
Abstract
In this study, the electrical resistance of the whole body and histological changes of skeletal muscle were investigated in rats according to the increase in radiation dose. A total of 15 male Sprague–Dawley rats (5-weeks-old) were randomly divided into 5 groups (each, n = 3). Each group received 1 Gy, 5 Gy, 10 Gy and 20 Gy systemic exposure, and the non-irradiated group was used as a control for morphological comparison. After attaching an electrode clip to the forelimb of the rat, an AC frequency was applied before and 4 days after irradiation using an impedance/gain-phase analyzer, and the measurement system was automatically controlled with LabVIEW. Comparing to before irradiation after 4 days, the difference in the average impedance values at 1 Gy, 5 Gy, 10 Gy, and 20 Gy was 1188±989 ohm, 3076±2251 ohm, 7650±6836 ohm, and 10478±6250 ohm, respectively. By comparing the normal group and the experimental group, muscle fiber atrophy and collagen fibers around blood vessels were observed (p < 0.05, control group vs 5 Gy or more high-dose group). These results confirmed the previously reported morphological changes of skeletal muscle and our hypothesis that whole-body impedance measurement enables to reflect tissue changes after irradiation.
Introduction
The intercellular space and the inside of the cell are ionic conductors, so they are usually represented as resistive components, whereas cell membranes are represented as capacitive components because of their structure. Therefore, in vivo tissues are generally known to be resistant to electricity [11].
Recently, the effects on organelles, cytoplasm, and extracellular matrix(ECM) caused by indirect action are also receiving attention [38]. Cell-to-cell signaling occurs at the cell membrane’s junction between cells and the physical connection between cells and the ECM [3]. The ECM not only functions to physically protect and support cells or tissues by forming cell junctions and networks, but also plays an important role in the formation, differentiation and homeostasis of tissues [37]. When such an inflammatory and anti-inflammatory response occurs, ECM remodeling occurs by various immunomodulatory mechanisms in the acute and chronic phases [32]. Tissues exposed to ionizing radiation cause an inflammatory response in the acute phase and remodel the ECM composition into a loose and hydrated environment for cell infiltration and recovery.
The living body has its own electrical properties due to various differences, such as the arrangement of cells and the shape of tissues, and when a biological change occurs in the tissue, the intrinsic electrical properties also change [16, 27]. The electrical resistance of a living body varies depending on the size, spacing, composition, structure, and matrix of the cell membrane, increases as the capacitance of the cell membrane increases, and decreases as the amount of water and electrolyte in the tissue increases [7]. In addition, the electrical resistance characteristics of living tissue spread through the tissue in a way that the electric current depends on the frequency, the common theory is that low-frequency current(∼kHz) mainly flows through the space between cells, and high-frequency current(several hundred kHz, MHz) is it flows through the cell membrane [18, 21].
Changes in electrical resistance inside and outside cells due to ionizing radiation are most affected by changes in water content in the body. The extracellular space has a lower water density than the cytoplasm, and if the cell membrane is destroyed after exposure to radiation and the hydrating substance in the cytoplasm flows into the ECM, it will be greatly affected by electrical resistance [9].
Exposure to ionizing radiation in the living body causes physical and chemical damage, resulting in intracellular biochemical reactions such as DNA damage and repair, cell death, stasis of the dividing cell cycle, changes in cell signaling, and oxidative stress response [2]. The most abundant fiber protein in ECM is collagen, which accounts for up to 30% of the total protein mass of multicellular animals [12]. When living body is exposed to ionizing radiation, the physical and chemical changes are terminated within microseconds, but reactive oxygen species (ROS), cytokines, and chemokines continuously interact to generate inflammatory and anti-inflammatory responses [26]. On that basis, the effects of ionizing radiation in normal tissues will form different environments in the intracellular fluid, cell membrane, and ECM according to the change in dose. And if more structural changes occur in cells, tissues, and ECM, and more water-soluble substances fill the ECM, it is expected that electrical resistance will change [10, 35].
We expected that the difference in the amount of collagen in the extracellular matrix (ECM) produced depending on the dose and the difference in electrical resistance would occur due to the change in cell size. In addition, it was hypothesized that ionizing radiation reduces the capacitance by destroying the cell membrane, and the electrical resistance in vivo decreases as the amount of water and electrolyte passing through the inside and outside of the cell increases.
Until recently, studies investigating the change of impedance in living body by ionizing radiation have been continuously reported. However, most of these studies mainly conducted local irradiation with high doses of 15 Gy or more to specific organs, the focus was on morphological changes and impedance changes in tissues over time [28]. There has been little progress in whole-body radiation studies on rats, especially impedance studies. However, the relationship between changes in electrical resistance and changes in skeletal muscle tissue before and after whole-body irradiation in living rat was not reported. In our previous paper, rupture of pork tenderloin tissue occurred severely at 10 Gy. In this study, we predict that there would be a lethal radiation dose at 10 Gy. And we try to study the impedance change at a lethal radiation dose when irradiating whole body radiation. In addition, X-ray was irradiated on the whole body of rat to compare the differences in electrical resistance according to dose and to confirm histological changes in skeletal muscle.
