
Abstract
Background:
Many viruses display affinity for polysaccharides presented at the surface of target cells with high biological relevance for virus attachment and entry. This raises the possibility of the application of polysaccharides, particularly their sulfated modifications, in studies of receptor binding and also in antiviral therapy.
Methods:
In this study, we analysed various sulfated glucans, generated from a commercial preparation of rice bran using chemical, chromatographic, spectroscopic and virological methods.
Results:
A number of sulfated polysaccharides with different charge densities were generated from a commercial rice bran preparation by aqueous extraction followed by chemical sulfation. The backbone of the type of glucans identified was made up mainly of α-(1→4)-linked glucopyranosyl residues. Sulfate groups were found to be located at C6 and partly at C2 or C3 of glucopyranosyl residues. Sulfation appeared to be very important for anti-cytomegaloviral activity, as desulfation experiments demonstrated an impairment of activity. Using an established cytomegalovirus replication assay with primary human fibroblasts, data demonstrated that the anti-cytomegaloviral effect was determined primarily at the stage of viral entry.
Conclusions:
Sulfated glucans derived from rice bran possess very promising characteristics for their potential use as entry-inhibiting anti-cytomegaloviral agents.
Introduction
Human cytomegalovirus (HCMV) is the type species of cytomegaloviruses in the β-subfamily of Herpesviridae. HCMV has the regulatory capacity to initiate a productive replication upon primary infection and to maintain a lifelong persistence, even within an entirely immunocompetent host [1]. From a latent state, HCMV can reactivate without causing symptoms in healthy individuals. In the absence of an adequate host immune response, the finely balanced host–virus interaction is shifted to an extreme situation, in which HCMV can eventually cause invasive disease and immunopathogenesis-associated effects [2]. This opportunistic pathogen is associated with significant morbidity and mortality among immunocompromised patients (in particular immunosuppressed patients after stem cell or solid organ transplantation, as well as immunocompromised AIDS or cancer patients). Additionally, congenital HCMV infection, occurring in 1–2% of all live births, is a leading cause of birth defects and pathogenic developmental failures in infants. Drugs currently available for the treatment of HCMV-associated disease include ganciclovir, its oral prodrug valganciclovir, cidofovir, foscarnet and fomivirsen. Although these drugs proved to be successful in the management of specific situations of HCMV disease, their use is often limited because of toxic side-effects, poor oral bioavailability (with the exception of valganciclovir) and the development of drug resistance. Furthermore, no drug has been licensed for use in the treatment of congenital HCMV so far; thus, there is a need to develop new compounds with strong anti-cytomegaloviral activity accompanied by favourable pharmacological and clinical properties. The search for novel inhibitors of the replication of cytomegalovirus and other herpesviruses has led to the identification of additional molecular targets, such as viral glycoproteins [3–5], and putative cellular entry receptors, such as platelet-derived growth factor receptor [6]. Herpesvirus glycoproteins can interact with both entry receptor proteins and specific sites on heparan sulfate [5,7–10]. The uniquely distributed sulfation pattern of heparan sulfate is believed to regulate its functional specificity [11,12]. Interestingly, heparin and several sulfated carbohydrates, including sulfated fucans, galactans, xylans and xylomannans, have been found to inhibit herpesvirus entry (for example, entry of herpes simplex virus) because of their structural similarity to heparan sulfate [13–15]. However, the naturally occurring sulfated polysaccharides are mostly complex and heterogeneous, which might complicate their use in medical applications. Hence, these complex macromolecules have the potential to bind to a variety of physiologically important proteins and thereby to harbour an intrinsic risk of inducing off-target effects and cytotoxicity. This risk, however, might be reduced by using structurally defined polysaccharides binding to their ligands with high specificity. In this regard, procedures of experimental sulfation are an important improvement in the generation of structurally defined polysaccharides exerting high biological activity.
Currently, huge amounts of rice bran (Oryza sativa), a byproduct of the rice processing industry, are regularly disposed of as waste, but might instead be utilized profitably as a source of various pharmaceutical products. Rice bran biomass retains heavy metals [16], has anti-oxidation activity [17] and can be used for the removal of selected organics from water [18]. The diversity of polysaccharides contained in rice bran is the basis of a number of very interesting biological activities [19,20]. However, the full exploitation of these polysaccharides in translational research and various applications requires an in-depth knowledge of their structure–activity relationship.
The present study describes the generation and chemical characterization of glucans from rice bran. Using chemical and chromatographic methods, as well as various forms of spectroscopy, we have been able to deduce structural features of this polysaccharide. The possibility to generate derivatives by chemical sulfation in the O-positions along the polysaccharide has led to the identification of an interesting series of related compounds possessing varied degrees of sulfation (DS) and charge distribution. An analysis of these compounds in cell culture-based assay systems indicated a substantial antiviral activity in a non-cytotoxic range of concentrations. In particular, a strong anti-cytomegaloviral effect could be demonstrated. The putative activity-defining components of this group of antiviral substances are discussed on the basis of our data on the structure–activity relationship.
Methods
Chemistry
Materials
The commercial rice bran preparation (Batch number G5458) was a gift from Laxmishree Rice Mill (Burdwan, West Bengal, India).
Isolation of polysaccharides from rice bran
The commercial rice bran (10 g) was extracted 3 times with water (pH 6.5–7.0) at a solute to solvent ratio of 1:100 (w/v) at 90°C for 1 h under constant stirring. Separation of the residue from the liquid extract was performed by centrifugation (Research Centrifuge R-24; Remi, Mumbai, India) followed by filtration through a G-2 glass filter (Borosil Glass Works, Ltd, Kolkata, India). The residue was briefly washed with additional distilled water and the wash was collected to maximize polysaccharide recovery. The liquid extract was dialyzed extensively against water and lyophilized (Cool Safe 55-F freeze drier; Scanvac, Lynge, Denmark). The recovered material was dissolved in water; the polysaccharides were precipitated twice with ethanol (four volumes) and then collected by centrifugation. The final pellet was dissolved in water and lyophilized to yield the water-extracted polysaccharide (RbWE; 1.2 g).
