
Abstract
Transient receptor potential (TRP) channels have been found to play important roles in cellular physiology and hold promise as therapeutic targets. These channels activate in response to a variety of chemical or physical stimuli and conduct non-selective cation currents (NSCC). Due to their unique activation properties, application of automated electrophysiology to measure the channel activity has been difficult. Using HEK293 cells stably expressing human TRP channels, hTRPC6 and hTRPA1, we developed and validated a high-throughput Rb+ efflux assay for NSCC channels. The assay was performed in cell-based 96-well format. A significant increase in Rb+ efflux can be detected upon channel activation by specific agonists, confirming that both TRPC6 and TRPA1 channels are permeable to Rb+ ions. The agonists induced Rb+ efflux can be blocked by known channel blockers and selected compounds from our high-throughput screening (HTS) hits. The assay is suitable for HTS with Z′ factors of 0.53 and above. We also tested the Ca2+ effect on channel activities in this assay. Both TRPC6 and TRPA1 channels were found to be inhibited by increasing the concentration of Ca2+ in the assay buffer. However, Ca2+ significantly reduced the potency of allyl isothiocyanate (AITC) on TRPA1 but did not affect the potency of carbochol on TRPC6. Using this assay for secondary confirmation screen, we successfully identified and confirmed the positive hits as TRPC6 inhibitors.
Introduction
Despite the significant improvement made in ion channel assay technologies, it still lacks robust high-throughput assay for TRP channels, especially for TRPC6 channel. Currently available high-throughput assay is mostly fluorescence-based using Fluorometric Image Plate Reader platform (FLIPR). Although this assay is applicable for some of the TRP channels, the fluorescence labeling-related issue inevitably generates higher nonspecific hit rate resulting from the interaction with autofluorescent or quenching compounds. To address this issue, it is necessary to have a non-fluorescence-based assay for either primary or follow-up confirmation screen. The automated electrophysiology platforms such as IonWorks and Q-Patch have quickly evolved in recent years and have been successfully applied to screening of some ion channels including both voltage- and ligand-gated classes. 14,15 However, due to the unique activation properties of TRP channels, this technology has not been successfully applied for these channels. Searching for a high-throughput, robust, and cost-efficient assay for screening on TRP channels is absolutely necessary for identification of novel compounds against these potential drug targets.
Rubidium is a metal element with the physicochemical properties very similar to potassium. Rb+ is often used as a tracer ion to assess potassium channel activities. The radioactive 86Rb+ was broadly used for measuring K+ channel function and/or K+ transportation across biological membranes. 16,17 In recent years, a nonradioactive label-free technology for Rb+ measurement, flame atomic absorption spectrometry has been developed in automated format and successfully applied to the high-throughput assays for K+ channel targets. 18 –20 The assays have been demonstrated sensitive, robust, and cost efficient. However, it is still unknown whether the assay can be utilized for NSCC channels. Using stable cell lines expressing TRP channels, we have demonstrated in the present study that Rb+ efflux assay can be a robust high-throughput screening (HTS)-suitable assay format for functional assessment of NSCC channel activities. The assay has been successfully applied to our efforts in searching TRP channel modulators.
Materials and Methods
Media and Chemicals
Cell culture media
For HEK293-hTRPC6, Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine serum (FBS), 1% penicillin/streptomycin, and 150 μg/mL hygromycin (Invitrogen, Carlsbad, CA) was used in cell culture. For HEK293-hTRPA1, DMEM/glutamax high glucose with 10% dialyzed FBS, 1% penicillin/streptomycin, 1% sodium pyruvate, 1% MEM NEAA, and 400 μg/mL hygromycin was used.
Patch clamp pipette solution
The patch clamp pipette solution contains (in mM) 140 CsCl, 4 MgCl2, 10 EGTA, and 10 HEPES (pH 7.2).
Patch clamp external solution
The patch clamp external solution contains (in mM) 135 NaCl, 2 KCl, 1 CaCl2, 1.17 MgSO4, and 10 HEPES (pH 7.4). For testing Rb+ permeability of the channel, NaCl was replaced by RbCl. For testing cation permeability, NaCl was replaced by N-methyl-
Rb+ efflux assay buffer
For Rb+ efflux assay, the assay buffer contains (in mM) 140 NaCl, 0.15 CaCl2, 3.3 KH2PO4, 0.8 K2HPO4, 1.2 MgCl2, 10
Rb+ efflux lysis buffer
1% Triton® X-100 solution was used as lysis buffer. HC030031 was purchased from TOCRIS Bioscience (Ellisville, MO). N-(p-amylcinnamoyl)anthranilic acid (ACA) was purchased from EMD Chemicals Inc. (Gibbstown, NJ). All other chemicals and compounds were purchased from Sigma-Aldrich (St. Louis, MO). Compounds were prepared in dimethyl sulfoxide (DMSO) for stock and freshly diluted into the buffer with DMSO concentrations below 0.2%.
Stable Cell Line Generation
TRPC6 stable cell line was generated using HEK293 cells transfected with the human TRPC6 channel expression construct. hTRPC6 full-length DNA (accession number: BC093660; Thermo Fisher Scientific, Huntsville, AL) was subcloned into pCDNA-hygro vector (Invitrogen, Carlsbad, CA). After sequencing confirmation, the construct DNA was amplified and used to transfect HEK293 cells. HEK293 cells were seeded into a 10-cm culture dish at a density of 3.5–4 × 106 cells. After overnight incubation, cells were transfected with DNA construct using Lipofectamine™ 2000 (Invitrogen, Carlsbad, CA) according to the manufacturer's protocol. Five hours later, transfection medium was replaced and the cells were incubated overnight at 37°C in 95/5% air/CO2. The transfected cells were then harvested and replated into a 96-well cell culture plate at a density of 2–5 cells/well in the medium containing 150 μg/mL hygromycin. After 48 h of incubation, colonies were observed. The cells were then individually picked and scaled up for functional determination of channel expression using FLIPR membrane potential assay. TRPA1 stable cells were obtained from our colleagues in Department of Neuroscience, Wyeth Research.