Method
Animal groups
Animal care and all experiments were approved by the Institutional Animal Care and Use Committee of Pusan National University Hospital (Approval No. ED-PNU2017-0080) and carried out in compliance with university’s scientific research guidelines and regulations. Male Sprague–Dawley rats that were 5-week-old (n = 15) and approximately weighing 144∼157 g were used. The rats were divided into 5 groups, with 3 in each cage. Rats were acclimatized in individual cages for 7 days prior to experimentation in a room with a constant temperature of 23±2°C and 55% relative humidity on a 12/12 h light-dark cycle. Animals were allowed free access to food and water ad libitum. During irradiation and measurement of electrical impedance characteristics, oxygen was supplemented while anesthesia was induced by isoflurane inhalation (induction 5%, maintenance 2%). Then, the experimental animals were euthanized 4 days after irradiation.
Irradiation and electrical impedance measurement
Radiation doses of 1, 5, 10, 20 Gy was irradiated to rats using a linear accelerator (Versa HD, Elekta AB, Stockholm, Sweden) 6 MV X-ray. Rats were irradiated by whole body with a collimation field 100×100 cm2, SAD 100 cm. To measure electrical resistance, anesthetize rat on an acrylic plate capable of continuous anesthesia, an electrode clip was attached to the forelimb of the rat (Fig. 1a). In order to obtain accurate measurement values, the electrode attachment location was wiped with an alcohol swab to minimize resistance. Using an impedance measuring instrument (Impedance/Gain-phase analyzer 4194A, Agilent, USA), an AC frequency was applied from 1 kHz to 200 kHz before and 4 days after irradiation (Fig. 1b). The change in Impedance Absolute Value of rat was measured repeatedly 10 times at 3 minutes intervals, LabVIEW (National Instruments, Austin, TX) real-time interface system was used for automatic control of the measurement system and data processing (Fig. 1c). Then, the measurement results before and 4 days after irradiation were analyzed to confirm the relationship between impedance changes.
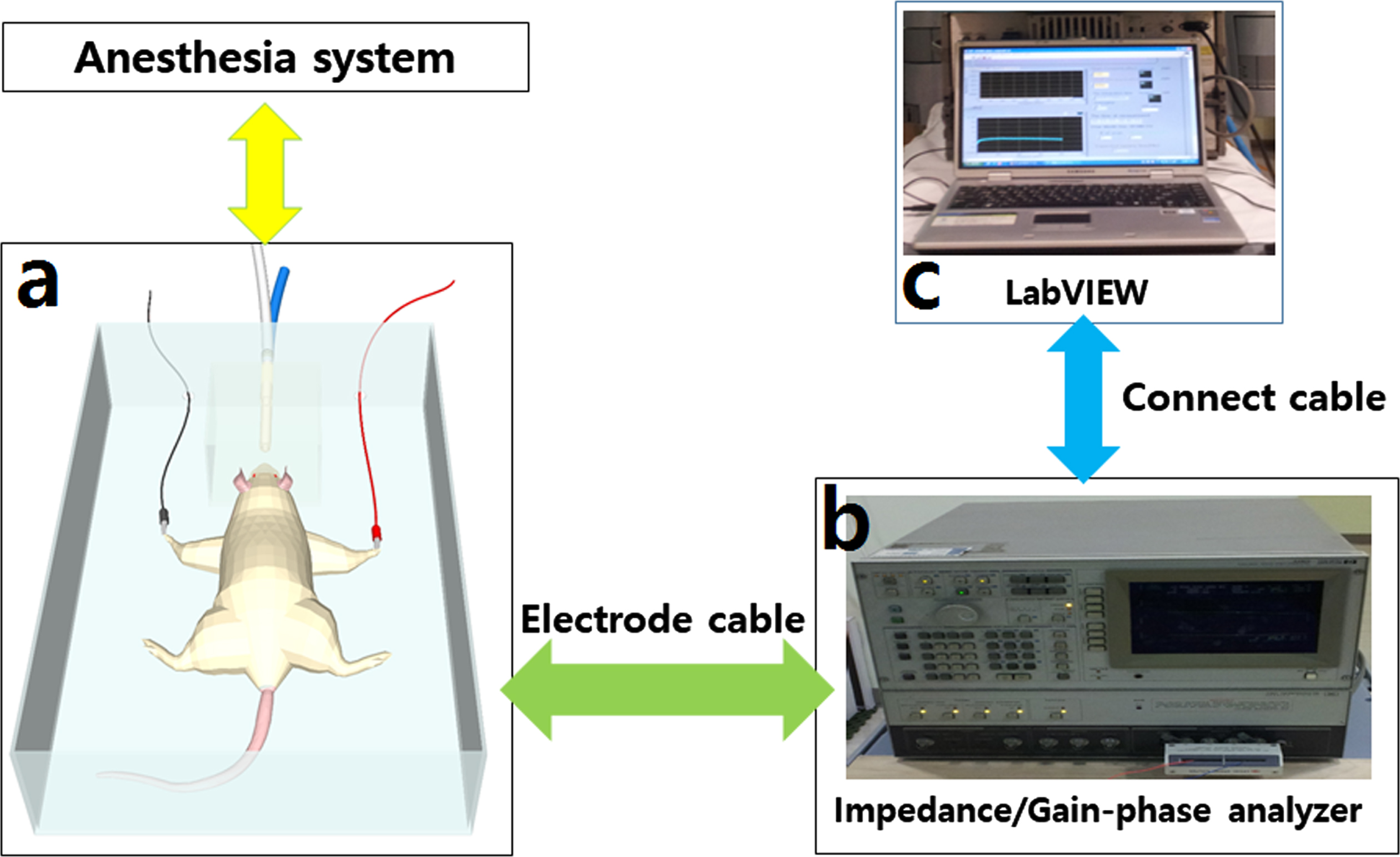
(a) The rat was anesthetized and electrodes were attached to the forelimbs. (b) Electrical resistance was measured with Impedance/Gain-phase analyzer before and 4 days after irradiation. (c) The data was processed with LabVIEW.