Partial acid hydrolysis
The glucan-rich pool (RbWE) was treated (0.5 mg/ml at 90°C for 2 h) with 100 mM HCl. After incubation, the resulting digest was cooled, neutralized with 2 M NaOH, dialyzed (molecular cutoff 1,000 Da; Sigma–Aldrich, Steinheim, Germany) and then lyophilized to produce rice bran glucan (RbG).
Chemical analyses
The chemicals used were of an analytical grade or the best available. All experiments were conducted at least in duplicate. Evaporations were carried out under reduced pressure at around 50°C (SB 1100 Rotary Evaporator; Eyela, Tokyo, Japan). Dialysis against distilled water was performed with continuous stirring, and toluene was added to inhibit microbial growth. Moisture was determined by drying ground material in an oven at 110°C for 3 h. Total sugars were estimated as anhydroglucose by the phenol-sulfuric-acid assay [21]. Uronic acids were assayed as anhydrogalacturonic acid using m-hydroxydiphenyl phenol colour reagent [22]. Neutral sugars were analysed after hydrolysis with 2 M sulfuric acid (3 h, 100°C), preceded by 1 h with aqueous 72% (v/v) H2SO4 at room temperature for insoluble residues. Sugars were reduced, acetylated and analysed as their alditol acetate [23] by gas–liquid chromatography (GLC; Shimadzu GC-17A; Shimadzu, Kyoto, Japan) on columns of 3% SP-2340 on Supelcoport 100–120 mesh and DB-225 (J&W Scientific, Folsom, CA, USA and by gas–liquid-chromatography–mass spectrometry (GLC-MS; Shimadzu QP 5050 A, Shimadzu). Myoinositol was used as an internal standard. Sugars in the acid hydrolysate were also analysed by thin-layer chromatography as described [24].
Sulfation and sulfate estimation
Fraction RbG was sulfated using a method previously described [25]. Briefly, the freeze-dried polysaccharide (100 mg) was soaked in 2 ml of dry N,N-dimethylformamide (DMF) and SO3-pyridine complex dissolved in 12.5 ml DMF was mixed with the polysaccharide. For every mol of SO3-pyridine complex, 1 mol of pyridine was added to the mixture. The reaction was carried out under a nitrogen atmosphere at 30°C. The pH of the solution was then adjusted to a level of 7 with aqueous 20% NaOH. Finally, sulfated polysaccharides were dialysed against distilled water and freeze-dried. The sulfated derivatives S1G, S2G, S3G and S4G were obtained when the reaction times were 4.0, 3.5, 2.5 and 2.0 h, respectively. Estimation of sulfate by the modified barium chloride method [26] and infrared (IR) spectrometry [27] was carried out as previously described [24].
Size exclusion chromatography
In system A, solutions (3–5 ml) of sulfated glucans (S1G, S2G, S3G and S4G) in 500 mM sodium acetate buffer (pH 4.0) were loaded to a Superdex™ 30 prep grade column (2.6×60 cm; Amersham Pharmacia Biotech AB, Uppsala, Sweden) equilibrated with the same buffer. The column was eluted ascendingly with the same buffer at 15 ml/h, and the temperature was 30–35°C. The elution of glucans was expressed as a function of the partition coefficient Kav (Kav=[Ve-V0]/[Vt-V0], where Vt and V0 were the total and void volume of the column determined as the elution volume of glucose and blue dextran, respectively, and Ve was the elution volume of the sample). The column was calibrated with standard dextrans within a molecular weight range of 1,000–30,000 Da. Fractions of 5 ml were collected and analysed for their total sugar contents by the phenol-sulfuric-acid method [21] using glucose as the standard.
In system B, all sulfated derivatives (S1G, S2G, S3G and S4G) were collected from the Superdex™ 30 column and injected separately on a Sephadex™ G-10 column (2.6×90 cm; Amersham Pharmacia Biotech AB). The desalted materials were concentrated and then lyophilized.
Linkage analyses
The triethylamine forms of the polysaccharides were methylated by lithium dimethylsulfinyl anion and iodomethane [28]. The permethylated polysaccharide was hydrolyzed with 2.5 M trifloro acetic acid at 120°C for 75 min, reduced with 1 M NaBD4 in 2 M NH4OH for 3 h at room temperature and acetylated using perchloric acid as a catalyst. The partially methylated alditol acetates were analysed by GLC and GLC-MS using a DB-225 column (J&W Scientific). The mass spectra were recorded with a Shimadzu QP 5050A GLC-MS instrument (Shimadzu) at 70 eV. The partially methylated alditol acetates were identified by the measurement of relative retention times, methoxyl substitution pattern as obtained from GLC-MS and carbohydrate composition of the non-methylated polymers as previously described [29].
Spectroscopy
IR spectra were recorded on a Fourier transform (FT)-IR spectrophotometer (Spectrum RX 1; PerkinElmer, Singapore) using KBr discs containing finely powdered samples. The proton nuclear magnetic resonance (1H-NMR) and 13C-NMR spectra of the RbG and experimentally sulfated glucan (S1G) were recorded on a Bruker 400 spectrometer (Bruker Biospin AG, Fallanden, Switzerland). The sulfated glucan was converted into its sodium salt by passage through a column (7 ml; Bio-Rad, Hercules, CA, USA) of Amberlite IR 120 (H+) followed by neutralization using 50 mM NaOH solution, and all samples were deuterium-exchanged by lyophilization with D2O (Cambridge Isotope, Ltd, Andover, MA, USA) and then examined as 1% solutions in D2O (99.96 atom % D).
Virology
Cell culture and viruses
Primary human foreskin fibroblasts were cultivated in Eagle's minimum essential medium containing 7.5% (v/v) fetal calf serum. HCMV (strains AD169-GFP and TB40 UL32-EGFP) [30,31] and varicella zoster virus (VZV; vaccine strain Oka) were propagated in human foreskin fibroblasts and virus replication was quantified by automated green fluorescence protein (GFP) fluorometry or plaque reduction assay, respectively [30,32]. Epstein–Barr virus (EBV; strain B95-8) was used for infection of 293T cells (cultivated in Dulbecco's modified Eagle's medium containing 10% fetal calf serum) and virus replication was quantified by applying a reporter-based replication assay [33].
