
Abstract
This short note highlights a procedure to distinguish the residues having similar masses, oxidized methionine and phenylalanine containing peptides using MALDI TOF/TOF. The isotope intensities give a preliminary recognition of peptides containing oxidized methionine. In the peptides with partial oxidation of methionine a mass difference of 16 Da can be observed in the mass finger print of the peptide. Neutral loss of methane sulphenate (CH3 SOH) in the MS/MS spectra is the most abundant ion in the peptide containing oxidized methionine, whereas this fragment ion is not produced from phenylalanine containing peptide. The mass spectra of methionine, oxidized methionine and phenylalanine containing peptides were examined from the proteins of Pseudomonas syringae Lz4W, whose genome sequence is not known.
Introduction
In Proteomics, proteins are identified using mass spectra of the trypsin-digested peptides called peptide mass figure print (PMF), and subsequently obtaining the sequence of these peptides using tandem mass spectra. Identification of peptide sequences by mass spectra involves the use of several algorithms such as MASCOT and SEQUEST. These programs also give an estimate of the false positive rates of identifying the peptide sequences. Proper identification of peptide sequence is essential for the identification of proteins, particularly when the genome sequence of the species under study is not available. De novo sequencing is also a preferred option to obtain the peptide sequence from the mass spectra. 1 However, it cannot be routinely used because of several limitations associated with the fragmentation of peptides such as the chemical nature of the peptides, the type of mass instrument used and the method of ionization. It is important to look for other methods in order to avoid false positives in identification. Methionine is a sulfur containing amino acid with an approximate natural abundance of 2.2% in proteins. 2 Most of the proteins occurring from biological sources contain methionine. This amino acid besides cysteine, tryptophan, tyrosine and histidines, is susceptible to oxidation. 3 The oxidation of methionine to sulfoxide form is of particular interest as it has been found to occur in a wide variety of proteins. The consequences of mehtionine oxidation has been studied in biological process and reviewed from time to time.4,5 Most often it reduces or eliminates biological activity of the protein.6,7 Methionine oxidation also brings about changes in the structure of the proteins.8,9 It was shown that methionine sulfoxide reductase present in the cells reduced oxidized methionine to methionine and protect dopaminergic cells from Parkinson's related complications. 10 These studies signify the importance to identify oxidation of methionine in proteins.
The origin and control of some of the oxidation reactions of methionine, cysteine and tryptophan residues in proteins are known. 11 Partial oxidation of methionine is a common phenomenon during some of the processes used in proteomics. The protein spots on the SDS-gel most often contain molecular species having oxidized methionine and methionine. Both of these molecular species appear at the same position and therefore can't be distinguished. The tryptic peptides containing methionine and the peptides containing oxidized methionine show two different signals in the PMF with a mass difference of 16 Da. Identification of oxidized methionine using FAB—MS in peptides was reported earlier. 12 However, the oxidized methionine and phenylalanine have similar mass of 166.0532 and 166.0862 respectively. Hence it is difficult to distinguish the peptides containing these residues. Therefore it is necessary to develop a methodology to identify oxidized methionine containing peptides. Mass spectrometry is extensively used to identify the various oxidation reactions in proteins and peptides. 13 Several factors of mass spectra are useful. The distribution of the isotope peaks of the parent ion serves as signatures of sulfur-containing amino acids. The y-series and b-series of ions can be used for determination of the sequence of peptides. In addition to these, neutral loss of methane sulfenate in these spectra confirms oxidation of methionine (Fig. 1).
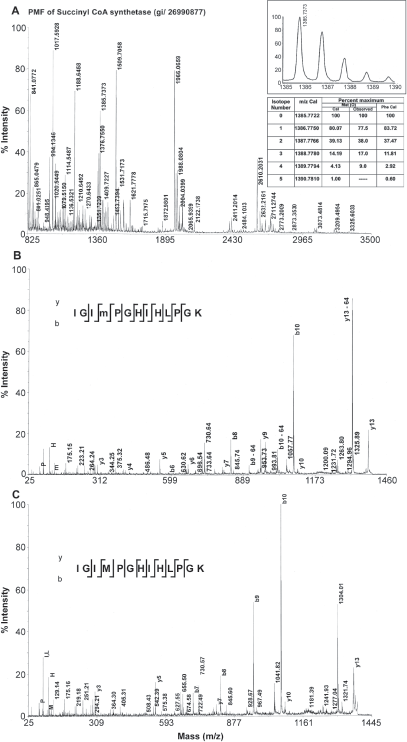
PMF of a protein from P. syringae obtained by tryptic digest from a 2D gel spot (Panel A). The inset shows the comparison of intensities of the isotope peaks of oxidized methionine (m) and phenylalanine containing peptides. Panel B and C exhibit the MS/MS spectra of oxidized methionine and methionine containing peptides respectively from the protein shown in Panel A.
Our laboratory is involved in identification of proteins from cold-tolerant bacterium Pseudomonas syringae Lz4W, isolated from soil samples in around a lake of Antarctica. 14 In the present investigation, the membrane proteins of this bacterium were separated on 2D SDS-gels (unpublished results). The peptides were extracted and analyzed using MALDI TOF/TOF. Several phenylalanine and oxidized methionine containing peptides were analyzed from the trypsin-digested proteins.