The irradiated rat’s quadriceps femoris were divided into two parts and divided into a longitudinal section and a cross section, respectively. Quadriceps tissues were fixed in 10% formalin, embedded in paraffin and sectioned at 3μm thickness. Changes in tissues pathology and collagen deposition were observed by hematoxylin-eosin (H&E) staining and Masson’s trichrome (MT) staining, respectively. We selected abnormal area in each slice for representative images taken under 20×, 40×objective AperioScanScope CS2 digital slide scanner (Leica Microsystems GmbH, Wetzlar, Germany). Next, we captured it using Aperio Image Scope viewing software (Leica Biosystems Imaging, Vista, CA, USA). Muscle fiber diameters in the quadriceps were measured from images captured from H&E staining of longitudinal sections. Muscle fibers with distinct cell border in the longitudinal direction were selected, and transverse or oblique borders were excluded. The degree of tissue collagen region, which is the blue region from the MT-stained area, and the diameter of muscle cells were evaluated as Image J (NIH image J).
Statistical analysis
Statistical analysis was done using the SPSS 26.0 software (SPSS, Chicago, IL, USA). Impedance data are represented by mean±standard deviation (mean±SD). The differences in muscle fiber diameter and collagen ratio in each group were compared as mean values. The difference between the Impedance Absolute Value was analyzed using the Wilcoxon Signed Ranks test. The difference in the mean values of muscle fiber diameters was followed by one-way ANOVA followed by Dunnett’s post hoc test. The difference in the mean values of the collagen ratio was determined by a Kruskal-Wallis nonparametric test (N < 30 per group) due to the small sample size. And when p < 0.05, it was considered to indicate a statistically significant difference.
Results
Changes in the electrical characteristics of the impedance according to the radiation dose
Impedance is a physical quantity that indicates the degree of difficulty in flowing an electric current in a biological material and is frequency dependent. Figure 1 shows the impedance and phase value of whole body. We analyzed impedance and phase at frequencies from 0 to 200 kHz. In the frequency of 1 kHz to 200 Hz, the magnitudes of the impedance and phase show a decrease and is proportional to the frequency, as shown in Fig. 2.
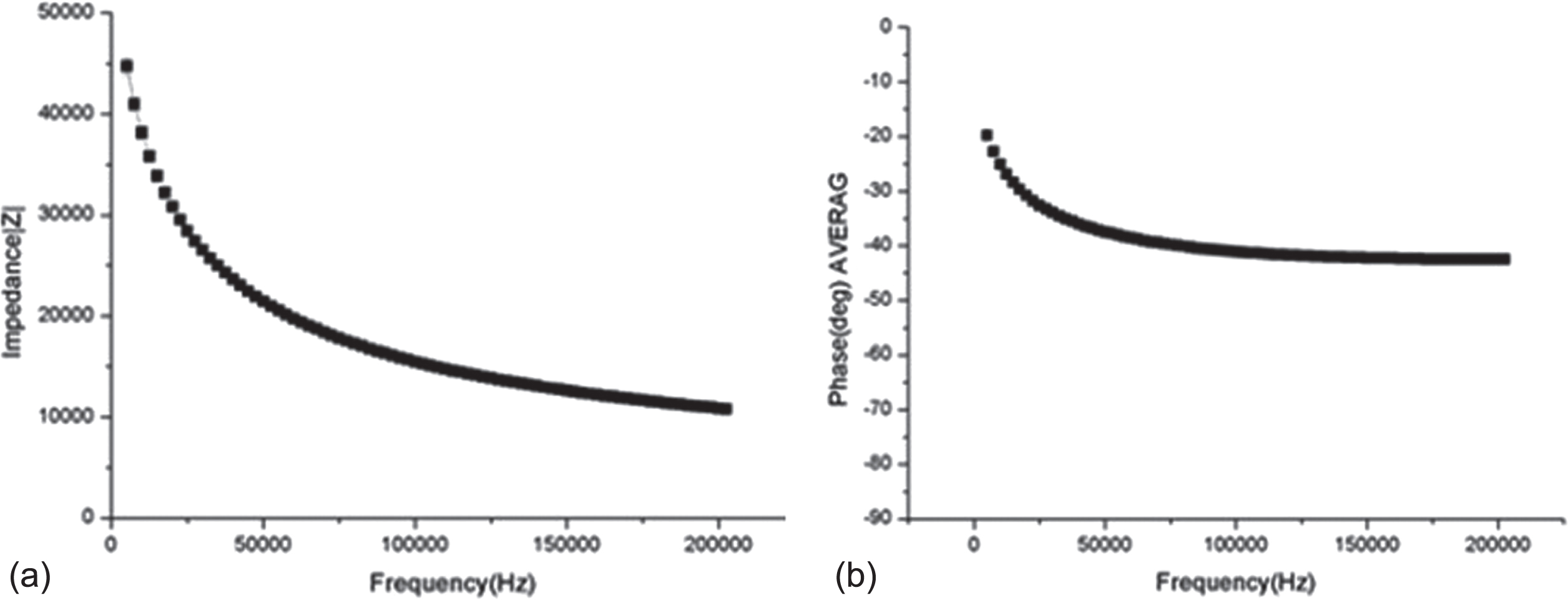
The impedance and phase value of whole body. Graph of the impedance (a) and (b) phase of whole body from 1 kHz to 200 kHz at the right graph.
