
Abstract
p75NTR is a neurotrophin receptor that can mediate either survival or death of neurons depending on the cell context. Modulation of p75NTR is a promising strategy to promote neuronal survival for treatment of cognitive disorders such as Alzheimer's disease. Despite years of investigation into the signaling mechanisms of p75NTR, no p75NTR signaling assay has yet been developed that is compatible with efficient screening of small-molecule modulators. In this work, we developed a homogeneous cell-based assay for screening p75NTR modulators and studying p75NTR function. Stimulation of p75NTR-transfected cells using either nerve growth factor (NGF) or Pro-NGF resulted in an enhanced caspase-3 activity as assessed by cleavage of a fluorescent caspase-3 substrate. Optimization of the assay with respect to time, cell density, NGF and Pro-NGF concentration, and other factors provided a twofold increase in the caspase-3 activity compared to background. Withdrawal of serum during the NGF or Pro-NGF treatment period was found to be essential for p75NTR-dependent caspase-3 activation. We validated the method by demonstrating that a signaling-incompetent p75NTR mutant could not substitute for wild-type p75NTR in mediating caspase-3 activation. A focused library screen identified new inhibitors of p75NTR signaling. This method will be useful for identifying small-molecule modulators of p75NTR as well as further characterizing downstream signaling events.
Introduction
p75NTR signaling has long been associated with regulating neuronal survival and death. 6 –8 Paradoxically, p75NTR ligation appears capable of causing both survival and death depending on the cellular context. 9,10 Several factors tend to be associated with p75NTR signaling in its apoptotic mode; these include pro-neurotrophins (such as Pro-NGF) as ligands, a strong stimulus magnitude and duration, and the absence of Trk A within the cell of interest. 11 In its apoptotic mode, p75NTR ligation has been associated with activation of signaling factors such as c-Jun N-terminal kinase (JNK), p53, caspase-3, and TRAF6, which is an E3 ubiquitin ligase that has been reported to interact with the protein NRIF, resulting in the nuclear translocation of the complex. 12 –14 Conversely, factors associated with p75NTR signaling in its pro-survival mode include classical neurotrophins (such as NGF) as ligands, a small-stimulus magnitude and duration, and the presence of Trk A in the cell. 11 In its survival mode, p75NTR often seems to promote signaling through Akt and PI-3′-kinase and can also signal through RIP2 in Schwann cells to promote survival. 15 Since p75NTR itself lacks intrinsic enzymatic activity, the molecules that are activated downstream of p75NTR provide the best opportunity to study p75NTR receptor ligation in a cellular context.
Due to its role in regulating neuronal survival and death, it is unsurprising that p75NTR signaling has been associated with neurodegenerative diseases, particularly Alzheimer's disease. 16 –19 For instance, p75NTR expression has been demonstrated to be increased in Alzheimer's disease, and p75NTR is expressed by neuronal populations that are susceptible in Alzheimer's disease, including hippocampal, cortical, and basal forebrain cholinergic neurons. 16 Recent evidence indicates that Aβ peptides, whose accumulation in plaques is widely believed to cause Alzheimer's disease, can modulate p75NTR signaling. 17 Aβ has been shown to bind to p75NTR and induce apoptotic signaling downstream of p75NTR through JNK. 17 Furthermore, Aβ-induced neuronal degeneration in hippocampal and basal forebrain cholinergic neurons was attenuated in p75NTR-deficient mice. 16
There is evidence that p75NTR itself may be a viable therapeutic target for Alzheimer's disease. 20 Small-molecule ligands, designed based on loop 1 of NGF, have been reported to modulate p75NTR receptor function, promote survival of hippocampal neurons, and induce survival signaling pathways similar to neurotrophins. 21 Furthermore, these molecules also may antagonize the effects of Aβ on p75NTR by blocking deleterious Aβ-induced signaling and cell death. 22 While the data described above suggest that p75NTR is an attractive therapeutic target, one factor limiting further progress toward targeting p75NTR is the lack of a sufficiently robust cell-based assay to interrogate p75NTR function and screen for small-molecule modulators. In this work, we report for the first time a homogeneous assay to monitor p75NTR signaling in cells that is sufficiently robust for small-molecule screening.
Materials and Methods
Materials
Human p75NTR and TrkA were purchased from Origene, and they were expressed from the pCMV6-XL5 vector. Rat p75NTR was synthesized at Geneart and cloned into pCDNA3.1-hyg. The mutations were made by site-directed mutagenesis (C256A & C257A p75NTR). All plasmids were sequenced and verified by Elim Biopharm. Recombinant human Pro-NGF was obtained from Neuromics, catalog # PR27004. FuGENE 6 transfection reagent, catalog # 11815061001, was purchased from Roche. Recombinant human NGF was obtained from R&D Systems, catalog # 256-GF/CF. Epoxomicin, catalog # E3652, and Staurosporine, catalog # S6942, were purchased from Sigma. The sources of antibodies were as follows: anti-p75NTR from Advanced Targeting Systems, catalog # AB-N07, and anti-TrkA from Cell Signaling Technology, catalog # 2508. HEK293 cells, catalog # CRL-1573, were purchased from the American Type Culture Collection. Poly-D-lysine–coated 96-well plates, catalog # 3667, were obtained from Corning Incorporated. The Apo-ONE Homogeneous Caspase 3 activity assay kit, catalog # G7791, was purchased from Promega.