Results and discussion
Composition of the rice bran and its water-extracted polysaccharide
As a major objective, low molecular weight sulfated polysaccharides should be generated from rice bran and characterized in order to study their anti-cytomegaloviral activity. Therefore, the first step was the determination of the monosaccharide composition of the commercial rice bran preparation and, on the basis of the information obtained, to devise strategies for the extraction of polysaccharides. The bran used contained 61% (w/w) sulfuric acid (2 M) hydrolysable polysaccharides (Table 1) of which a minor component (0.6–0.8%) was uronic acid. The main neutral saccharide was glucose (Table 1), but xylose and arabinose were also released. The RbWE amounted to 12% of the starting bran preparation and contained 69% polysaccharides on the basis of the fraction dry weight. It was composed mainly of glucose (Table 1). Xylose and arabinose were the two other monosaccharides found in small quantities.
Sugar composition of fractions generated from rice bran
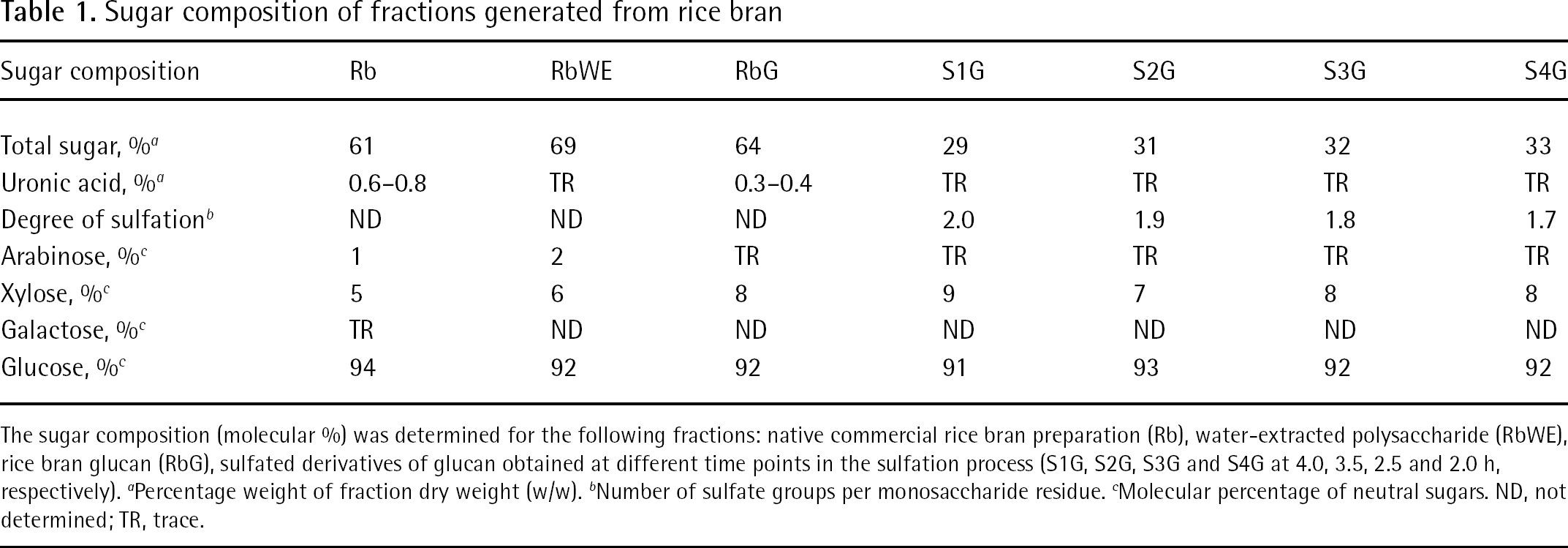
The sugar composition (molecular %) was determined for the following fractions: native commercial rice bran preparation (Rb), water-extracted polysaccharide (RbWE), rice bran glucan (RbG), sulfated derivatives of glucan obtained at different time points in the sulfation process (S1G, S2G, S3G and S4G at 4.0, 3.5, 2.5 and 2.0 h, respectively).
Percentage weight of fraction dry weight (w/w).
Number of sulfate groups per monosaccharide residue.
Molecular percentage of neutral sugars. ND, not determined; TR, trace.
Partial acid hydrolysis of rice bran glucan, sulfation and glycosidic linkage analyses
One poorly exploited potential of polysaccharides is their frequently reported ability to interfere with both very early and very late steps in viral replication, such as entry and release [14]. As far as the inhibition of viral release and spread is concerned, small-sized oligosaccharides were shown to affect cell-to-cell spread of viruses more efficiently than their high molecular mass counterparts [34]. Therefore, we have decided to generate polysaccharides with comparably low molecular mass from the RbWE. Based on the results of pilot experiments, we hydrolyzed the RbWE fraction with 100 mM of HCl to yield a fraction termed RbG. The total polysaccharide content of the RbG fraction was 64% on the basis of the fraction dry weight (w/w) and the major sugar was glucose (Table 1).
As a general rule, the DS has a major effect on the antiviral activity of polysaccharides [15]; therefore, a high charge density appears to be favourable for high antiviral activity, at least for particular classes of polysaccharides [14]. In addition to this well-documented effect of DS, the specific position of the sulfate group has also been reported to be important for antiviral activity of the various sulfated glycans [14,35]. To investigate the specific effect of DS, we sulfated RbG under various conditions to yield sulfated derivatives S1G, S2G, S3G and S4G. These sulfated glucans were purified by size elution chromatography on a Superdex™ 30 prep grade column. Fractions eluted at Kav values 0.3–0.6 were pooled and lyophilized. The polysaccharide content and their specific DS are given in Table 1. IR spectra of the RbG and its O-sulfated derivatives S1G and S4G (Figure 1) strongly suggest the conversion of hydroxyl groups to O-sulfate groups. The intensity of the absorbances at 1,260 cm−1 and 800–850 cm−1 attributed to the stretching of the S=O bond and C–O–S bonds, respectively, are dramatically increased by O-sulfation. Similarly, the intensity of the bands at 2,932, 1,424, 1,370 and 1,004 cm−1, attributed to the stretching and or deformation vibration of C–O–H bonds, are decreased in the spectrum of the O-sulfated glucan sulfate. Assignments of IR absorption bands at 1,260 cm−1 and in the 800–850 cm−1 spectral region were based on the principle originally published by Orr [36] and by Lloyd and Dodgson [37]. Bands at approximately 818 cm−1 were tentatively ascribed to 6-O-sulfate groups based on the report of Grant et al. [38], and the band at 828 cm−1 was ascribed to C–O–S stretching of equatorial 2-O and 3-O-sulfate groups of glucose residues based on reports by Lloyd and Dodgson [37] and Lloyd et al. [39].