AC frequency is applied from 1 kHz to 200 kHz before and 4 days after irradiation, the results of measuring changes in the Impedance Absolute Value of rats are the same as shown in Table 1. The difference in impedance mean values before and after radiation irradiation was 1188±989 ohm in R1 group, 3076±2251 ohm in R2 group, 7650±6836 ohm in R3 group, and 10478±6250 ohm in R4 group, as shown in Fig. 3.
Impedance Absolute Value result on data of measurement
R1 : 1 Gy R5 : 5 Gy R10 : 10 Gy R20 : 20 Gy *P < 0.05.

Comparison of Impedance Absolute Values of impedance between pre-irradiation and irradiation after 4 days of irradiation. R1 : 1 Gy; R2 : 5 Gy; R3 : 10 Gy; R4 : 20 Gy. Box plot representing the Impedance Absolute Value in the experimental (radiation expose) and positive control group (Wilcoxon signed rank test, alpha = 0.05).
Hematoxylin-eosin staining
Histological analysis through H&E staining was performed qualitatively. Figure 4 s a box diagram showing the cross-section of the quadriceps muscle and the diameters of muscle fibers by radiation dose. There was no histological difference between the control group and the 1 Gy group. However, in the 5 Gy, 10 Gy, and 20 Gy groups, polynuclei was found within muscle fibers (white arrow), and the boundaries of muscle fibers were irregular (yellow arrow). As shown in Fig. 4f, the diameter of muscle fibers decreased as the radiation dose increased. There was no significant difference between the control group and 1 Gy, but 5 Gy, 10 Gy, and 20 Gy were significantly decreased(muscle fiber diameter (μm): control group, 28.89±3.65; 1 Gy group, 28.33±5.21; 5 Gy group, 24.81±5.32; 10 Gy group, 24.69±5.44; 20 Gy group, 22.05±4.05; p < 0.05, control group vs 5 Gy or more high-dose group; n = 40 per group).
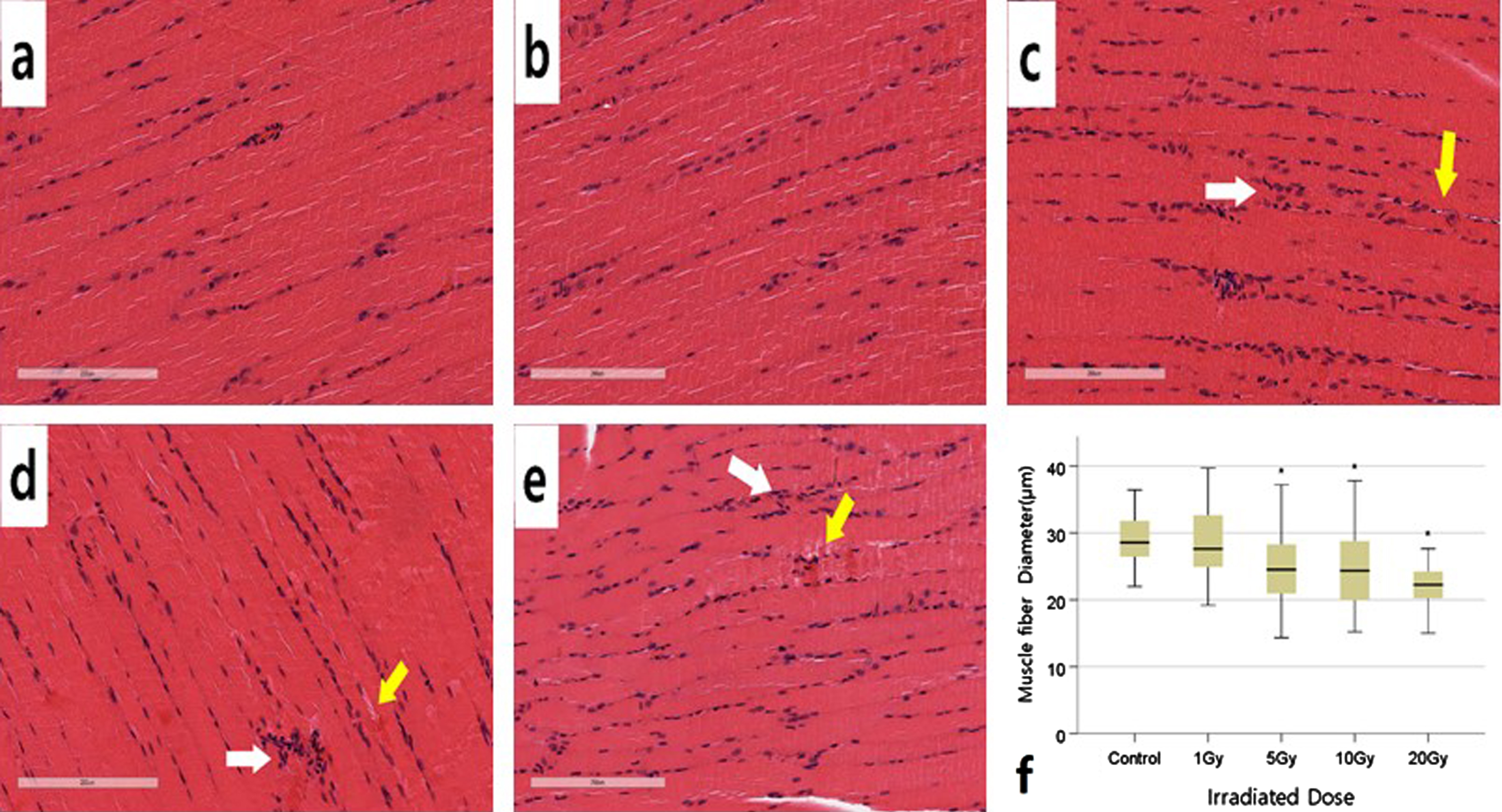