Cell Culture and Transfection
HEK293 cells were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 2 mM L-Glutamine, 1 mM sodium pyruvate, and 1% of 50 I.U./mL penicillin and 50 μg/mL streptomycin. Cells were passaged weekly at a 1:10 to 1:20 ratio of cell suspension to fresh media. One day before the transfection experiment, HEK293 cells were plated in poly-D-lysine–coated 96-well plates in 100 μL of medium. Cells were then transfected with plasmid DNA using procedures from Roche. Briefly, Roche FuGENE6 Transfection Reagent was diluted with the serum-free DMEM without antibiotics at a 1:13 ratio for 5 min at room temperature. Plasmid DNA was added to diluted FuGENE 6 Transfection Reagent at 3:1 ratio (0.39 μL, for FuGENE 6, and 0.13 μg for DNA). The transfection reagent–DNA complex was mixed and incubated for 45 min at room temperature before adding to the cells. In these experiments, the transfection efficiency was quantified as the percentage of cells with intensity more than twofold over the background level, irrespective of the total expression level. No effect of cell-passage number on the assay was observed.
Immunofluorescense Staining
After 48 h of transfection, cells were fixed with 4% formaldehyde in phosphate-buffered saline (PBS) containing Hoechst (1 μg/mL final) for 30 min at room temperature, washed with PBS, and permeabilized with 0.4% Triton in PBS for 10 min. Cells were washed with PBS, blocked with 6% goat serum in PBS for 1 h at room temperature, and then washed with PBS. Anti-p75NTR antibody (3 μg/mL) or anti-Trk A (1:500 dilution) in PBS containing 3% bovine serum albumin (BSA) was added to cells overnight. Cells were then washed in PBS and Cy3 Goat Anti-Mouse at 1:500 dilutions or Fluorescein Goat Anti-Rabbit secondary antibody at 1:300 dilution (Jackson ImmunoRsearch) was added in PBS with 3% BSA for 1 h at room temperature. Cells were then washed with PBS, and images were captured on the Cellomics ArrayScan VTi platform using Target Activation BioApplication to quantify the transfection efficiency or percentage of cells expressing p75NTR, green fluorescent protein (GFP), and TrkA. A cell was deemed to have a positive immunostain if its intensity was twofold over the background level.
Transient Transfection Assay Method
On day 0, HEK293 cells were plated in poly-D-lysine–coated 96-well plates at 2,000 cells per well in DMEM supplemented with 1% FBS. On day 1, cells were transfected with plasmid DNA at 0.13 μg DNA/well. On day 3, cells were rinsed with PBS and treated with 100 ng/mL of Pro-NGF or 200 ng/mL of NGF unless otherwise indicated in the serum-free medium. Expoxomycin (30 nM) and Staurosporine (300 nM) were pretreated 30 min before NGF and Pro-NGF addition. On day 4, caspape-3 activity in cells undergoing apoptosis was measured using the Promega Caspase 3 assay kit. In the presence of caspase-3 in cell lysates, a nonfluorescent substrate is cleaved, and the Rhodamine 110 leaving group becomes intensely fluorescent, which can be quantified by the LJL-Analyst plate reader with excitation and emission wavelengths set at 485 and 530 nm, respectively.
Pilot Compound Screen
In the pilot screen, each 96-well screening plate had six untreated controls and six wells treated with pro-NGF, which were used to calculate the Z′ 23 on each screening plate. In addition, each screening plate had four wells treated with Pro-NGF and 1 μM of compound 1, which served as a positive control compound on each plate. Finally, we also employed an independent apoptosis control in two wells using 30 nM epoxomicin. Compounds were applied to cells at a 1 μM final concentration for 24 h and were screened in a single-point mode. The final dimethyl sulfoxide (DMSO) concentration was set at 0.1%. The caspase-3 activity in cells undergoing apoptosis was measured using the Promega Caspase-3 assay kit. Compounds that resulted in a 5-standard deviation (SD) decrease from the caspase-3 signal observed in the pro-NGF-treated wells were scored as hits.
Measuring Cell Health Using Automated Fluorescence Imaging
HEK293 cells were plated at 2,000 cells/well in a 96-well Corning tissue culture plate overnight and were challenged with compounds at a final concentration of 1 μM in 0.1% DMSO for 52 h. Subsequently, prewarmed paraformaldehyde at a final in-well concentration of 4% was added to the wells for 30 min. The wells were aspirated and then washed three times with PBS. The fixed cells were stained with a 1 μg/mL concentration of Hoechst dye for 30 min and then washed three times with PBS. The plate was read with an Array Scan VTI (Cellomics, Pittsburgh, PA) using Cellomics Cell Health Profiling Bioapplication v.2. The parameters of object count/field and nuclear morphology were then automatically determined by Bioapplication. We have found that object count/field to be a sensitive parameter of cell health because if cells are undergoing nonlethal cell injury, they tend to detach from the plate immediately after the addition of paraformaldehyde.
