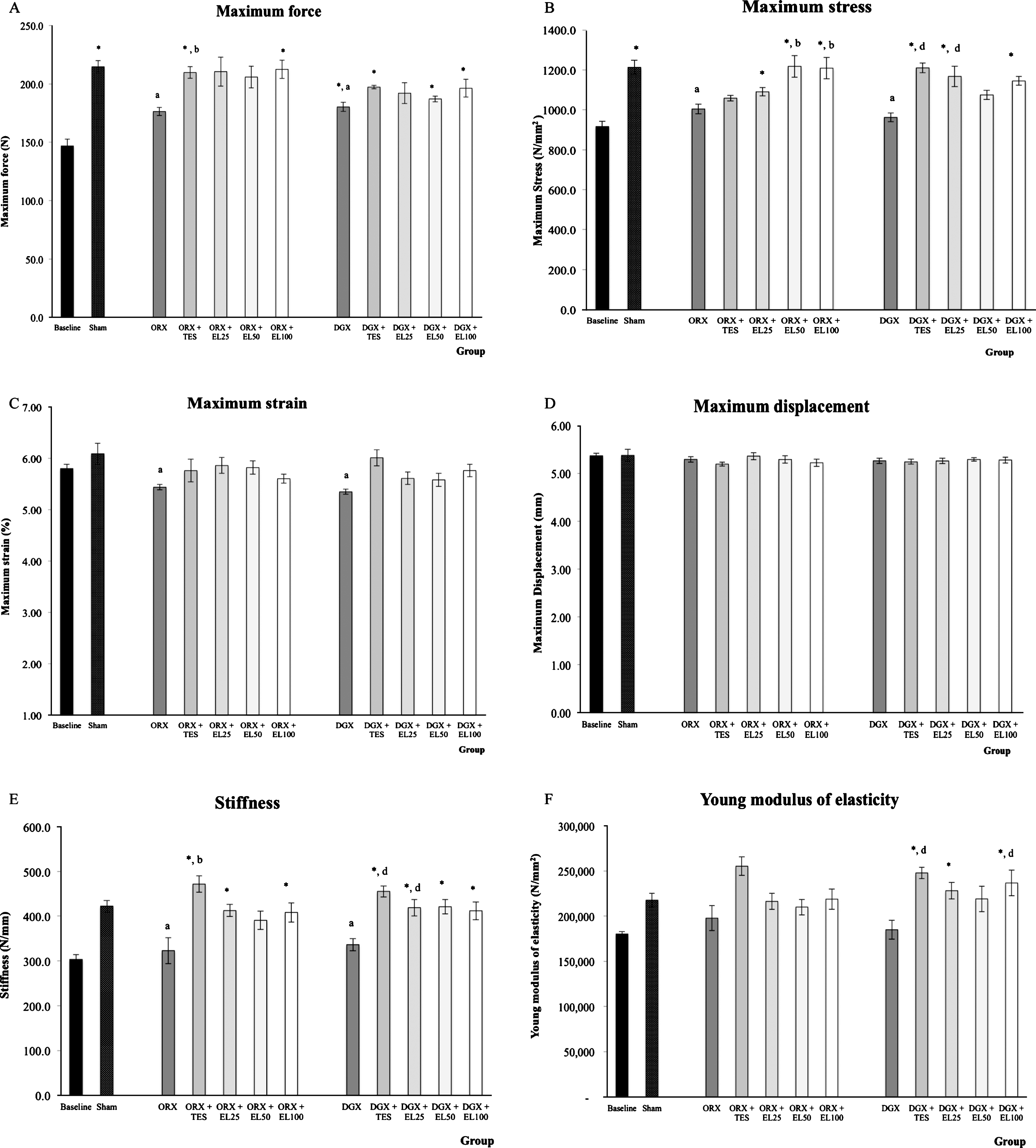
Mechanical strength of the femur midshaft. The bar charts (A-F) show mechanical strenth of the femur midshaft evaluated using 3-point bending method. The data is presented as the mean and standard error of the mean. *Significant difference compared to baseline group; aSignificant difference compared to SHAM group; bsignificant difference compared to ORX group; csignificant difference compared to ORX + TES group; dsignificant difference compared to DGX group.