Representative Hematoxylin-eosin staining of quadriceps femoris muscle longitudinal sections and statistical analysis of muscle fibers 4 days after irradiation. (a) Control (b) 1 Gy (c) 5 Gy (d) 10 Gy (e) 20 Gy (f) The quantitative results were demonstrated as a box plot. *p < 0.05 significantly different from the normal group. n = 40 muscle fibers for each group. Data were analyzed using the one-way ANOVA, followed by Dunnett’s post hoc test. Magnification: x20.
To determine the difference in collagen fiber area by ionizing radiation dose, it was quantitatively analyzed through MT staining. Collagen fibers were stained blue by Masson’s trichrom, and the area of collagen fibers for the selected section increased with increasing radiation dose. Figure 5 shows the area of collagen fibers for a selected section in the quadriceps muscle, differences in areas between groups were compared and analyzed using box plots. Collagen fibers were mainly observed around the perivascular, and a small amount of collagen fibers were observed around endomysium and perimysium. The area of collagen fibers was not significantly different in the 1 Gy group compared to the group, but significantly increased in the 5 Gy, 10 Gy, and 20 Gy groups(area of collagen(%): control group, 0.22±0.12; 1 Gy group, 0.69±0.70; 5 Gy group, 3.18±3.60; 10 Gy group, 3.90±1.74; 20 Gy group, 7.11±5.39; p < 0.05, control group vs 5 Gy or more high-dose group; n = 3 per group).
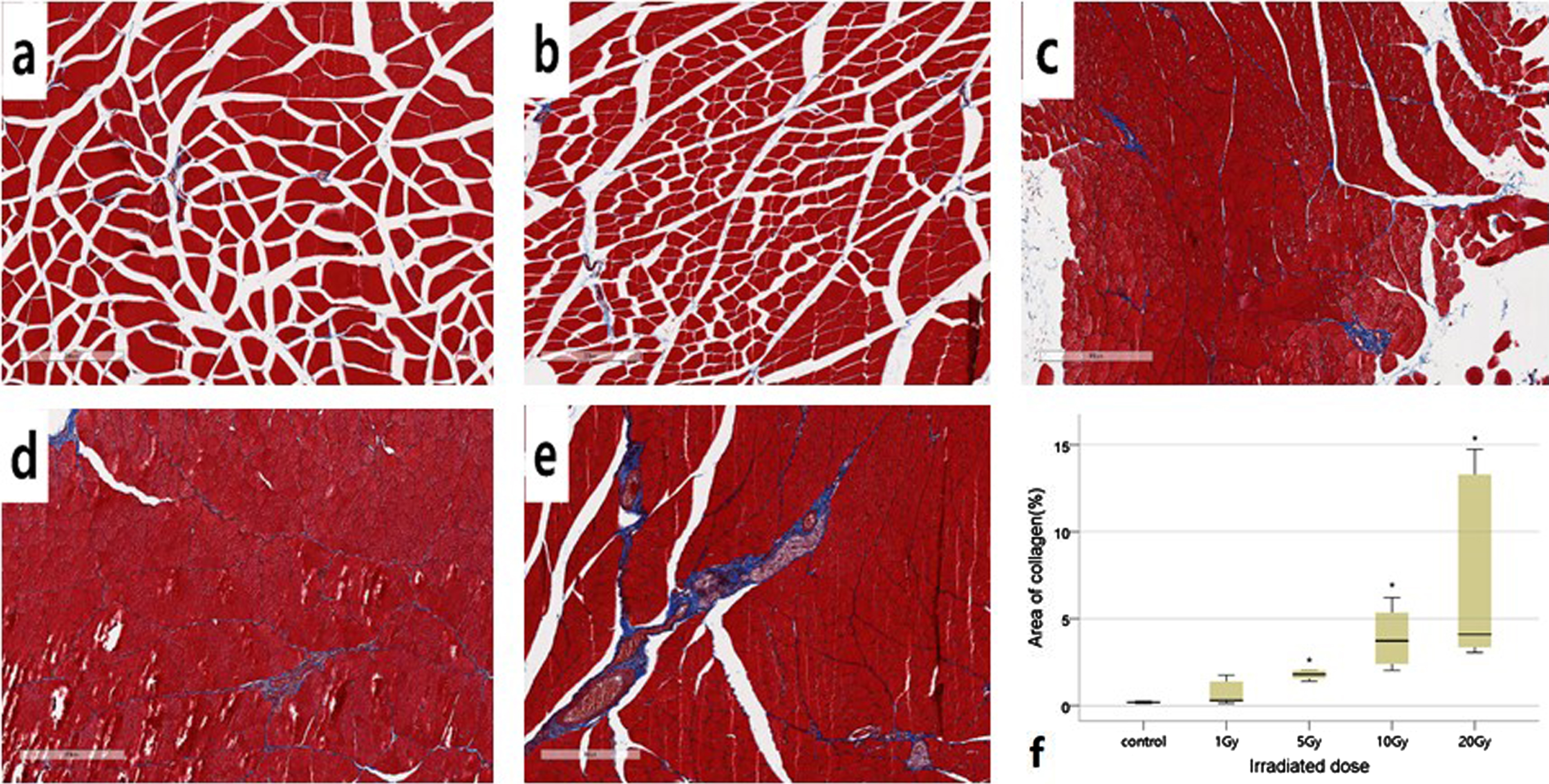
Representative Masson’s trichrome staining of quadriceps femoris muscle cross sections and statistical analysis of collagen fibers 4 days after irradiation. Collagen fibers were stained blue by Masson’s trichrom. (a) Control (b) 1 Gy (c) 5 Gy (d) 10 Gy (e) 20 Gy. (f) Analysis of collagen fiber area percentage in each dose. *p < 0.05 significantly different from the normal group, n = 6 slice for each group. Data were analysed using the Kruskal Wallis test. Magnification: x10.