Results
Establishing Transfection Conditions for the p75NTR-Signaling Assay
To develop a homogeneous cell-based assay for p75NTR, we sought to employ a cell line that is highly amenable to screening and focused attention on HEK293 cells. We noted that p75NTR transfected into HEK293 cells had been reported in the literature to mediate cleavage of caspase-3 upon stimulation of cells by NGF, as assessed by Western blotting. 5 Hence, we examined whether p75NTR transfected into HEK293 cells increases the caspase-3's enzymatic activity in these cells. The activity of caspase-3 in cell lysates was evaluated using a caspase-3 substrate that is nonfluorescent in its uncleaved form, but becomes fluorescent upon cleavage by caspase-3. 24
Initial efforts focused on optimizing the procedure to transfect p75NTR into cells. A p75NTR construct was initially transfected alongside two control constructs, one containing an empty vector and the other containing GFP as a control (Fig. 1). All cells were fixed, and then the p75NTR- and empty-vector-transfected samples were immunostained with a monoclonal antibody directed against p75NTR. For the p75NTR-transfected samples, it was observed that 30% of cells had been transfected after 24 h (Fig. 1B, D). In contrast, no staining was observed using the empty vector control, indicating that HEK293 cells do not endogenously express p75NTR, and p75NTR expression is not induced by the transfection process (Fig. 1A, D). The 30% transfection efficiency obtained with p75NTR was more than twofold less than that obtained using GFP as a control (Fig. 1C, D).
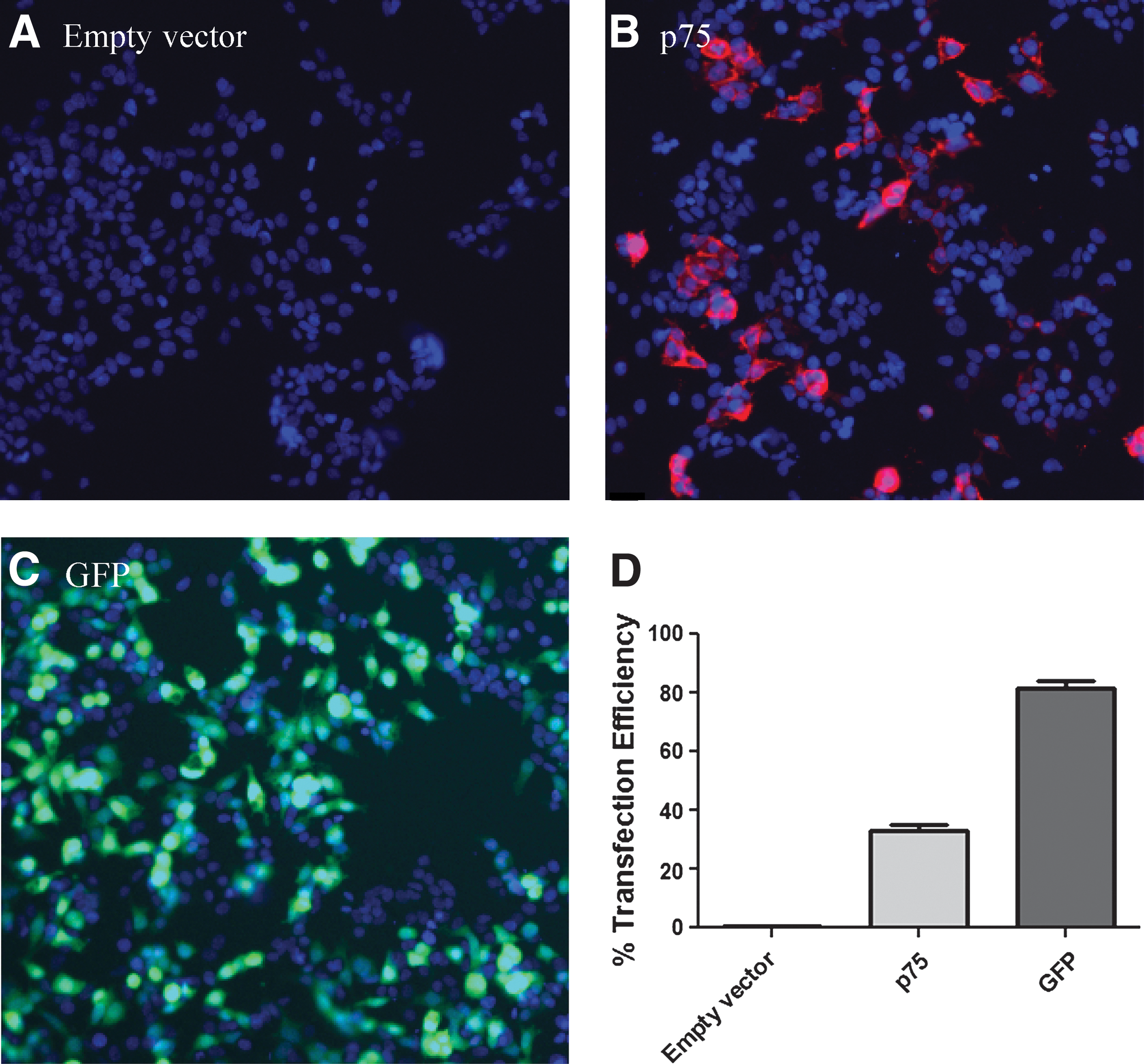
Transfection efficiency of the human p75NTR construct.
Several additional experiments were undertaken to optimize the p75NTR transfection procedure. To specifically optimize the p75NTR immunostaining, differing amounts of p75NTR antibody were employed, and a concentration of 3.75 μg/mL (the lowest concentration examined) was found to provide a sufficiently strong staining intensity (Fig. 2A). The amount of p75NTR DNA utilized for transfection was varied, and a DNA concentration of 0.13 μg/well was found to be the least amount required to provide 30% transfection efficiency at 24 h (Fig. 2B). It was also determined that the transfection efficiency was not observed to be affected by the density of cells on the plate (Fig. 2C). Finally, the transfection efficiency was examined at both 24 h and 48 h after transfection (Fig. 2D). The transfection efficiency was found to improve to 50% at 48 h (Fig. 2D) and remain near that level for up to 96 h, so subsequent experiments were performed using a time of 48 h between transfection and either NGF or Pro-NGF stimulation.