Patch Clamp Recording
Cells were dissociated using trypsin (0.05 mg/mL) and harvested into 15-mL centrifuge tube. The freshly isolated cells were used within 4 h after preparation. High-resolution whole-cell current recordings were acquired with a computer-based patch clamp amplifier system (EPC-10; HEKA, Lambrecht, Germany). Patch pipettes had resistance between 2 and 5 megohm after filling with the standard pipette solution. After establishing whole-cell configuration, the membrane potential was held at −20 mV. A 1-s ramp voltage protocol from −100 to 100 mV for TRPC6 or from −60 to 60 mV for TRPA1 was applied repetitively to evoke channel currents in an interval of 5 s. Liquid junction potential, capacitive currents, and series resistance were determined and corrected using the automatic compensation of EPC-10. Compounds were delivered through a gravity-driven perfusion system. The current amplitudes at ±60 mV were used for determining compound effect.
Rb+ Efflux Assay
For Rb+ efflux assay, cells were plated in 96-well poly-
Percent cellular Rb+ content = Rb+ content in cell lysis/total Rb+ content × 100
Starting from Rb+ loading process, all experiments were performed at room temperature (22°C ± 1°C).
Data Analysis
The Rb+ loading curves were fitted to single exponential growth equation as follows:
where CRb+
is the Rb+ content at the time point t; C
T is the maximal Rb+ content that can be loaded; τ is the time constant for the loading. For Rb+ efflux curves, the following equation of single exponential decay was used to get fitting parameters:
The dose-dependent response was fitted using normal Hill equation to generate IC50 (reduced signal) or EC50 (increased signal). All data analyses were performed using the software Origin® 6.1 (Origin Lab Corporation, Northampton, MA). Data are presented in mean ± SD.
Results and Discussion
HEK293-hTRPC6 stable cell line and HEK293-hTRPA1 stable cell line are used in this study. These 2 channels are members of TRP family and share some common features such as nonselective permeability but have the least amino acid identity within the family (<20%). To confirm the channel expression and validate the cell lines for assay development, we performed patch clamp recording to characterize the channel currents. As shown in Figure 1A , HEK293-hTRPC6 cell had a small current (<200 pA at ± 60 mV) at resting state. The application of carbachol (CCH), an M cholinergic receptor agonist, dramatically increased both inward and outward currents. The CCH effect appeared quickly but declined immediately after reaching the peak value. The declining process was slow and incomplete during 8-min treatment despite only small current retained. The reapplication of CCH evoked a small current increase, likely due to the depletion of the receptor channel coupling components. Figure 1B shows the voltage dependence of the currents obtained by the ramp voltage protocol. The CCH-induced current reversed at −3.65 ± 1.2 mV (n = 8), indicating that it is a nonselective current. The current amplitude was greater in the outward direction than in the inward direction and a slight dip was seen at the voltage from 0 to 30 mV, resembling the characteristic hallmarks of TRPC6 channels. Although the current was small without CCH, it had indeed the same voltage-dependent kinetics as the CCH-induced current, suggesting that the channel had basal opening. To examine Rb+ permeability of the channel, the external Na+ was replaced by the equal concentration of Rb+. No noticeable shift of reversal potential was observed, indicating that the channel was equally permeable to Na+ and Rb+. However, when Na+ was replaced by NMDG, the reversal potential was shifted to −62.1 ± 5.4 mV (n = 5), indicating that the channel was only permeable to cations. We were able to record the CCH-induced current with the amplitude ranging from 0.3 to 2 nA at 60 mV from 67% of the cells (8 of 12).
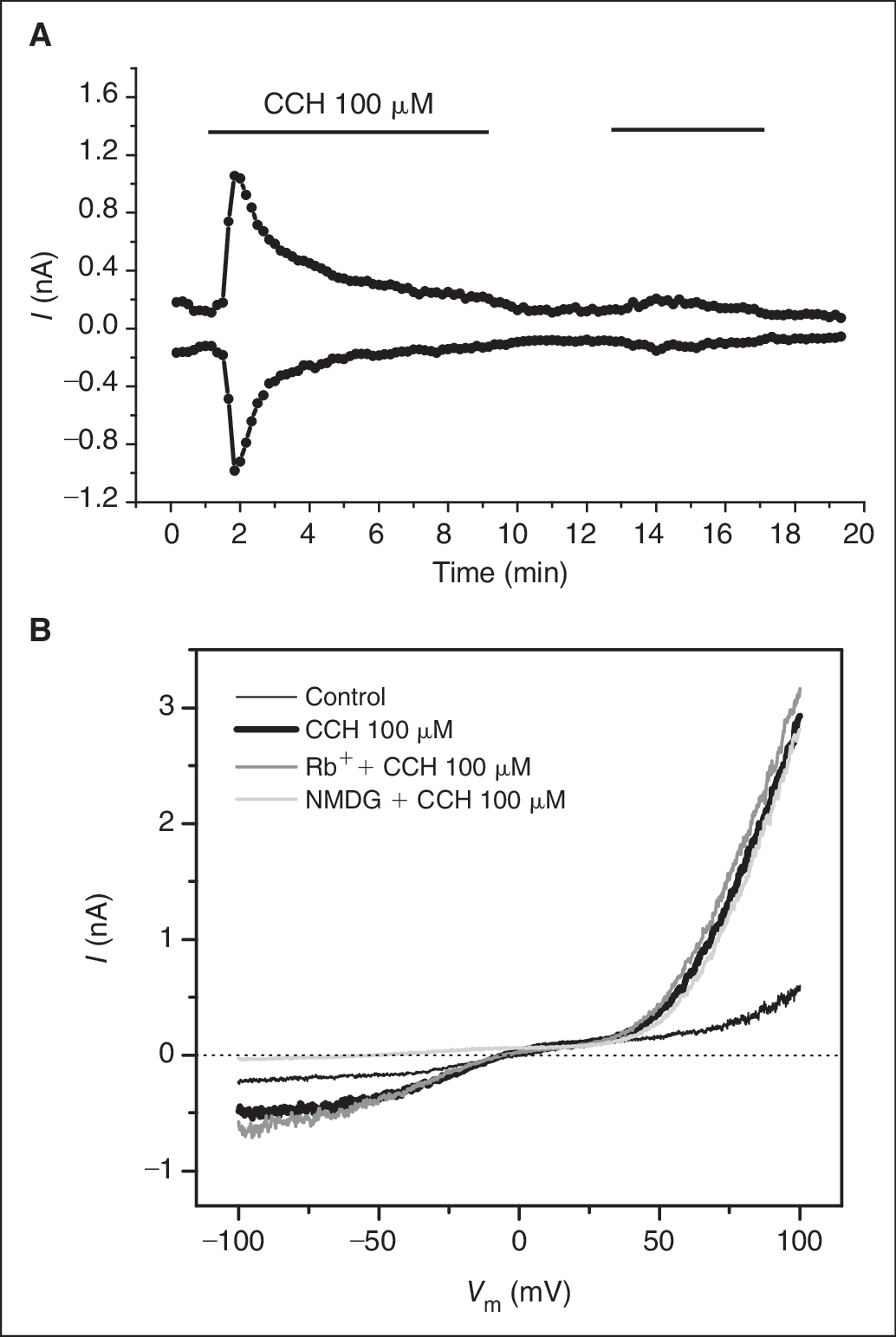
Patch clamp recording of TRPC6 stable cell line. (
For HEK293-TRPA1 cell line, we did the same experiment to characterize the current induced by known TRPA1 agonist, allyl isothiocyanate (AITC). As shown in Figure 2A , HEK293-TRPA1 cell had little current before the addition of AITC. AITC increased both inward and outward currents evoked by ± 60 mV membrane voltage. This process was relatively slow but the current was sustained. No spontaneous decline was observed. The increase in extracellular Ca2+ further potentiated the current and then suddenly decreased and abolished it. This phenomenon has been previously described as Ca2+ potentiation and inactivation. 21 –23 Figure 2B shows the voltage dependence of the current derived by the ramp voltage protocol. The AITC-induced current reversed at near 0 mV, indicating that this channel conducts a nonselective current. The channel was highly potentiated by Ca2+ before being completely inactivated, resembling the unique property of TRPA1 channel. Similar to TRPC6 channel, TRPA1 was also equally permeable to Na+ and Rb+ with no noticeable shift in reversal potential when Na+ was replaced by Rb+. NMDG significantly shifted the reversal potential to the left side, indicating the cationic permeability of the channel. The current evoked by 20 μM AITC appeared in every cell tested with the amplitude in the range of 0.4 to 3 nA at 60 mV (n = 12).
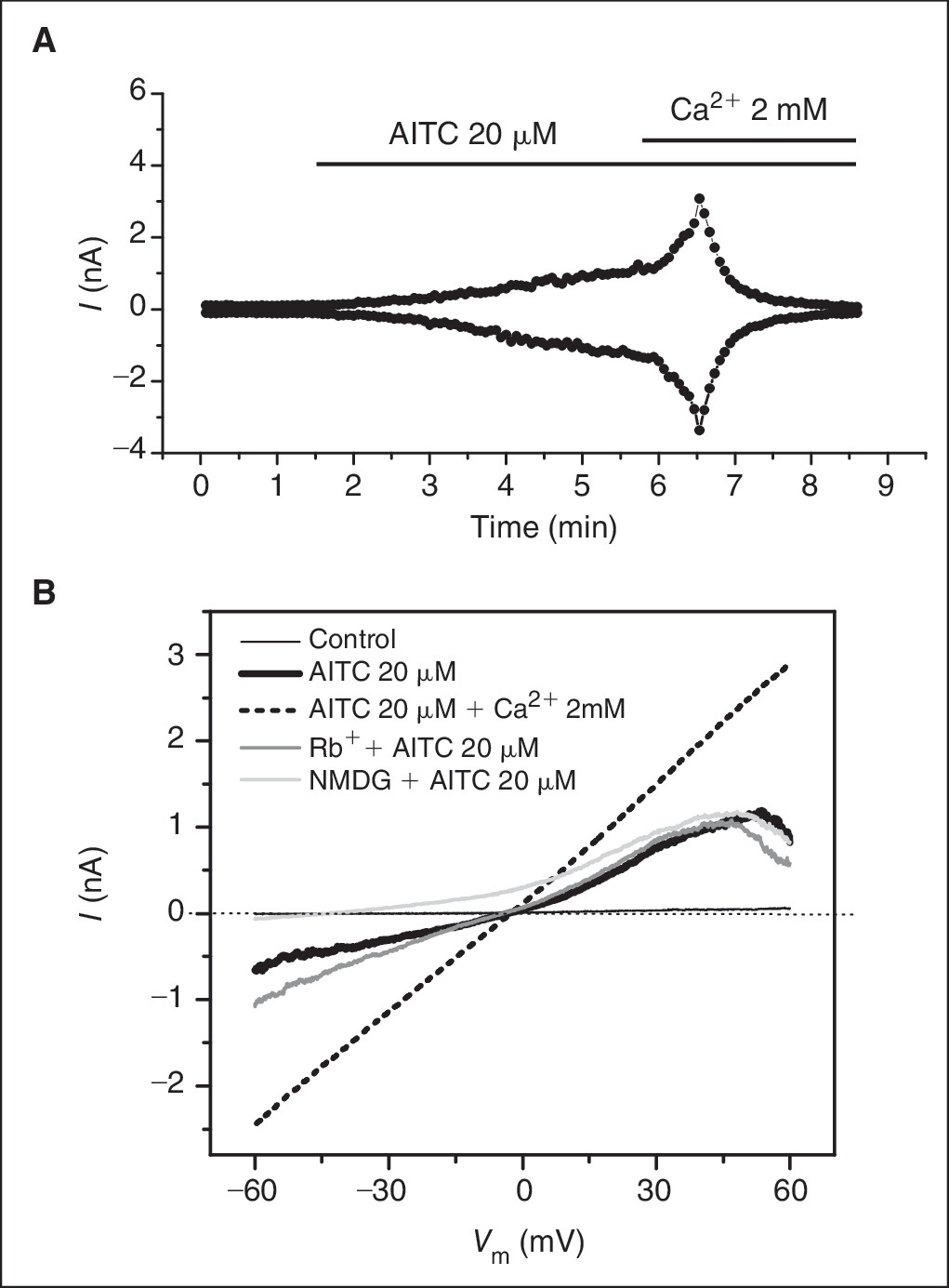
Patch clamp recording of TRPA1 stable cell line. (
Next, we started the Rb+ efflux assay development by examining the Rb+ loading process. In the presence of 5 mM of Rb+ in the loading buffer, the increase in intracellular Rb+ in both cell lines followed the single exponential growth equation ( Fig. 3A and 3B ). The time constants for HEK293-hTRPC6 and HEK293-TRPA1 are 45.5 and 64.9 min, respectively. The faster loading in TRPC6 cells may reflect the basal opening of the TRPC6 channels as described earlier.