The 1H-NMR spectrum of the RbG shows the anomeric signal (H1) at 5.42 ppm and ring protons (H2–H6) at 3.6–4.0 ppm, characteristic of α–glucan (Figure 2A). The 13C-NMR spectrum of a sulfated glucan fraction (S4G) was measured (Figure 2B), and signals were assigned by comparison with data published for maltose, amylose, amylopectin and a sulfated glucan from alga [40,41]. The C6 signal of α-(1→4)-linked glucopyranosyl residue appears around 60–61 ppm [42]. Besides this, it has been reported that O-sulfation of α,β-D-glucose causes a strong, downfield shift (+6.5 to +9.5 ppm) of the substituted resonances that are accompanied by smaller, up- or down-field displacement (−2.2 to +0.2 ppm) of the signals of adjacent carbon atoms [43]. The presence of signals around 67–68 ppm and absence of signals at 61–62 ppm suggests that all hydroxyl groups at the position C6 of the glucans analysed in the present study were sulfated. This was the expected situation because the hydroxyl group at the C6 position is primary, whereas those at C2 and C3 are secondary. The chemical shift of C6 is in good agreement with that of algae-derived sulfated glucans (67.6 ppm). In maltose, amylose, amylopectin and algal glucans, the signal of C2 is usually downfield from the C3 signal, which in turn is downfield from C5 [41]. Therefore, signals around 76, 74 and 73 ppm in our analysis could tentatively be attributed to the C2, C3 and C5, respectively, of monosulfated glucose residues that are O-sulfated at C6. Another striking characteristic in the chemical shifts of values are the downfield shifts in C2 and C3 that might indicate their participation in O-sulfation. Taken together, signals at 82.8 and 81.4 ppm could tentatively be assigned to the C2 and C3 of the 2,6- and 3,6-disulfated residues. Comparison of the intensities of these signals with C2 and C3 signals of mono-O-sulfated residues suggests that almost one-half of the glucose residues were not O-sulfated at C2 and C3 positions. The chemical shift values (79–80 ppm) of the signals of C4 indicate its participation in the glycosidic linkage (a similar shift to 79.9 ppm in the sulfated glucans derived from algae has been previously demonstrated [41]).
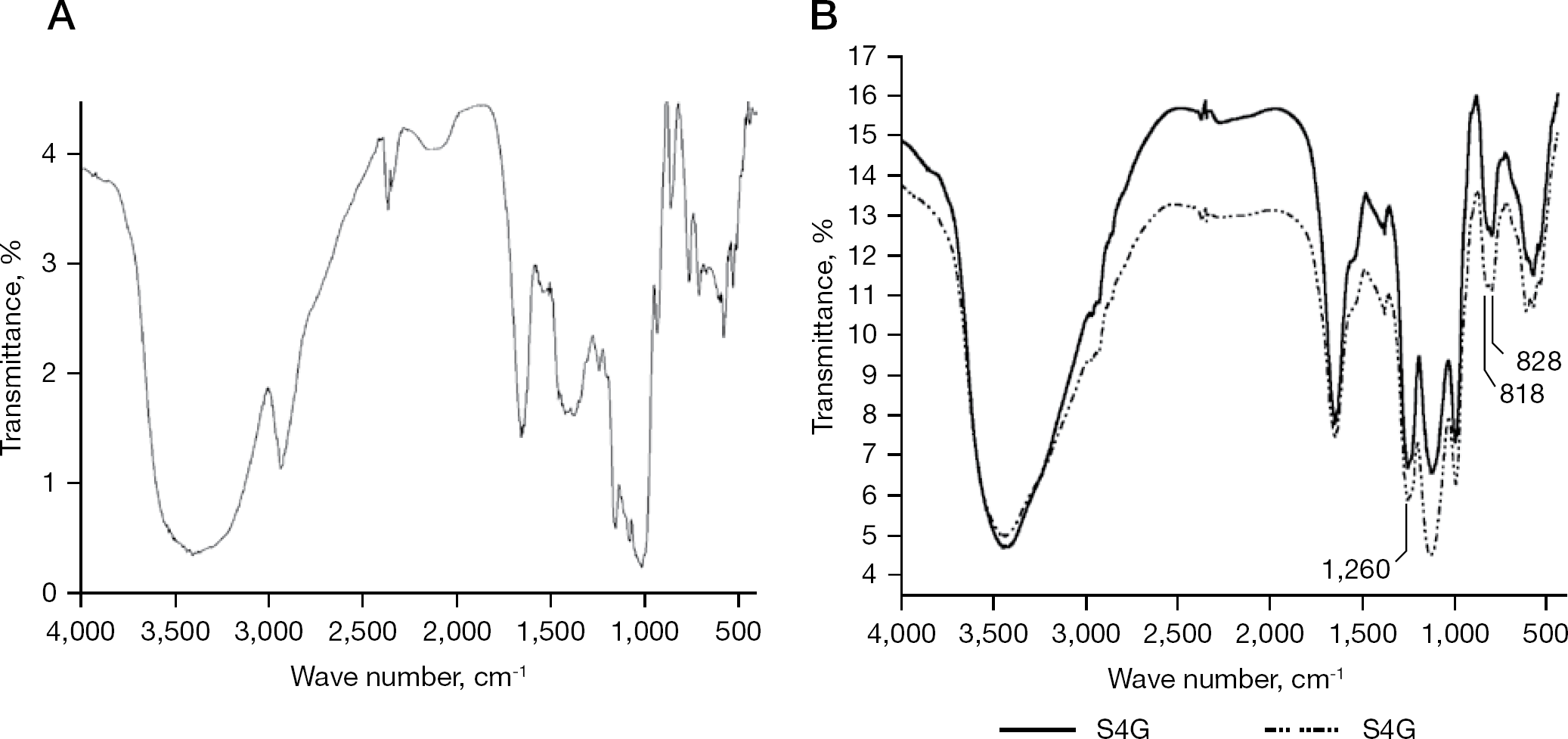
Fourier transform-infrared spectra of RbG and its sulfated derivatives
Glycosyl linkage analysis of the RbG indicated the presence 1,4,5-tri-O-acetyl-2,3,6-tri-O-methylglucitol residue suggesting that the polymer is linear and possesses a (1,4)-linked backbone (Table 2). Analysis of S4G showed five different partially methylated alditol acetates indicating the presence of sulfate groups at various positions. The appearance of 1,4,6-linked residues suggests the presence of 1,4-linked glucopyranosyl units that are O-sulfated only at C6. Similarly, the presence of 1,2,4,6- and 1,3,4,6-linked glucopyranosyl residues suggests the presence of 2,6- and 3,6-disulfated residues, respectively. Another striking feature of this analysis is the presence of 1,4-linked residues. This might indicate that some of the glucopyranosyl residues are non-sulfated. In the present study, methylation analysis was carried out under strongly basic conditions, which might have induced partial desulfation (particularly at C6 positions), possibly followed by a conversion of the hydroxyl groups into their methyl ethers.