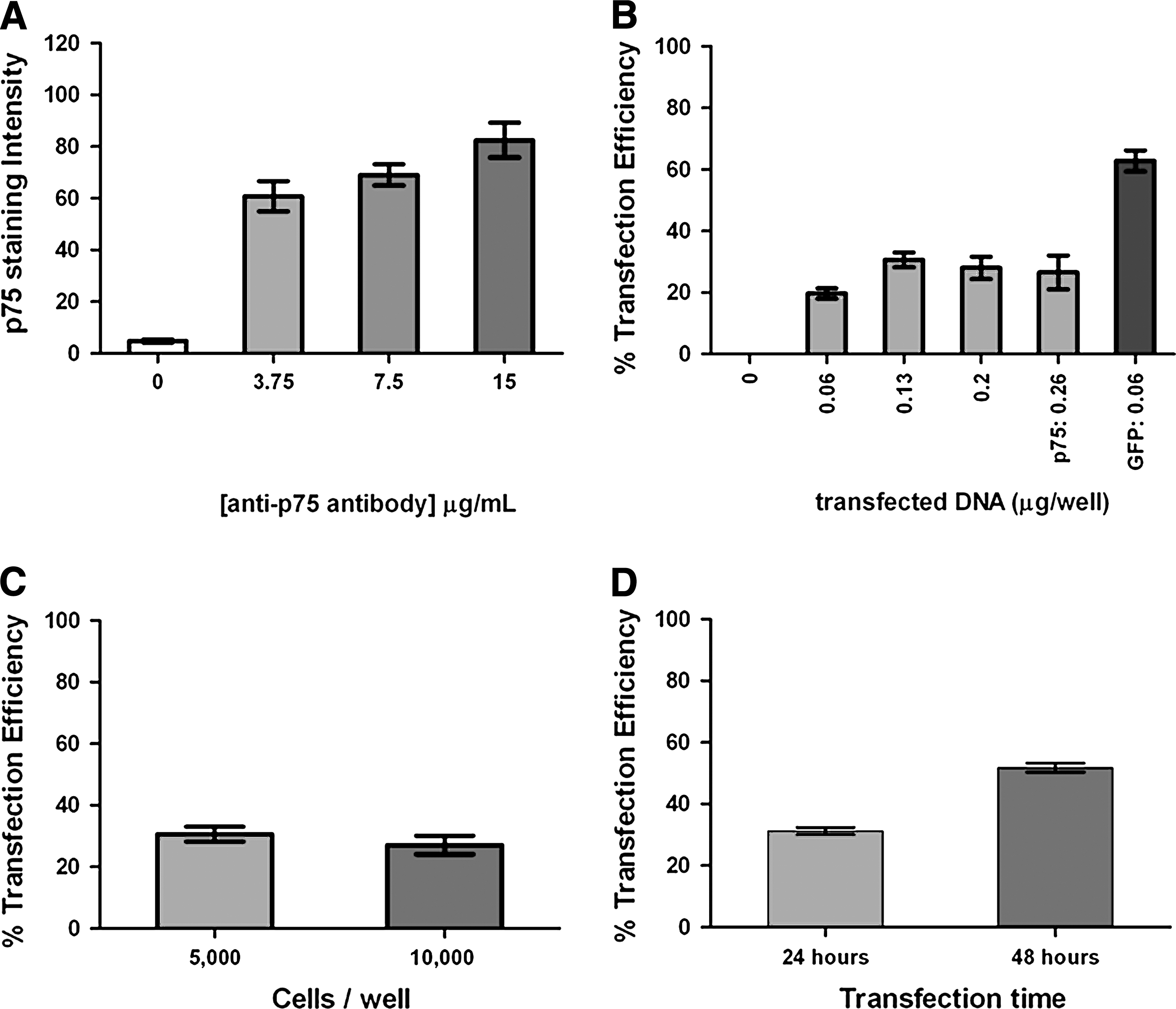
Optimization of the transfection conditions for the p75NTR construct.
Stimulation of p75NTR-Transfected Cells with Pro-Ngf or Ngf Increases the Caspase-3 Activity
It was next explored whether stimulation of p75NTR-transfected cells with various neurotrophins would result in caspase-3 activation. Initial experiments were performed with Pro-NGF as a ligand due to the reported relationship between Pro-NGF- and p75NTR-mediated apoptotic signaling. 11 In addition to Pro-NGF, cells were also stimulated with control compounds known to be associated with apoptosis and caspase-3 activation; these compounds were the proteasome inhibitor epoxomicin and the nonspecific kinase inhibitor staurosporine. 25,26 A 76% increase in the caspase-3 activity over baseline was observed after treatment of p75NTR-transfected cells with 100 ng/mL Pro-NGF (Fig. 3A, left), suggesting that p75NTR receptor ligation causes caspase-3 activation. In contrast, cells transfected with an empty vector control (rather than p75NTR) and then stimulated with 100 ng/mL Pro-NGF showed no increase over baseline in caspase-3 activity (Fig. 3A, right). This finding indicates that Pro-NGF-mediated caspase-3 activation is specifically mediated by p75NTR. Treatment of p75NTR-transfected cells with either 30 nM epoxomicin or 300 nM staurosporine also resulted in caspase-3 activation at levels of 218% and 62% over background, respectively (Fig. 3A, left). However, unlike Pro-NGF, epoxomicin and staurosporine were also found to induce caspase-3 activation in cells transfected with an empty vector (Fig. 3A, right); this finding was expected given that epoxomicin and staurosporine do not require p75NTR to activate caspase-3.