Rb+ loading time course. Cellular Rb+ content was measured every 20 min after the cells were incubated with 5 mM RbCl. For each graph, the data were collected from 8 wells for each data point in a 96-well plate. The solid line represents the fitting curve using single exponential growth equation. (
Based on the loading kinetics, we chose the 3-h loading period for the Rb+ efflux determination. We then examined the basal Rb+ efflux in 1 h after Rb+ loading and the effect of channel agonists, CCH and AITC for TRPC6 and TRPA1, respectively, on Rb+ efflux. As shown in Figure 4A , under control condition, cellular Rb+ content decreased time dependently with a time constant of 13.8 min. CCH induced a significant increase in the rate of reduction. Fitting to the single exponential decay curve resulted in a time constant of 11.7 min. Fifty minutes after treatment of CCH, the cellular Rb+ content decreased to 45% of the control. Figure 4B shows the Rb+ efflux in TRPA1 cells in the absence and presence of AITC. Similar to TRPC6 cells, the cellular Rb+ content in TRPA1 declined following the single exponential decay, but the rate was slower with the time constant of 25.1 min, consistent with the observation that TRPA1 showed much less basal current than TRPC6. AITC at 50 μM induced a quick decline with the time constant of 3.9 min. After 30 min treatment with AITC, the cellular Rb+ content decreased to 8% of the control. The faster and more complete Rb+ efflux in TRPA1 cells was in agreement with the results acquired in patch clamp recording.
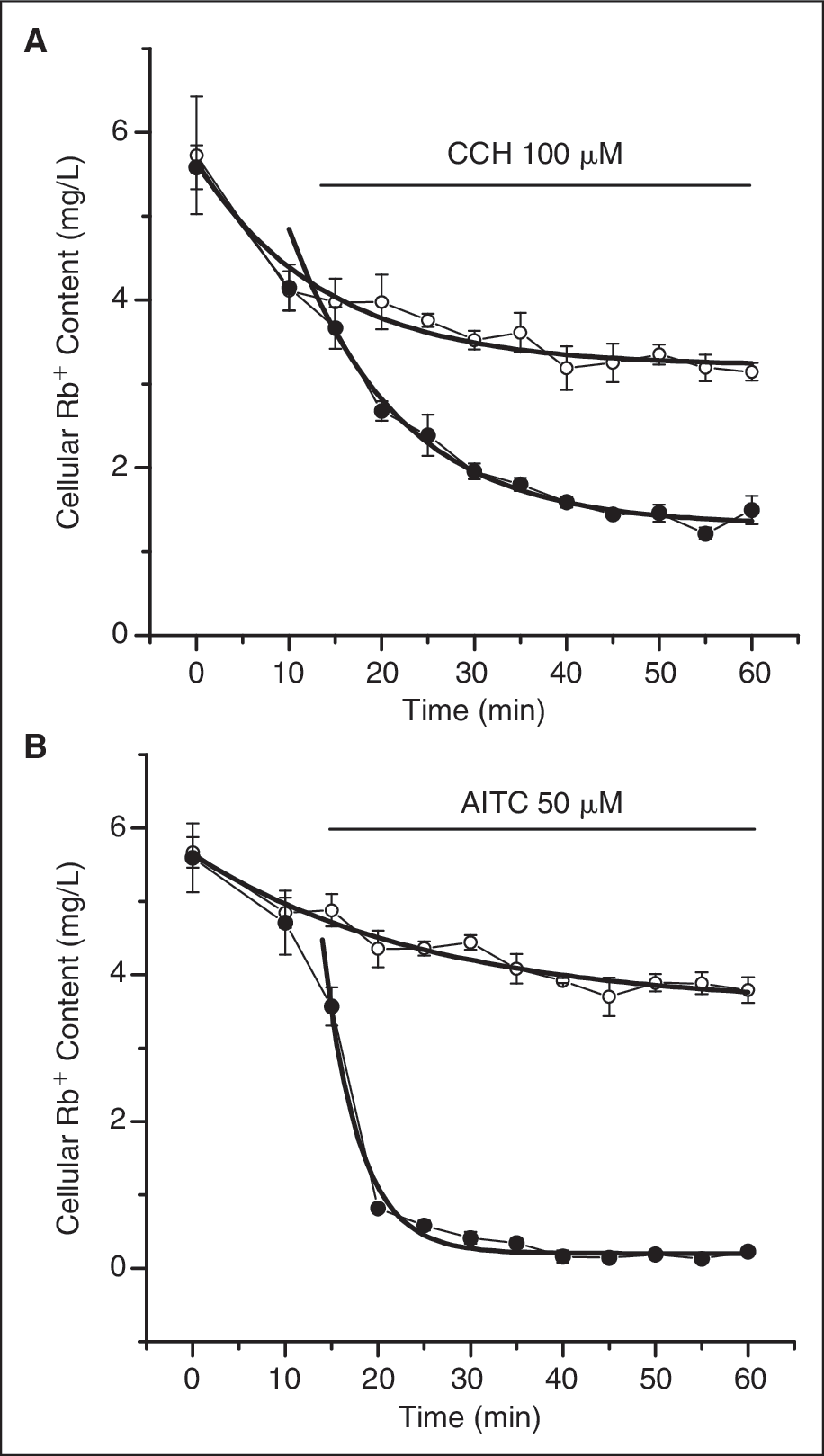