Partially methylated alditol acetates derived from RbG and its sulfated derivatives S1G and S4G
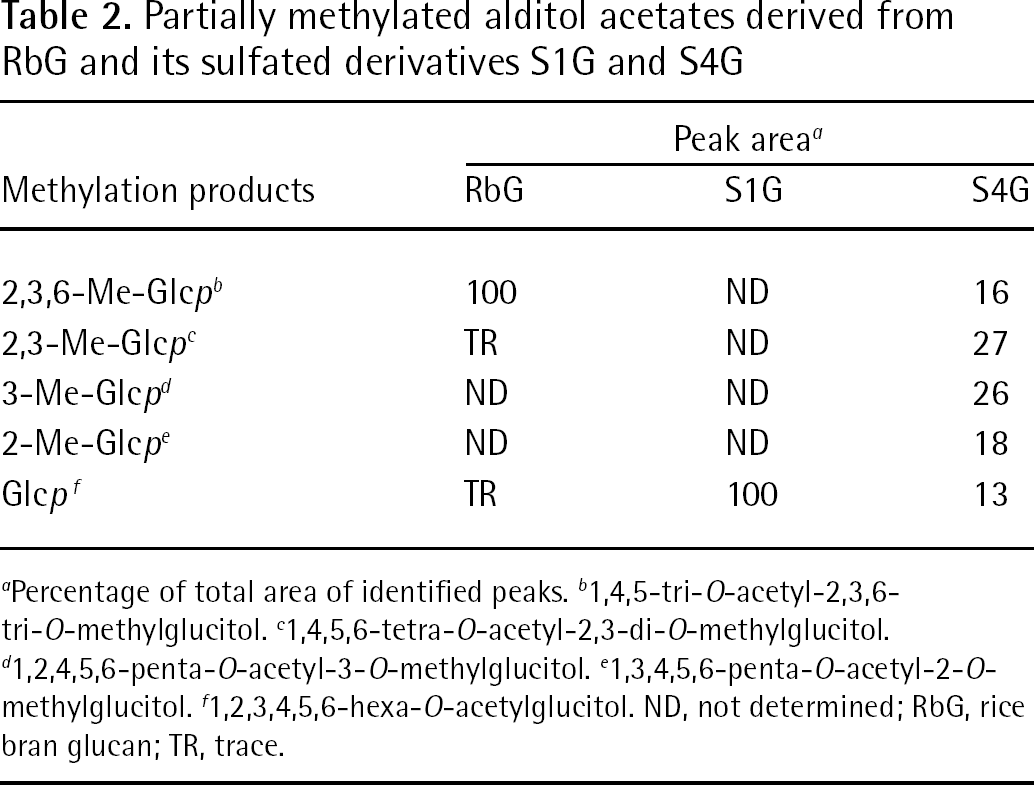
Percentage of total area of identified peaks.
1,4,5-tri-O-acetyl-2,3,6-tri-O-methylglucitol.
1,4,5,6-tetra-O-acetyl-2,3-di-O-methylglucitol.
1,2,4,5,6-penta-O-acetyl-3-O-methylglucitol.
1,3,4,5,6-penta-O-acetyl-2-O-methylglucitol.
1,2,3,4,5,6-hexa-O-acetylglucitol. ND, not determined; RbG, rice bran glucan; TR, trace.
Appearance of 1,2,3,4,6-linked glucopyranosyl residues suggests the presence of 2,3,6-trisulfated residue, but complete methylation of sulfated polysaccharides is generally difficult as the steric hindrance and electronic effect exhibited by the sulfate esters represents a limitation [44–46]. Finally, analysis of S1G revealed a single peak corresponding to glucitol hexa-acetate, which indicates that this compound might have been completely sulfated (Table 2).