Response of p75NTR-transfected cells to NGF and Pro-NGF. ) HEK293 cells. Caspase-3 activity was modulated by stimulating either p75NTR-transfected or empty vector-trasfected cells with no stimulation (control), 100 ng/mL Pro-NGF, 30 nM epoxomicin, or 300 nM staurosporine.
Subsequent experiments sought to determine the dose–response relationship between Pro-NGF stimulation and p75NTR-mediated caspase-3 activation and also establish whether NGF could function like Pro-NGF and induce p75NTR signaling in these cells. Titrating the amount of Pro-NGF used to stimulate p75NTR-transfected cells revealed an EC50 of 5.3±2.5 ng/mL with a full response of 80% over background evident at 100 ng/mL (Fig. 3B). Furthermore, stimulation of p75NTR-transfected cells with NGF also resulted in an increase in caspase-3 activity. The increase in caspase-3 activity upon stimulation by NGF was also dose dependent with an EC50 of 8.0±7.1 ng/mL and a maximal response of 57% over background (Fig. 3B). The greater efficacy of Pro-NGF compared to NGF for stimulating p75NTR signaling is consistent with previous findings reported in the literature. 9,27
Optimization of the p75NTR Transient Transfection Assay
Having established that neurotrophins mediate caspase-3 activation in p75NTR-transfected HEK 293 cells, we next set out to establish the most robust conditions for the assay. We focused on three aspects of the procedure: (1) the neurotrophin stimulation time, (2) the density of cells in the well, and (3) the presence or absence of serum during stimulation. p75NTR-transfected cells were first stimulated with 100 ng/mL Pro-NGF for 4, 8, 12, 24, and 48 h, and the level of caspase-3 activity was examined. It was determined that maximal caspase-3 activity was observed after 24 h of Pro-NGF incubation (Fig. 4A). Similar findings were observed upon NGF stimulation of p75NTR-transfected cells (Fig. 4B). A second factor that proved to be essential to observing p75NTR-mediated caspase-3 activation by neurotrophins was the cell density. Varying the concentration of cells in the well revealed a bell-shaped dependence of caspase-3 activity on cell density (Fig. 4C); a cell density at initial plating of 2,000 cells/well provided a greater percent increase in caspase-3 activity upon NGF treatment than 1,000 or 4,000 cells/well (Fig. 4C). Surprisingly, at an initial cell density of 6,000 cells/well, no increase in caspase-3 activity was observed upon NGF stimulation (Fig. 4C). Hence, the ability of neurotrophins to mediate induction of caspase-3 activity is strongly dependent on the cell density at plating. This finding is supported by previous studies. 