Time-dependent Rb+ efflux induced by the transient receptor potential (TRP) channel agonists. Immediately after washout of the loading buffer, Rb+ content was determined at each 5 min in 1 h except the first interval of 10 min. After the first 2 time point determinations, the agonists were added into the wells for agonist-treated group and the results were plotted together with the control group without agonist addition. For each data point, n = 8. The time course for both control and agonist-treated groups were fitted with single exponential decay equation and plotted in smooth solid lines. (
The dose-dependent effects of the agonists were examined in both cell lines. To test the specificity of the response, we cross examined the specific agonists in both cell lines. As shown in Figure 5A , CCH dose dependently decreased the cellular Rb+ content in TRPC6 cells with the EC50 at 3.6 μM. AITC had no effect in the same concentration range. In contrast, AITC evoked a dose-dependent reduction of the cellular Rb+ content in TRPA1 cells with the EC50 at 6.2 μM but had no effect on TRPC6 cells ( Fig. 5B ). These results further demonstrate that the responses induced by the agonists are specific to the corresponding channel activities.
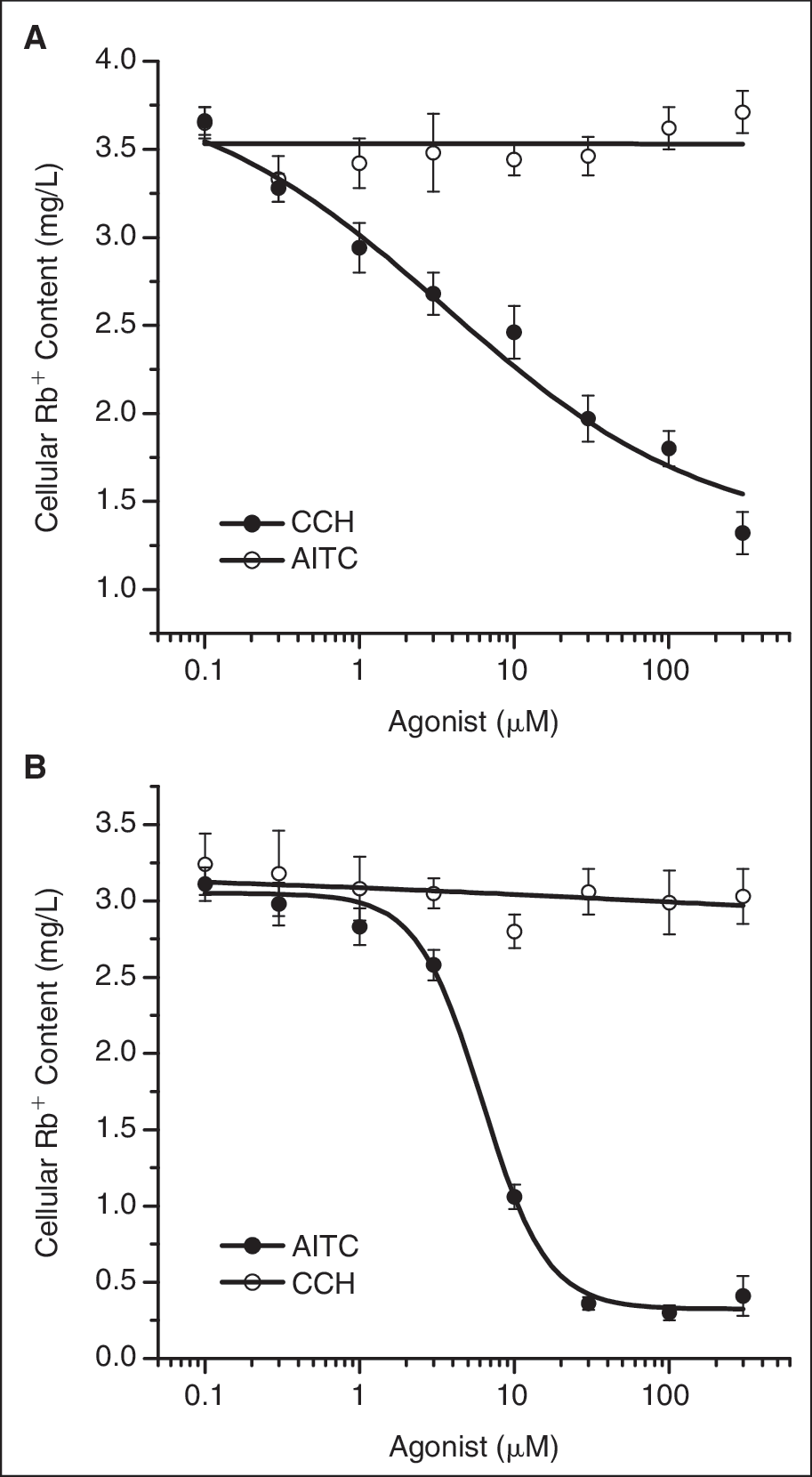
Dose-dependent effects of the TRP channel agonists. The cellular Rb+ content was plotted against the concentration of both channel agonists. The data are fitted with Hill equation. (
We next tested the Ca2+ effect on the Rb+ efflux since both TRPC6 and TRPA1 have been reported to be regulated by calcium ions. We increased the Ca2+ concentration to 1.5 and 5 mM from 0.15 mM that we used in the earlier experiments. As shown in Figure 6A , increasing Ca2+ in assay buffer for TRPC6 shifted the dose–response curve up without changing the potency of CCH effect, suggesting that Ca2+ inhibits the channel activity regardless of CCH effect. But, increasing Ca2+ reduced the potency of AITC in TRPA1 cells ( Fig. 6B ), indicating that Ca2+ may affect AITC-binding properties.