The difference in impedance mean values before and after radiation irradiation was 1188±989 ohm in 1 Gy group, 3076±2251 ohm in 5 Gy group, 7650±6836 ohm in 10 Gy group, and 10478±6250 in 20 Gy group, as shown in Fig. 2. After 4 days of irradiation, the difference between the control group and the muscle fiber diameter (μm) was 1 Gy group, 0.56±5.01; 5 Gy group, 4.07±8.88; 10 Gy group, 4.19±6.06; 20 Gy group, 6.03±7.14, and as the dose increased, the muscle fibers atrophied. And the difference in the area of collagen(%) compared to the control group was 1 Gy group, 0.47±0.64; 5 Gy group, 2.96±3.61; 10 Gy group, 3.68±1.77; 20 Gy group, 6.88±5.35 and the area of collagen fibers increased as the dose increased. There were significant changes in the decrease in muscle fiber diameter and increase in the area of collagen fibers in the average value of the group irradiated with more than 5 Gy (p < 0.05). However, in the 1 Gy group, the impedance change increased to 1188±989 ohm, but there was no significant change in the morphological analysis.
Discussion
In vivo electrical signal analysis is non-invasive and easy to use at low cost, so it is widely used in various fields such as the body, electrocardiogram, and electromyography. However, there is no study on the relationship between the electrical signals and biological changes caused by whole-body irradiation of ionizing radiation. The impedance in vivo would change as the physical characteristics change depending on the dose of ionizing radiation and the type of organ [39]. It is expected that the electrical resistance will decrease if the intracellular resistance factor decreases due to physical changes and the hydrating substance increases due to changes in the intracellular and extracellular matrix. In vivo electrical resistance is determined by extracellular resistance, intracellular resistance, and cell membrane capacitance, damage to living tissue by irradiation causes a change in bioelectrical resistance in the tissue. Paulsen, Osterman, and Hoopes reported that as radiation dose increases, the cell membrane loosens, the extracellular fluid increases, and the impedance decreases [29].