28 Finally, it was also determined that activation of caspase-3 in p75NTR-transfected cells requires that neurotrophin stimulation be performed in serum-free media (Fig. 4D). In this experiment, p75NTR-transfected HEK293 cells were stimulated with 10 ng/mL Pro-NGF while in either serum-free media (as performed in experiments described thus far) or media containing 0.3%, 1%, 2%, 3%, or 10% fetal bovine serum. Cells stimulated with Pro-NGF in serum-free media demonstrated a 1.5-fold increase in caspase-3 activity compared to unstimulated cells, while cells stimulated in the presence of 0.3% or greater amounts of serum showed no increase in caspase-3 activity (Fig. 4D). This finding is consistent with the notion that for p75NTR to mediate an NGF- or pro-NGF-dependent apoptosis, cells must be under stressful conditions. Together, these experiments led us to our optimized protocol, which is shown in Table 1.
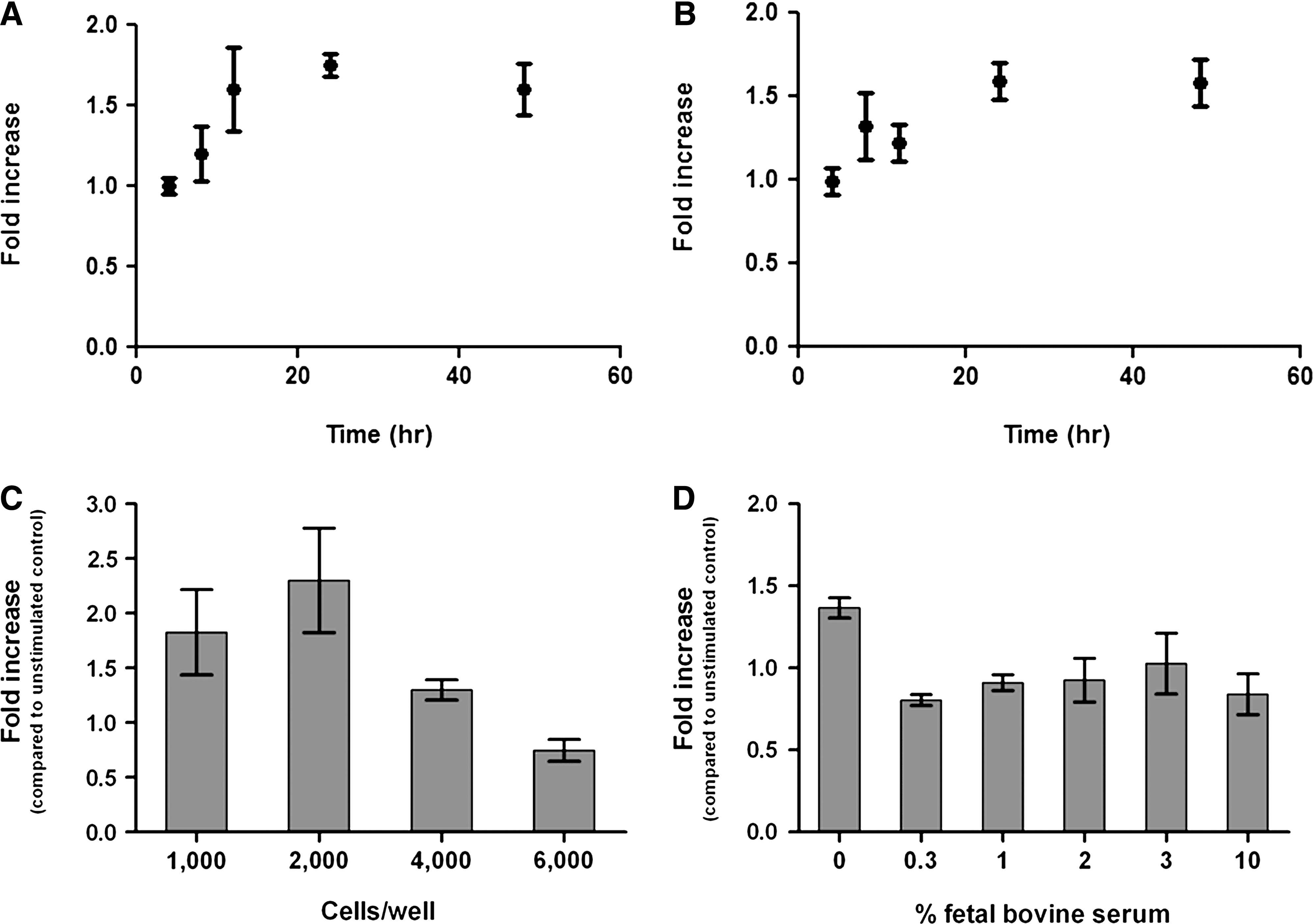
Optimization of the time, cell density, and serum concentration for p75NTR-mediated caspase-3 activation.
Protocol
FBS, fetal bovine serum; p75NTR, p75 neurotrophin receptor; NGF, nerve growth factor.
Validation of the p75NTR Transient Transfection Assay
After establishing optimized assay conditions, we next set out to definitively validate that the increase in the caspase-3 activity that occurs after NGF or Pro-NGF treatment that was attributable to p75NTR receptor ligation and signaling. To accomplish this, experiments were performed utilizing a signaling-incompetent p75NTR mutant. This point mutation (Cys 256 to Ala) has recently been demonstrated to abrogate downstream p75NTR signaling without modulating the ability of p75NTR to bind NGF 5 . We observed that the p75NTR C256A mutant was expressed in cells (Fig. 5A, B, E), but, unlike wild-type p75NTR, demonstrated no increase in caspase-3 activity after treatment with either pro-NGF (Fig. 6A) or NGF (Fig. 6B). Exploring for ways to improve the assay also led us to investigate rat constructs, rather than human constructs, of p75NTR. Both the wild-type rat p75NTR construct and a signaling-incompetent rat mutant (C257A) were well expressed in HEK293 cells (Fig. 5C, D, E). Transfection of wild-type rat p75NTR caused an increase in caspase-3 activation after NGF and Pro-NGF treatment, while the rat C257A mutant did not show a neurotrophin-mediated increase in caspase-3 activation (Fig. 6C, D). Finally, we demonstrated that cells transfected with Trk A also showed no increase in caspase-3 activity after treatment with NGF (data not shown). Together, these experiments indicate that the increase in caspase-3 activation that is observed in HEK293 cells in response to NGF and Pro-NGF is specifically attributable to p75NTR.