Bone density, microarchitecture and mechanical strength were evaluated in orchidectomized and intact degarelix-induced rats supplemented with three different doses of standardized quassinoid-rich EL extract for 10 weeks. In the present study, for the first time, we have managed to demonstrate administration of degarelix, a GnRH antagonist, as an alternative method for inducing osteoporosis in male rats. We suggested that, apart from orchidectomy, this model is an appropriate model for male osteoporosis, as it closely resembles aging men with osteoporosis. This is because the testes are left intact, as opposed to them being removed in orchidectomy model. Studies on the effect of crude aqueous EL extract in orchidectomized rat model have been extensively carried out [26–28, 30]. In the light of these reports, we anticipated that the use of EL extract active compounds in different castration models may unveils the true potential of EL as an anti-osteoporotic agent.
Dual energy x-ray absorptiometry was reported to be a sensitive tools to measure time-related differences in bone mass or density [31]. However, DXA measurements should not be the only criteria used for assessing bone quality though it is considered as the gold standard in assessing the risk of osteoporosis [32]. In this study, we found that whole body and femoral BMD were adversely affected by orchidectomy and degarelix administration following 10-weeks of experimental period. Femoral BMC was reduced in orchidectomized rats but not in degarelix-induced rats. By the end of the treatment period, testosterone administration prevented the whole body BMD and femoral BMC reduction in orchidectomized rats. However, the supplementation of quassinoid-rich EL extracts was unable to reveal a significant improvement on the bone densitometry parameters.
In addition to bone densitometry, bone microarchitecture plays an essential role in determining bone quality. The assessment of trabecular and cortical bone microarchitecture serves an additional insight into the skeletal response to treatment [33]. A number of studies have reported the deterioration of trabecular bone microarchitecture in orchidectomized rats [34, 35]. In the present study, bone structural indices including trabecular bone volume, number, and separation were compromised in response to orchidectomy and degarelix administration. The changes in the structural indices due to testosterone deficiency may lead to increased bone porosity which can be revealed by the reduced connectivity density [36].
To our knowledge, this is the first study on the effect of standardized quassinoid-rich EL extract on bone microarchitecture of chemically castrated rats. As expected, testosterone treatment was found to be more effective than EL extract in preventing trabecular bone deterioration in the femur. Noteworthily, there was a trend towards higher trabecular bone volume, number and connectivity density in both orchidectomized and degarelix-induced rats following 10 weeks supplementation of quassinoid-rich EL extract. In addition, the deterioration of trabecular bone volume, number and separation in degarelix-induced groups supplemented with quassinoid-rich EL extracts was halted as no significant changes in these parameters was observed when compared to SHAM group. This data indicated that EL supplementation may produces a significant effect in bone if longer experimental period were allowed. Data from the present study also showed that the 100 mg/kg of standardized quassinoid-rich EL extract was able to improve SMI in degarelix-induced rats. The ability of EL extract to improve SMI suggests that EL is capable of inhibiting the perforation and thinning of the trabecular plates, which in turn reduces their transition to rod-like structures [37].
In a previous study, supplementation of crude EL extract at the doses of 30 and 60 mg/kg for 6 weeks failed to protect bone from orchidectomy induced-changes and even worsened all the bone structural indices [28]. Some protection on the trabecular bone volume was only observed by the supplementation of 90 mg/kg crude EL extract. The active compounds found in the crude aqueous extract used in the previous study were glycosaponin (41.1%), eurypeptide (22.0%) and eurycomanone (1.6%). Eurycomanone was reported to be the major and the most potent form of quassinoid in EL root extract [29]. However, the eurycomanone was detected in very low concentration in the crude aqueous EL extract, and this may explain the less prominent effects of EL on bone microarchitecture found in the previous studies.
The measurement of cortical area and cortical thickness are considered as strong predictors of bone strength and resistance to fracture [38]. Total cross-sectional area, cortical bone area, cortical bone area fraction and cortical thickness are the variables that should be used to describe cortical bone morphometry [39]. Assessment by micro-CT showed that the total area inside the periosteal envelope, cortical area and cortical thickness were significantly decreased after orchidectomy and DGX administration. Previous studies have demonstrated that the decrease of cortical area and cortical thickness in the orchidectomized rats is indicative of cortical thinning [40, 41].