When an electric current flow through a living body, it is divided depending on the individual resistance and volume of each tissue. Bone and adipose tissue are more electrically resistive than muscle and therefore do not affect changes in electrical resistance [19]. In this study, we tried to confirm the change in electrical resistance of the whole body due to the effect of ionizing radiation. However, there was a difficulty in reflecting all the damage of various type of organ. Therefore, it was attempted to reflect changes in electrical resistance of the whole body as changes in skeletal muscle on the following grounds. Skeletal muscle, the largest organ in the body, constitutes about 40% of the total body weight in humans [13]. When an electric current flow through a living body, it is divided depending on the individual resistance and volume of each tissue. Skeletal muscle has a large volume and low resistance, so most of the current flows through the skeletal muscle [10]. The trunk of whole body is of little importance in measuring systemic electrical resistance, although it is also flowed by other components that reflect small proportions, such as parenchyma and lymph [14]. It affects resistance only when the volume of adipose tissue exceeds the volume of skeletal muscle, but the effect is small [4]. And bone has very high resistance, so the effect on resistance is small [19].
So, Skeletal muscle which has the greatest influence on the electrical resistance of the whole body was selected [24]. Therefore, in this study, we expected that the measured impedance value at whole body would be representative of that measured at skeletal muscle. In other words, among tissues, skeletal muscle changes play the greatest role in the change in electrical resistance of the whole body, and changes in parenchymal tissue or lymphatic system have only a slight effect on electrical resistance, so this study was conducted only with skeletal muscle changes.
In this study, it was confirmed that there was a difference in electrical resistance of the whole body due to ionizing radiation dose, and it was confirmed that the difference in dose also caused differences in morphological changes in skeletal muscles. As a result of whole body irradiation, AC frequency is applied from 1 kHz to 200 kHz before and 4 days after irradiation, the difference in impedance mean values before and after radiation irradiation was 1188±989 ohm in R1 group, 3076±2251 ohm in R2 group, 7650±6836 ohm in R3 group, and 10478±6250 ohm in R4 group.
However, only changes in skeletal muscle were observed, and other factors that cause damage to the whole body were not identified. Due to the difference in radiation sensitivity, the degree of damage to skeletal muscle does not match the degree of damage in each tissue in the living rat. It was not confirmed whether the change in electrical resistance of the whole body only reflected damage to the skeletal muscle, or whether it reflects damage to systemic tissues or whether there was a difference due to recovery after damage. In order to clearly determine the causal relationship between changes in electrical resistance due to total body irradiation, additional measurements during real time irradiation and over time are required in using the same protocol. In addition, it is necessary to accumulate additional data on electrical resistance due to the degree of tissue damage and frequency changes in the future.
According to previous studies, Challapalli et al. reported that LD50/30 was 6.6 Gy when whole body was irradiated with a linear accelerator on the Albino Wistar Rat [5]. But, in our pilot study of this experiment, it did not survive more than 5 days when X-rays of 10 Gy or higher were whole body irradiated in Sprague-Dawley rats. Therefore, electrical resistance was measured before irradiation and 4 days after irradiation, and histological changes were confirmed by euthanasia. In our previous paper, rupture of pork tenderloin tissue is similar to skeletal muscle occurred severely between at 10 Gy [21]. We expected this to be related to the results of our pilot experiments.
Small fibers were found in the non-irradiated muscle fibers, but small fibers accounted for only 1% of the total in the irradiated muscles [30]. This means that new fibers are generated in the non-irradiated muscle, but there are few new small fibers in the irradiated muscle. Adams, Gregory R., et al. also reported that there was no compensatory hypertrophy up to 4 months when 25 Gy was applied to the hindlimb of Sprague-Dawley rats [1]. Jurdana et al. reported that adult skeletal muscle is radiation-resistant, whereas developing skeletal muscle is sensitive to radiation [20]. Radiation exposure during the developing skeletal muscle causes satellite cells to inactivate and interfere with hypertrophy, producing few small fibers [20]. Ghasemi et al. reported that Rat, aged 5 to 8 weeks, is 11–14 in human age [15]. On that basis, the rats used in this experiment are about puberty, and the small fiber can be said to be a fiber previously formed.
The muscle fiber diameter measured in the 10 Gy or higher dose group did not show compensatory hypertrophy or normal growth due to injury. Damage to other organs due to whole body irradiation may affect muscle fiber atrophy.
Cytokine such as Transforming growth factor-β (TGF-β1), an anti-inflammatory factor, act as growth inhibitory agents, regulate apoptosis in parenchymal cells, and stimulate collagen production in the ECM [26]. Intracellular cytokines stimulate anti-inflammatory effects at low doses of less than 1 Gy, and the incidence of apoptosis is higher than that of necrosis, but at high doses of 1 Gy or more, necrosis occurs and an inflammatory reaction occurs [34]. In the chronic phase, the ECM is converted into a dense, rigid environment with more collagen and other fibrous proteins, which is called fibrosis [40] [6]. Radiation-induced fibrosis (RIF) can occur in all organs exposed to skin and subcutaneous tissue, lungs, gastrointestinal tract, urogenital organs, and radiation, and RIF in muscles and liver may typically occur weeks and months after exposure to ionizing radiation [17, 22].