Materials and Methods
The isotope abundances for various peptides can be calculated using the web site http://prospector.ucsf.edu. MS-Isotope utility program was used from this web site to calculate the isotope abundances of peptides. The experimental intensities of the isotope peaks and mass values can be obtained from the spectra. The genome sequence of Pseudomonas syringae Lz4W being not known, the genome sequences of the other Pseudomonas sp. available at NCBI (updated up to July 2007) was used to identify the proteins as described earlier 15 using MASCOT search tools.
Results
Phenylalanine and oxidized methionine have same molecular weight, but they differ in the isotope distribution. The natural abundance of sulfur isotopes is 32S-95.02%, 33S-0.75%, 34S-4.21%, 36S-0.02%. 16 The isotope distribution of methionine containing peptides in general showed a higher intensity of isotopic peaks due to increased natural abundance of 34S from second isotope peak (Fig. 1A inset). This is suggestive of a preliminary indication to distinguish between oxidized methionine containing peptide and phenylalanine containing peptide. A decrease in relative intensity of the first isotope peak and an increase in the relative intensity of third isotope of a tryptic peptide obtained from succinyl CoA synthetase alpha sub unit were provided as an example (Fig. 1A). The mass and the intensities of different isotopes were calculated by replacing oxidized methionine with phenylalanine. The oxidized methionine residues could be differentiated from phenylalanine residues by the higher intensity of the second isotope (34S-4.21%) associated with the former. Table 1 shows some tryptic peptides containing methionine, oxidized methionine and phenylalanine peptides from a few identified proteins using database search. The peptide mass finger print of a protein succinyl CoA synthetase that had produced peptides with partial oxidation of methionine was shown in Figure 1. Both oxidized methionine-containing peptide and methionine-containing peptide appeared in the mass spectrum of the tryptic peptide with a mass difference of 16 Da (Fig. 1 A, Table 1). Loss of 64 Da from precursor ion has been found to be the most abundant ion in the MS/MS spectrum (Fig. 1B). The peaks help to distinguish oxidized methionine form phenylalanine are shown in the Table 1. Manual verification of the MS/MS spectra clearly identified the oxidized methionine ruling out the possibility of the residue being a phenylalanine.
Analysis of some tryptic peptides form the proteins of Pseudomonas syringae containing methionine, oxidized methionine (M*) and phenylalanine.
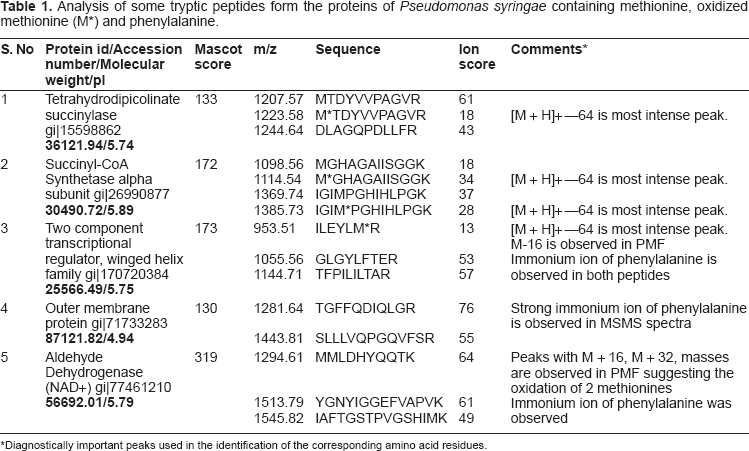
Diagnostically important peaks used in the identification of the corresponding amino acid residues.
Discussion
Tryptic digests of several peptides obtained from the proteins of an Antarctic bacterium P. Syringae, containing methionine, oxidized methionine and phenylalanine residues were analyzed. Oxidation of methionine was used as a marker for the identification and confirmation for the presence of methionine residue in the peptide sequences as is shown in this study. This analysis has helped identify the presence of methionine residues, and absence of phenylalanine residue in the peptide. The crucial information to distinguish oxidized methionine and phenylalanine has been obtained from the MS/MS spectra of the peptides. The evidence for the presence of oxidized methionine in the peptide was further obtained by the appearance of a most abundant ion, which was formed by the neutral loss of methane sulfenate, as indicated by a loss of 64 Da from the precursor ion in the CID mass spectra. The algorithms used for de novo sequence does not consider the intensity of the fragment ions for sequence determination. A recent review highlighted the influence of the abundant ions produced in tandem mass spectrometry experiment and statistical methods to discover these factors. 17 These studies highlight the importance of the abundance of the ions produced in tandem mass spectra and their significance in the determination of the sequence of peptides. In addition, the immonium ions of oxidized methionine and phenylalanine appear at m/z 147.03 and 147.06 respectively. Methionine sulfoxide undergoes immonium ion formation, although this ion is of low abundance in the methionine sulfoxide spectrum. This is due to the instability of methionine sulfoxide immonium ion and this leads to easy elimination of CH3SOH. Direct loss of complete side chain from protonated molecule of methionine sulfoxide [M + H-91] seems to result in a stable ion during FAB MS. 12 The spectra obtained using MALDI TOF/TOF also produced this ion.
In conclusion, using methionine oxidation as a chemical marker, the sequence of methionine containing peptides has been determined. This has helped in the identification of peptide sequences containing these residues from an Antarctic bacterium. Manual verifications the MS/MS spectra are essential for correct identification.
Disclosure
The author reports no conflicts of interest.
Footnotes
Acknowledgements
I acknowledge the financial support from the department of Biotechnology, New Delhi through a research grant (BT/PR7383/BRB/10/474/2006). I sincerely thank Dr. M. K. Chattopadhyay for his help in the preparation of this short note.