Anti-cytomegaloviral activity of rice-bran-derived sulfated polysaccharides
The preparations of sulfated polysaccharides were analysed for antiviral activity. For this purpose, an established cell culture-based assay for quantifying the replication of HCMV was applied, and proved to be highly reliable for the screening of novel antiviral substances [30–33,47]. Amongst all substances analysed, S1G and S2G exerted a particularly strong and concentration-dependent inhibitory activity towards HCMV replication (Figure 3A and 3B). No sign of cytotoxicity was observed for these substances in the analysed range of concentrations (determined by microscopic inspection of cells and testing of cytotoxicity using the CytoTox 96 Non-Radioactive Cytotoxicity Assay; Promega, Mannheim, Germany; TG et al. data not shown). The lowest 50% inhibitory concentration (IC50) of anti-HCMV activity was determined for S1G as a mean ±SD of 3.46 ±0.63 μg/ml. Importantly, these hypersulfated glucans (S1G and S2G) showed clear anti-cytomegaloviral potential, whereas the non-sulfated parental substance (RbG) was negative (Figure 4A). Thus, these data support the previously stated notion that the DS is one of the determinants conferring high antiviral activity to polysaccharides.
Viral entry is the main mode of activity of sulfated glucans S1G and S2G
As a characteristic feature of the anti-cytomegaloviral activity of rice-bran-derived sulfated glucans S1G and S2G, virus inhibition was primarily achieved when the substances were preincubated on the cells (added 1 h prior to HCMV infection) and/or were present during the virus adsorption phase. When these substances were not present during this initial period of the experiment, but added at later time points (for example, directly after the virus adsorption phase), very little or no inhibitory effect was measured (Figure 3B; central bars). In particular, the preincubation of S1G and S2G alone was sufficient to produce a substantial antiviral effect (Figure 3B; left-side bars). The most pronounced antiviral activity of these polysaccharide substances, however, was measured when they were used for preincubation and remained continuously present during virus replication until the termination of the experiment (Figure 3B; right-side bars). This feature pointed to an inhibition of HCMV entry and penetration, and stood in contrast to the well-characterized inhibitory effect produced by the reference drug, ganciclovir (GCV), an antagonist of viral genome replication. Although GCV showed no inhibitory potential when exclusively preincubated on cells and removed before the addition of virus inoculum, later time points of addition were highly effective. This finding demonstrated a