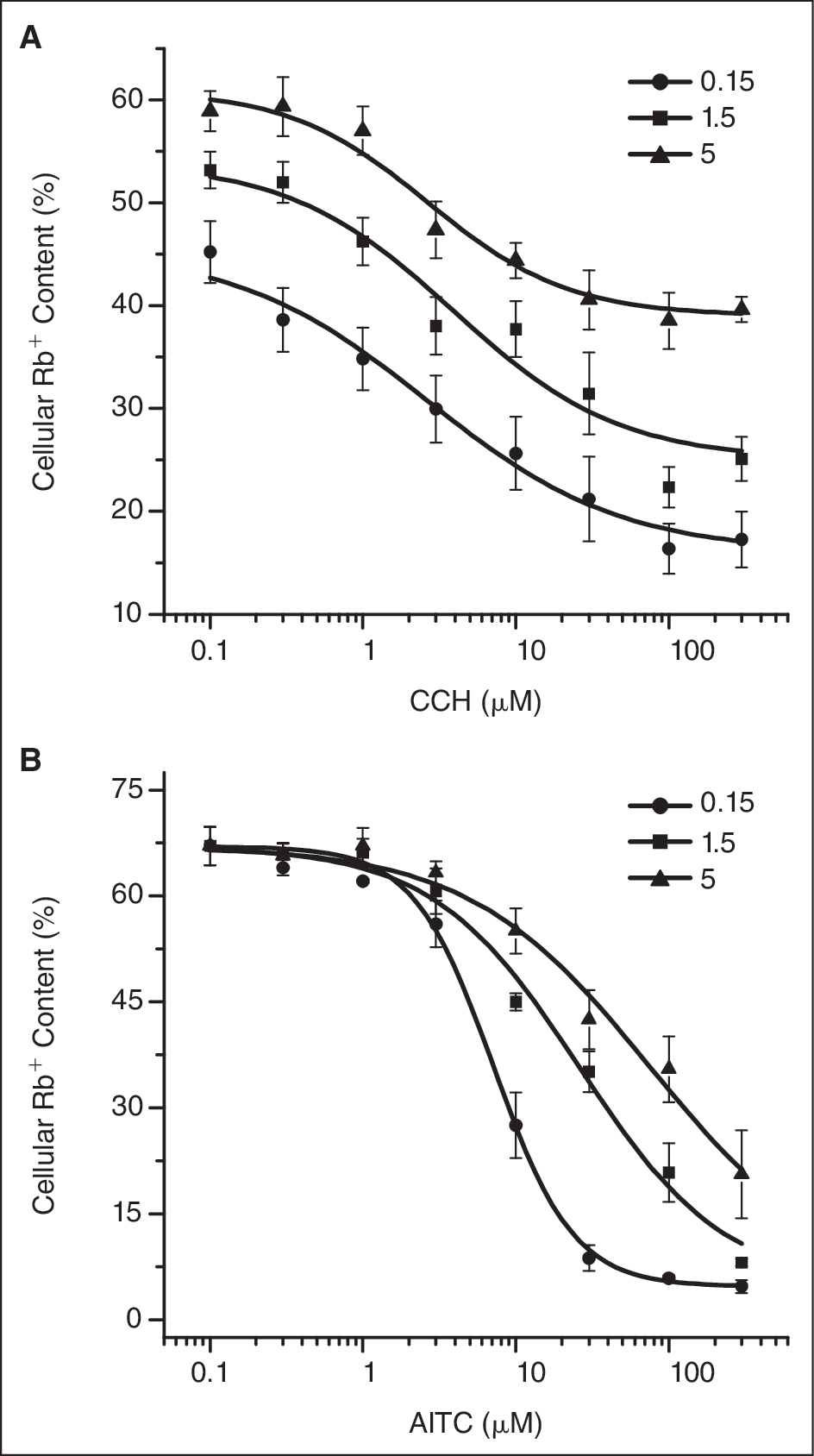
Effect of Ca2+ on Rb+ efflux induced by channel agonists. The experiment was performed in the presence of the different concentrations of Ca2+ as indicated and the dose–response curves were established as described in
Figure 5
, except that the data presented here are percent cellular Rb+ contents (see Materials and Methods). (
Using 100 μM of CCH as agonist in the presence of 0.15 mM of Ca2+, we examined the effect of the known TRPC6 blockers, SKF-96365, and ACA on TRPC6 cells. As shown in Figure 7A , both blockers dose dependently inhibited the response induced by CCH. The IC50s for these 2 compounds are 19.5 and 8.3 μM, respectively, comparable with the IC50s acquired in FLIPR membrane potential assay (6.2 and 14 μM, respectively, unpublished data). The calculated Z′ factors using maximal inhibition values are 0.67 and 0.55 for the 2 compounds, indicating that the assay is suitable for HTS (>0.5). Figure 7B shows the dose–response curves for our lead compounds derived from the primary HTS screen. These 2 compounds displayed potent dose-dependent inhibitions with IC50s at 0.34 and 0.72 μM, nearly 20-folds greater than the potencies of the known blockers. These values are also in accordance with the IC50s from FLIPR assay (0.52 and 0.72 μM, respectively, unpublished data). Figure 7C shows the raw data from a representative 96-well plate. The plate was treated with the buffer control and 100 μM ACA before addition of 100 μM CCH. The percent cellular Rb+ contents are 22.1 ± 3.2 and 55.4 ± 2.1, respectively. The Z′ factor is 0.53, indicating that the assay is robust for HTS.