In the present study, we found that cortical bone was also compromised in addition to trabecular bone deterioration at 10 weeks following orchidectomy and degarelix administration. Treatment with testosterone increased the total area inside the periosteal envelope, cortical area and cortical thickness in orchidectomized rats. Interestingly, supplementation of quassinoid-rich EL extracts at all doses resulted in the improvement of cortical thickness in the orchidectomized rats whereas the cortical thickness of degarelix-induced rats was improved by the supplementation of 50 mg/kg and 100 mg/kg of EL extract. The improvement of cortical thickness may be associated with an increased in periosteal bone formation on the external surface of the cortical bone, enhancement of bone formation and/or a decrease in resorption on the endocortical surface [42].
Bone quality or tissue fragility can be characterized by measurements of intrinsic biomechanical properties which includes maximum stress, strain and Young modulus [43]. Meanwhile the extrinsic parameters, maximum force, displacement and stiffness reflect the property of whole bone. In our previous study, bone strength parameters were not significantly altered at 6 weeks post or chidectomy [44]. Another study by Díaz-Curiel, De La Piedra [45] have also found that there was no significant difference in the biomechanical strength of the orchidectomized rats. In the present study, the mechanical strength of the midshaft femur was equally compromised by orchidectomy and degarelix administration at 10 weeks of experimental period. This indicated that 10 weeks of androgen deprivation in experimental animal was sufficient to affect bone strength.
Three-point bending test showed significantly lower maximum force, stiffness, stress and strain in both testosterone-deficient rat models. Our findings from the biomechanical testing confirmed the thinning of cortical bone found in the microarchitecture assessment. The bones of the orchidectomized rats treated with testosterone showed significant improvement in maximum force, stiffness and Young modulus while testosterone treatment in the degarelix-induced rats significantly improved bone stiffness, stress and Young modulus. The supplementation of quassinoid-rich EL extracts at the dose of 50 mg/kg and 100 mg/kg was able to improve stress value in the orchidectomized rats. Stress is an intrinsic parameter that is defined as the strength of the bone tissue under a given loading condition. Significant improvement of stiffness and stress value was found in degarelix-induced rats supplemented with 25 mg/kg of quassinoid-rich EL extract. Stiffness is an extrinsic parameter that is also used to assess mechanical integrity of bones. The increase in stiffness indicates an increased in bone mineralization [46]. An effective treatment for bone fragility should improve the extrinsic biomechanical properties and at the same time preserved the intrinsic properties of the bones [43].
The standardized extract used in this study contain more than 10 folds of eurycomanone and its analogue, 13α(21)-epoxyeurycomanone, 13α,21-dihydroeurycomanone and eurycomanol. The bioactive compounds of EL extract especially eurycomanone was postulated to be able to increase the testosterone synthesis via its direct effect on the hypothalamic-pituitary-gonadal (HPG) axis [29]. The production of testosterone by the Leydig cells in the testes is stimulated by the luteinizing hormone (LH) secreted from the anterior pituitary [47]. The secretion of the pituitary gonadotropin, the LH and FSH is regulated by the hypothalamus. Eurycomanone was reported to be responsible for the increase in the plasma testosterone and the two gonadotropins, LH and FSH synthesis in intact male Sprague-Dawley rats [29]. Testosterone may exerts its effect via signaling through the androgen receptors found on bone skeletal surface or indirectly via aromatization to estrogen [9]. In the degarelix-induced model, it is possible that the eurycomanone from the quassinoid-rich EL extract worked directly on the HPG axis, thus increasing testosterone synthesis.
In this study, we have shown that, quassinoid-rich EL extract has the potential to preserve bone microarchitecture and strength in orchidectomized and chemically castrated rats. Though the efficacy of EL extract in preventing trabecular bone microarchitecture is less superior to testosterone, the ability of EL extract in preventing the deterioration of cortical bone and mechanical strength is on par with testosterone treatment. This is important since biomechanical strength is the best parameter that reflects the bone function and risk of fracture. Further investigation on the exact mechanism by which quassinoid-rich EL extract exerts its anti-osteoporotic potential is warranted to clarify the influence of EL extract on bone metabolism.
Footnotes
Acknowledgments
We are grateful to the staff of Pharmacology Department, UKM Medical Centre for their technical support. We thank Universiti Kebangsaan Malaysia for providing the grant for this study.