marked mechanistic difference between GCV and the sulfated glucans S1G and S2G. To address the question of whether the measured antiviral activity might have resulted from a direct virucidal effect, S1G was preincubated with inoculum virus before infection. After 1 h, infectious virus particles were separated from the substance by high-speed centrifugation (3 h at 14.000 rpm in an Eppendorf centrifuge 5415 R; Hamburg, Germany) and subsequently used for the infection of cells. Under these conditions, no significant inhibitory effect could be determined up to the highest concentration analysed (270 μg/ml), indicating that virucidal activity did not play a major role in the anti-cytomegaloviral potency of S1G (Figure 3C). Moreover, an experiment was performed to analyse the possible inhibitory activity of S1G on viral cell-to-cell spread. For this purpose, virus infection (HCMV strain AD169-GFP) was performed in the presence of a neutralizing human immunoglobulin preparation (Biotest HIg 6030398; kindly provided by Michael Mach and Barbara Kropff, University of Erlangen–Nuremburg, Erlangen, Germany) or a non-neutralizing control serum (rabbit), which were both added to the culture media after viral entry. At 7 days post-infection, viral replication was quantified by measuring the GFP signals expressed (by GFP microscopy and fluorometry). When S1G had been added to the cells after viral entry (post conditions), inhibition of HCMV (strain AD169) was very inefficient, both under suppressed levels of viral spread by neutralizing HIg, as well as in the control without neutralizing HIg. Therefore, no reduction in viral plaque size was noted, indicating that effects on cell-to-cell spread were poor or missing (MM et al., data not shown). When S1G was incubated throughout the infection (pre and post conditions), inhibition was very efficient in both cases, thus confirming the earlier statement that the main mode of anti-cytomegaloviral activity was directed to viral entry.
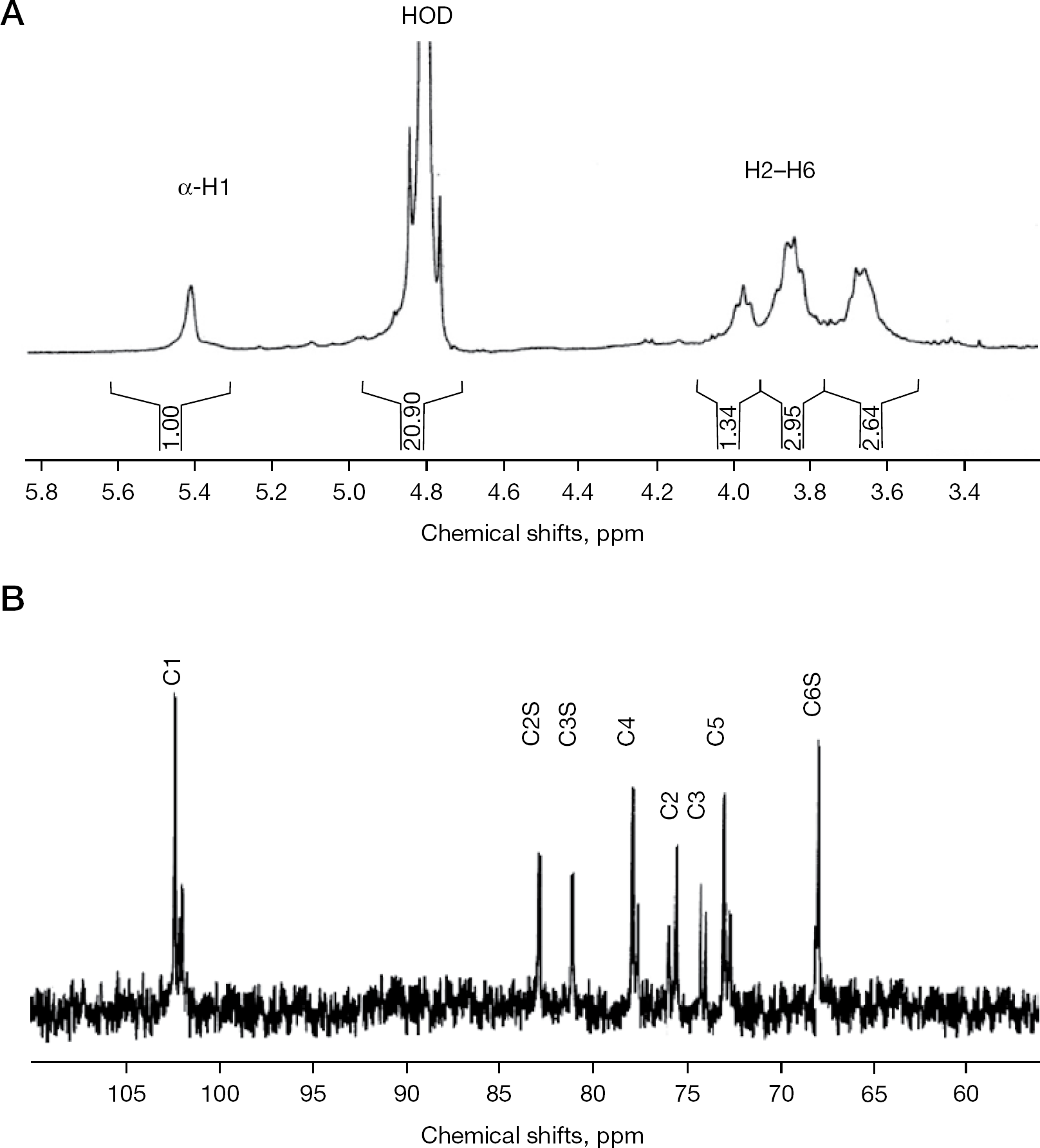
Spectra of RbG and S1G
Antiviral activity of S1G is selectively detectable for human cytomegalovirus but not for other human herpesviruses
The question of whether S1G and possibly related rice-bran-derived sulfated polysaccharides exert an antiviral activity selective or non-selective for HCMV (that is, whether S1G produces a similar inhibitory effect towards other human herpesviruses) was addressed. In order to do so, a comparative analysis was performed using HCMV, VZV and EBV. Interestingly, S1G selectively inhibited HCMV but was entirely ineffective against VZV and EBV (Figure 4). The anti-HCMV activity was independent from viral strain, as very similar inhibitory effects were determined