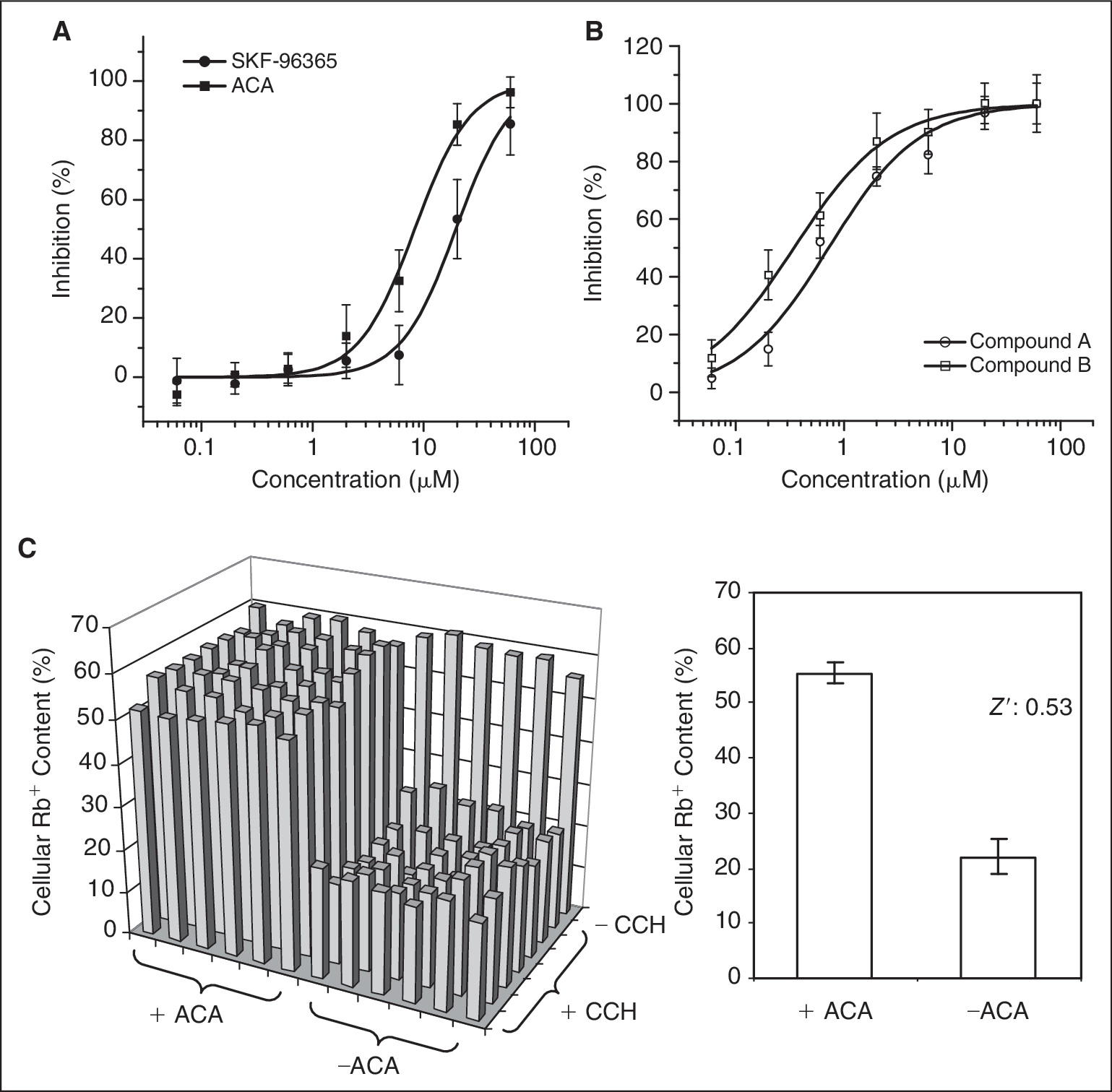
Effects of the TRPC6 channel blockers on CCH-induced Rb+ efflux in TRPC6 cells. (
For TRPA1 assay, we examined the effect of a relatively selective channel blocker, HC030031, on AITC-induced Rb+ efflux. 24 Because Ca2+ could significantly reduce AITC effect, we performed this assay in the presence of 0.15 and 1.5 mM of Ca2+ for comparison. Using 50 μM of AITC as agonist, HC030031 caused a dose-dependent increase in cellular Rb+ content under both Ca2+ conditions ( Fig. 8A ). However, increase in Ca2+ concentration remarkably shifted the dose–response curve to the left side. The IC50 was changed from 60 to 4.1 μM. Meanwhile, the assay window was significantly reduced from 51% to 27% in cellular Rb+ content due to a significant reduction in AITC response in the presence of 1.5 mM of Ca2+. Then we increased AITC to 300 μM and examined the effect of HC030031 in the presence of the different Ca2+ concentrations. As shown in Figure 8B , in 0.15 mM of Ca2+, HC030031 showed a weak effect with the maximal inhibition <10%. Increase in Ca2+ to 1.5 mM significantly improved the assay window. The further increase to 5 mM only slightly changed the assay window and the slope of the curve, but elevated both basal and maximal responses. The optimal Ca2+ condition for this assay would be 1.5 mM, not only because it provided a better assay window but also because this is within the range of physiological Ca2+ concentration. Under this condition, the IC50 for HC030031 is 30.8 μM. Figure 8C shows the raw data from a representative 96-well plate. The plate was treated with the buffer control and 100 μM HC030031 before addition of 300 μM AITC. The percent cellular Rb+ contents are 3.0 ± 0.6 and 68.6 ± 2.2, respectively. The Z′ factor is calculated to be 0.87, indicating that the assay is robust for HTS.