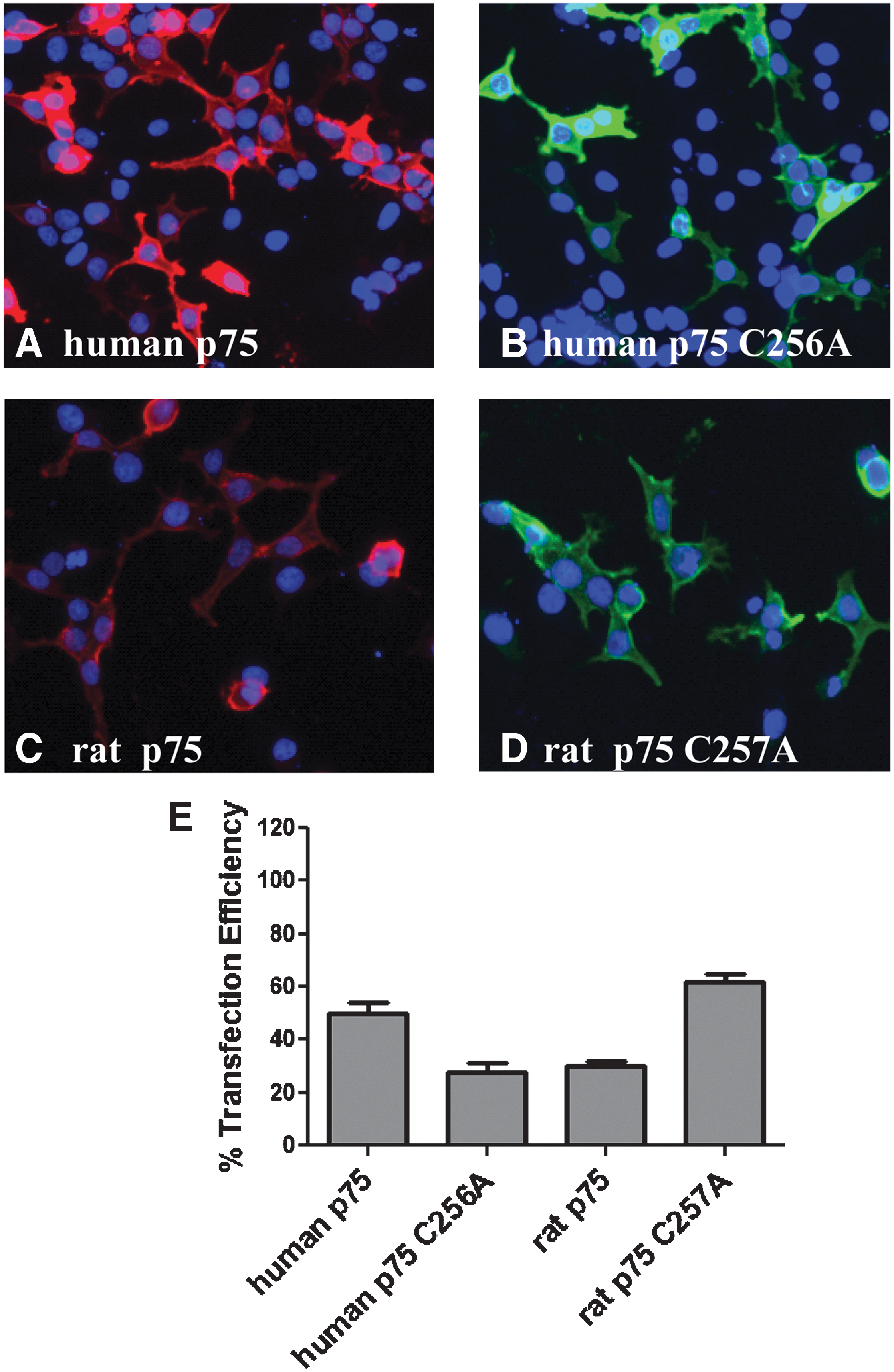
Transfection efficiency of various forms of p75NTR and Trk A.

Assay validation using p75NTR mutant constructs. ) or human p75NTR C256A (□) that received either no stimulation (control) or stimulation with 100 ng/mL Pro-NGF.  ) or human p75NTR C256A (□) that received either no stimulation (control) or stimulation with 100 ng/mL NGF.
) or human p75NTR C256A (□) that received either no stimulation (control) or stimulation with 100 ng/mL NGF.  ) or rat p75NTR C256A (□) that received either no stimulation (control) or stimulation with 100 ng/mL Pro-NGF.
) or rat p75NTR C256A (□) that received either no stimulation (control) or stimulation with 100 ng/mL Pro-NGF.  ) or rat p75NTR C256A (□) that received either no stimulation (control) or stimulation with 200 ng/mL NGF. Error bars represent the SD of 3 or more independent experiments.
) or rat p75NTR C256A (□) that received either no stimulation (control) or stimulation with 200 ng/mL NGF. Error bars represent the SD of 3 or more independent experiments.
The p75NTR/Caspase-3 Activation Assay Is Sufficiently Robust to Characterize a Small-Molecule Antagonist of p75NTR Signaling
After validation of the assay, efforts were initiated to identify a small molecule that modulates p75NTR signaling. Since the JNK-signaling pathway has been reported to link p75NTR activation to caspase-317, it was examined whether a commercial JNK inhibitor would block neurotrophin-mediated caspase-3 activation in p75NTR-transfected cells. Indeed, SP600125 was found to block NGF-mediated caspase-3 activation in a dose-dependent manner (Fig. 7). The average IC50 was found to be 0.13±0.02 μM from evaluation of the IC50 on four separate occasions. In these experiments, the assay gave an attainable Z′ of 0.25 and hence was sufficiently robust to identify inhibitors of p75-mediated caspase-3 activation. This finding demonstrates that the p75NTR/caspase-3 assay can be utilized to characterize small-molecule modulators of p75NTR.