for strains AD169 and TB40 (Figure 4A and 4B; GFP-expressing recombinants of the two viral strains were used to quantify viral replication). Notably, in the range between 1.1 and 90 μg/ml, a very pronounced selectivity of this sulfated glucan for HCMV could be demonstrated. By comparison, the non-hypersulfated parental preparation RbG was negative for all four viruses analysed, thereby highlighting the significance of DS of these substances for antiviral activity. The mode of anti-HCMV activity of S1G and the molecular basis of selectivity will have to be determined to learn more about this highly interesting group of rice-bran-derived substances.
In summary, the findings of this study highlight several novel and important aspects of the commercial rice bran preparation-derived polysaccharides with regard to their antiviral properties: a number of different soluble polysaccharides could be easily generated from commercially available rice bran preparation with high yield; these substances exerted high biological activity that could be analysed in cell culture-based assay systems at a low level of cytotoxicity (undetectable up to the concentration of 270 μg/ml); a strong antiviral effect was demonstrated for two strains of HCMV (mean ±SD IC50 of S1G was 3.46 ±0.63 μg/ml for HCMV AD169-GFP); the DS appeared to be an important hallmark of anti-HCMV activity; the main mode of action was directed to HCMV entry; and amongst four human herpesviruses investigated, the antiviral activity of S1G was selective for HCMV. Further studies will be needed to characterize the mode of antiviral activity in detail and to define the potential of sulfated polysaccharides for their putative usefulness for topical administration in antiviral therapy [14].
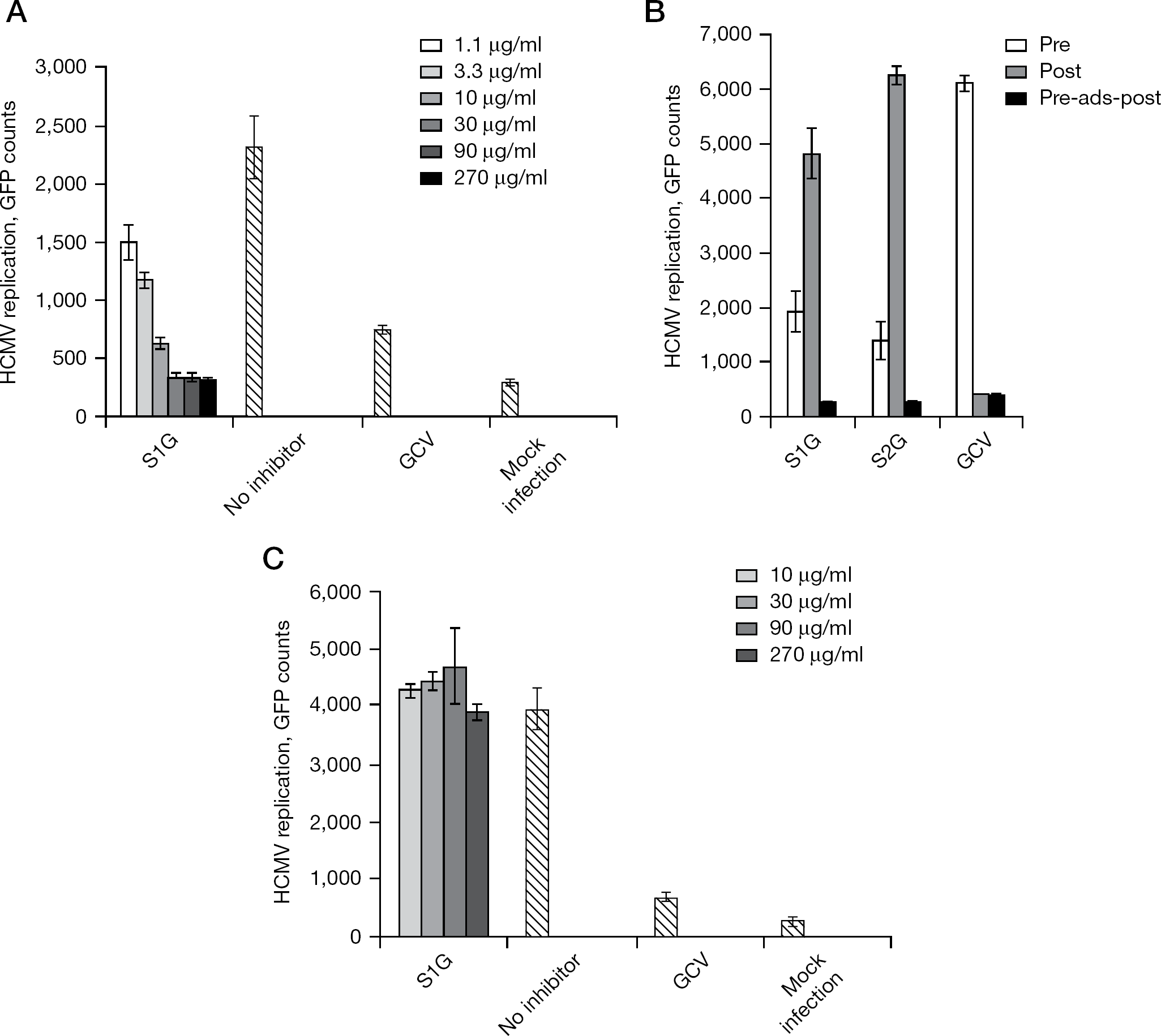
Inhibition of HCMV replication by sulfated glucans
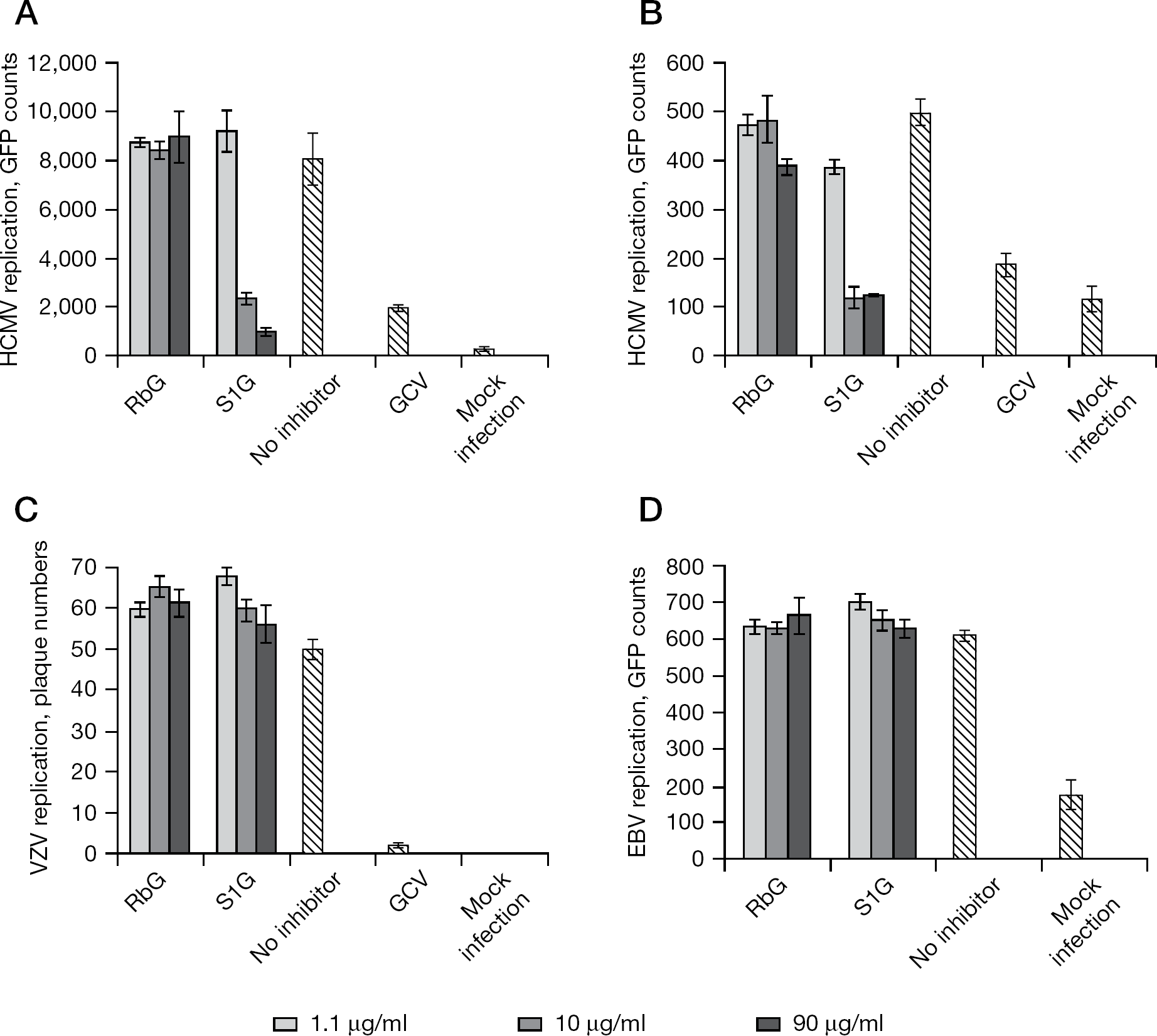
Comparative analysis of the replication of herpesviruses: potential inhibitory effects of experimentally hypersulfated glucan versus poorly sulfated glucan
Footnotes
Acknowledgements
The helpful scientific and methodical collaboration of Sabine Rechter, Jens Milbradt and Rike Webel is gratefully appreciated. Standard dextrans were a gift from Tapani Vuorinen (Helsinki University of Technology, Helsinki, Finland). TG and SS thank the Council for Scientific and Industrial Research for their fellowships.
This work was supported by grants from the Department of Science and Technology, New Delhi, India (project number SR/S1/OC-50/2007 to BR), ARGUS Foundation, Berlin, Germany (to Rike Webel and MM) and Deutsche Forschungsgemeinschaft (SFB 796/C3 and MA 1289/7-1 to Jens Milbradt, SA and MM). The authors declare no competing interests.