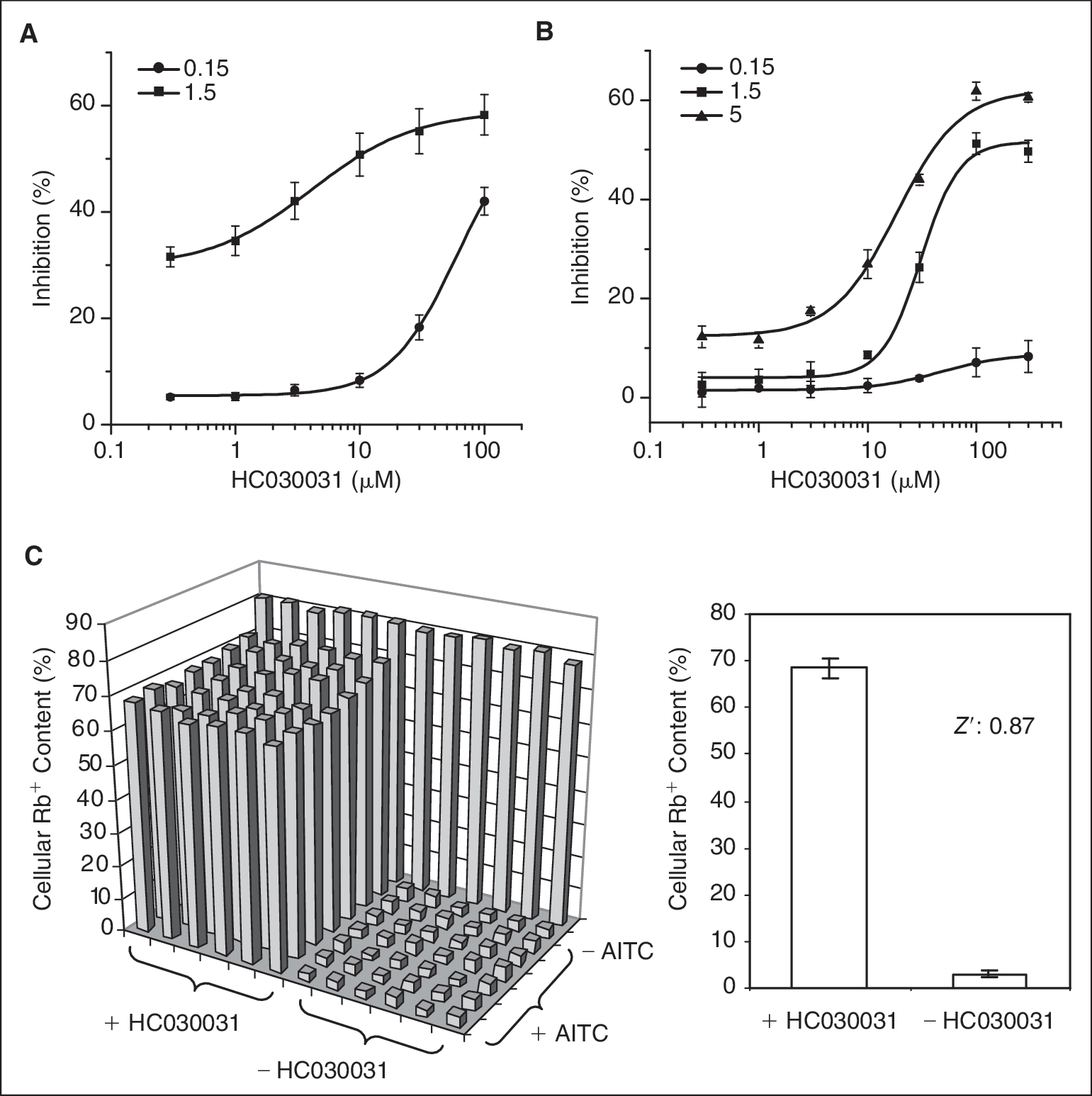
Effect of HC030031 on Rb+ efflux induced by AITC in TRPA1 cells. (
Taken together, in the present study, we have developed a robust high-throughput Rb+ efflux assay for NSCC channels using recombinant TRP channel cell lines. NSCC channels exist in a majority of cell types. The channel permeability to Rb+ is sufficient for developing this robust assay. Rb+ is often used for detection of potassium transport across membrane. The membrane Rb+ fluxes detected by either radioactive or nonradioactive approaches are often interpreted as a reflection of the specific mechanism for K+ transportation. 16,25 Based on the results from this study, NSCC channels are able to carry Rb+ upon channel activation. These channels normally provide ionic entries for Na+ and Ca2+ under physiological conditions. Although they can be equally permeable to K+, cells usually have membrane potential close to K+ reversal potential when the channels get activated. The K+ flux under such conditions is much weaker than Na+ and Ca2+. So, the Rb+ flux detected using this assay can be a result of the increased NSCC channel activities. Care should be taken when interpreting the data obtained from Rb+ efflux assay, especially when working with primary cells.
Conclusion
Rb+ efflux assay has been demonstrated in this study to be robust for measurement of NSCC channel activities. The examples presented as NSCC channels are TRPC6 and TRPA1, 2 recently identified potential drug targets. Due to the unique activation properties, the available assays for these channels especially in HTS format are very limited. Rb+ efflux assay is sensitive, label-free, and HTS applicable. It provides a robust way to bridge the gap between the extremely low-throughput patch clamp recording and the high-throughput fluorescence-based assay. In addition, it has the potential to be used as an alternative for primary HTS.
Footnotes
Acknowledgments
We thank Dr. Janet Paulsen and Wei Cao for their help in generating TRPC6 expression construct. We also thank Dr. Ravikumar Peri for providing the HEK293-hTRPA1 stable cell line.
Author Disclosure Statement
No competing financial interests exist.