TNF-α released from liver fibrosis circulates systematically and can promote atrophy of skeletal muscle [23]. When skeletal muscle is damaged, muscle fiber atrophy and nuclei loss occur in the order of injury, inflammation, and regeneration [33]. It is said that even in adolescent mice, a decrease in muscle fiber size and loss of musclei are discovered 14 months after irradiation. However, in our study, as the radiation dose increased, the muscle fiber size decreased, polynuclei were observed in the muscle fibers in the 10 Gy or higher dose group, as shown in Fig. 4. Polynuclei deviates from normal peripheral sides of muscle fibers, it shows a process in which the nucleus is located within the muscle fibers and degeneration or regeneration proceeds.
In-vivo electrical signals are affected by changes in water content inside and outside the cell. Ionizing radiation interacts with the water-rich cytoplasm and blood to produce ROS through early indirect action. ROS damages adjacent vascular endothelial cells, causing an initial inflammatory reaction and changing blood vessel permeability [41]. Such damage to vascular endothelial cells is often detected in the liver in the early stages of radiation exposure, and is called veno-occlusive disease (VOD) [8, 31]. VOD activates myofibroblasts by intrablood anti-inflammatory cells (Neutrophils, monocytes and lymphocytes) and TGF-β1 in swelling, stenosis, vascular destruction, thrombosis, etc. of vascular endothelial cells, releases collagen around blood vessels.
In the next stage, new blood vessels are created due to inflammation, edema, or hypoxia, which makes bleeding easier, thus activating the wound healing process, excess collagen leads to decrease vascularity, resulting in collagen deposition in the ECM [36]. In rats exposed to radiation, vascularity increased in a short time, and it was observed that collagen was not deposited in parenchymal cells but was formed around blood vessels. As shown in Fig. 5, it shows that collagen flows out of the blood vessels and is formed in the space between muscle fibers in skeletal muscles.
Although research on living body has consistently reported biological changes that occur during whole body exposure, electrical changes along with such changes have not been reported so far. The mechanism linking the electrical resistance of the whole body to changes in tissue caused by radiation exposure is complex and difficult to fully understand. Therefore, the existing method uses specific materials such as air, film, and semiconductors rather than directly measuring the living body. the radiation dose of the whole body is judged by replacing it with the ionization amount of a specific ionized material. The radiation dose of the whole body is replaced with the ionized amount of the specific ionized substance and measured. The problem with this method is that the size or component of the detector is different from that of the living body, so the exposure dose may be overestimated or underestimated.
The electrical resistance of the whole body must take into account changes in body mass index, body fat, extracellular fluid, intracellular fluid, and the like [39]. In this study, only changes in skeletal muscle were observed, and other factors that damage the whole body were not reflected in changes in electrical resistance. Compared with the control group, in the dose group with a dose of 10 Gy or higher, a significant change occurred in the selected collagen area as the dose increased, and the decrease in electrical resistance also increased. Although it was confirmed that the electrical resistance decreased in the 1 Gy group, there was no significant change in the collagen area in the selected tissue. This discrepancy means that other factors that affect electrical resistance have not been fully figured out. Also, even if there was no change in the tissue that could be found with an optical microscope, this means that changes at the molecular level or organelles within cells may affect electrical resistance. These results show that the body’s response to ionizing radiation is more sensitive to electrical signals than morphological ones.
Various parameters must be considered when analyzing the interaction between ionizing radiation and living body with electrical resistance. The reproducibility of electrical resistance may vary depending on radiation factors such as dose, dose rate, and quality, and differences in bioequivalence such as size, age, recovery capacity, and radiation sensitivity of the individual. Therefore, we expect that measurement of electrical resistance would be a useful method if the biological response to ionizing radiation could be confirmed to be independent of these factors and quantified by electrical resistance.
In this study, the relationship between histomorphological changes of specific organs and electrical resistance according to radiation dose was confirmed. It was limited to confirm changes in the cell nucleus, cytoplasm, and cell membrane using light microscopy. However, as the radiation dose increased, it was found that many changes occurred in the cell nucleus, cytoplasm, and cell membrane due to structural changes and damage to cells in the tissue. As the dose increases, the cell membrane will be severely damaged, and the cell membrane’s intrinsic capacitance will decrease. In addition, a weight loss and cell size, and depletion of parenchymal cells cause structural changes and can be factors that lower impedance. At the same time, if hydrating substances such as collagen increase and the support structure between cells becomes loose, the impedance will decrease.
In the future, if we prove the relationship between changes in electrical resistance and changes in body components such as intracellular fluid, extracellular fluid, and fat, and collect data, it will be possible to directly evaluate the radiation dose as a change in electrical resistance.
Conclusion
In this study, we investigated the electrical resistance and skeletal muscle changes in the living body due to the dose of ionizing radiation in rat. It was found that the structure in the tissue changed with the increase in radiation dose, the production of collagen increased, and the electrical resistance of the whole body decreased. This indicates that the decrease in electrical resistance due to ionizing radiation is due to histological changes and increased water content. Therefore, it was confirmed that the measurement of electrical resistance during exposure to ionizing radiation can be used as a criterion to determine the radiation dose by reflecting the dose-response relationship.
Footnotes
Acknowledgments
This study was supported by Biomedical Research Institute Grant (2018B014), Pusan National University Hospital.