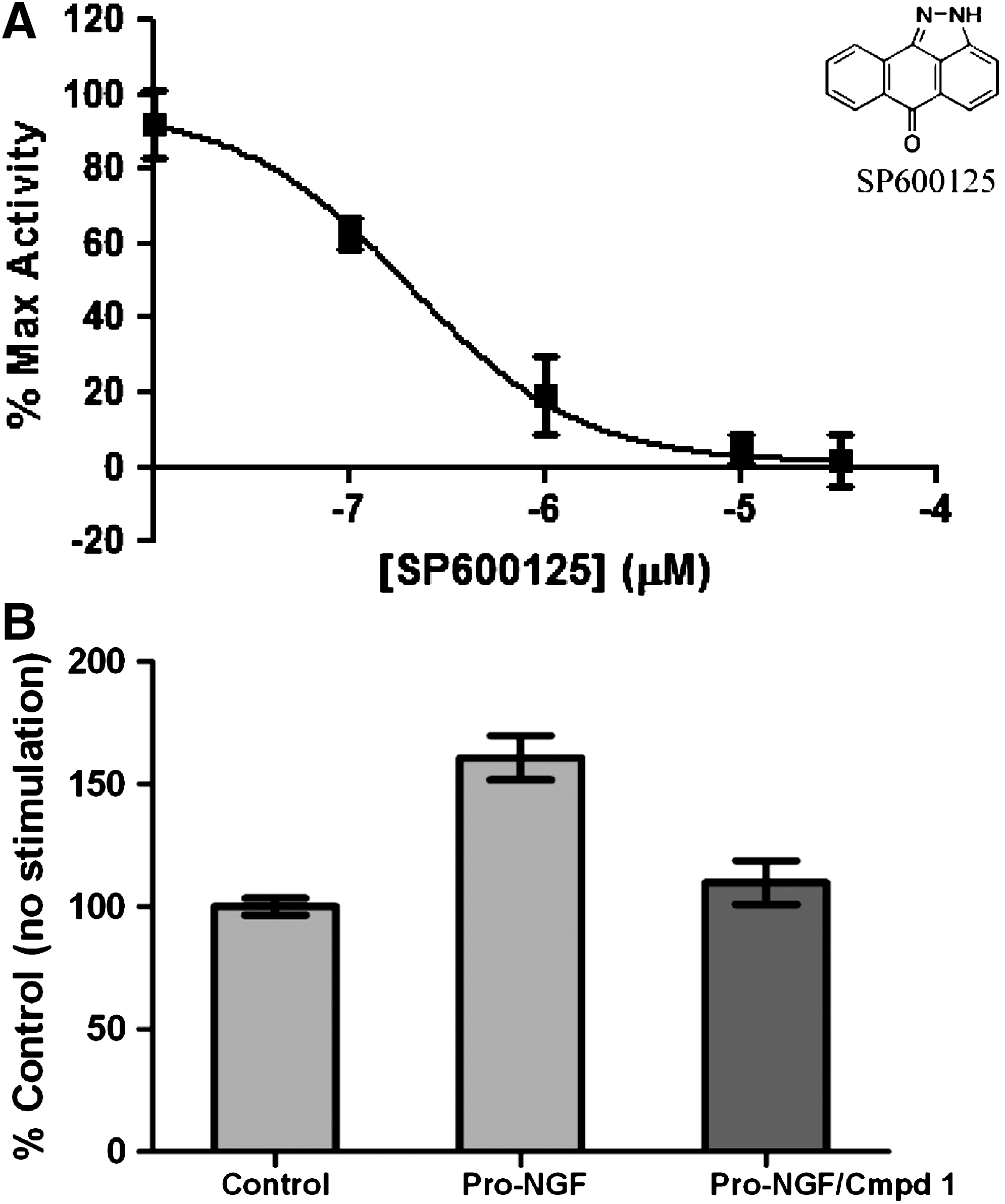
Utilization of the p75NTR/caspase-3 activation assay for characterization of small-molecule p75NTR antagonists.
Validation of the p75NTR/Caspase-3 Assay Using a Pilot Screen
Having preliminarily validated the ability of the assay to characterize small-molecule modulators of p75NTR signaling using SP600125, we sought to further validate the assay utilizing a pilot compound screen. An initial small-scale effort revealed a molecule (which we refer to as compound 1) that provided an 85% reduction in Pro-NF-mediated caspase-3 activation in p75NTR-transfected cells when tested at a concentration of 1 μM (Fig. 7B). After this initial effort, we then screened 168 compounds from our proprietary pilot screening set also using a final compound concentration of 1 μM and a Pro-NGF concentration of 200 ng/mL. The activities of the compounds in the screen (relative to the Pro-NGF stimulated control at 100%) are shown in Figure 8. Using a hit threshold of 5 SDs, we observed 13 hits in this screen. The activities of the hits are shown in Table 2. Of the 13 hits, 5 contained a common core scaffold (referred to as Scaffold I, see Table 2).

Activity of compounds from the pilot screen. Shown is a scatter plot of the percentage of caspase activation (y-axis) versus compound number (x-axis). Data were normalized such that 100% is the mean activity of Pro-NGF-stimulated cells, while 0% is the mean activity of unstimulated cells. The dashed line indicates the hit threshold at 5 SDs, which was <24% caspase activity. Compound 25, compound 45, compound 55, and compound 120 are shown at the bottom of the graph, since their % caspase activation values are ≤1% of the total caspase activity.
High-Throughput Screening Active Molecules
Following the screen, we chose 4 representative compounds (compounds number 120, 145, 146, and 155) and performed a 4-point dose–response curve on these active molecules (Fig. 9). Each dose–response curve confirmed the activity of the compound and provided IC50 values for each compound that ranged from 0.17 μM to 0.002 μM (Fig. 9 ). The IC50 value of compound 1 was also determined on multiple occasions and found to reproducibly provide an IC50 value of 0.32 μM. To characterize the robustness of the screen, we evaluated the Z′ from the different screening plates (see Materials and Methods). We found that the average Z′ between the 4 plates was 0.28, consistent with our initial results reported above. The independent Z′ determinations were 0.36, 0.22, 0.25, and 0.29, indicating reproducible behavior between different test occasions. In addition, Table 3 highlights the %CV of the positive and negative controls and the 3-SD hit thresholds for plates 1–4 for this pilot screen. The %CV of the pro-NGF-treated samples ranged from 4.6 to 10.2 and the %CV of the untreated controls ranged from 5.5 to 11.6. Hence, while the Z′ value of this screen was modest, and the %CVs allowed us to select a reasonable hit threshold and identify modulators.
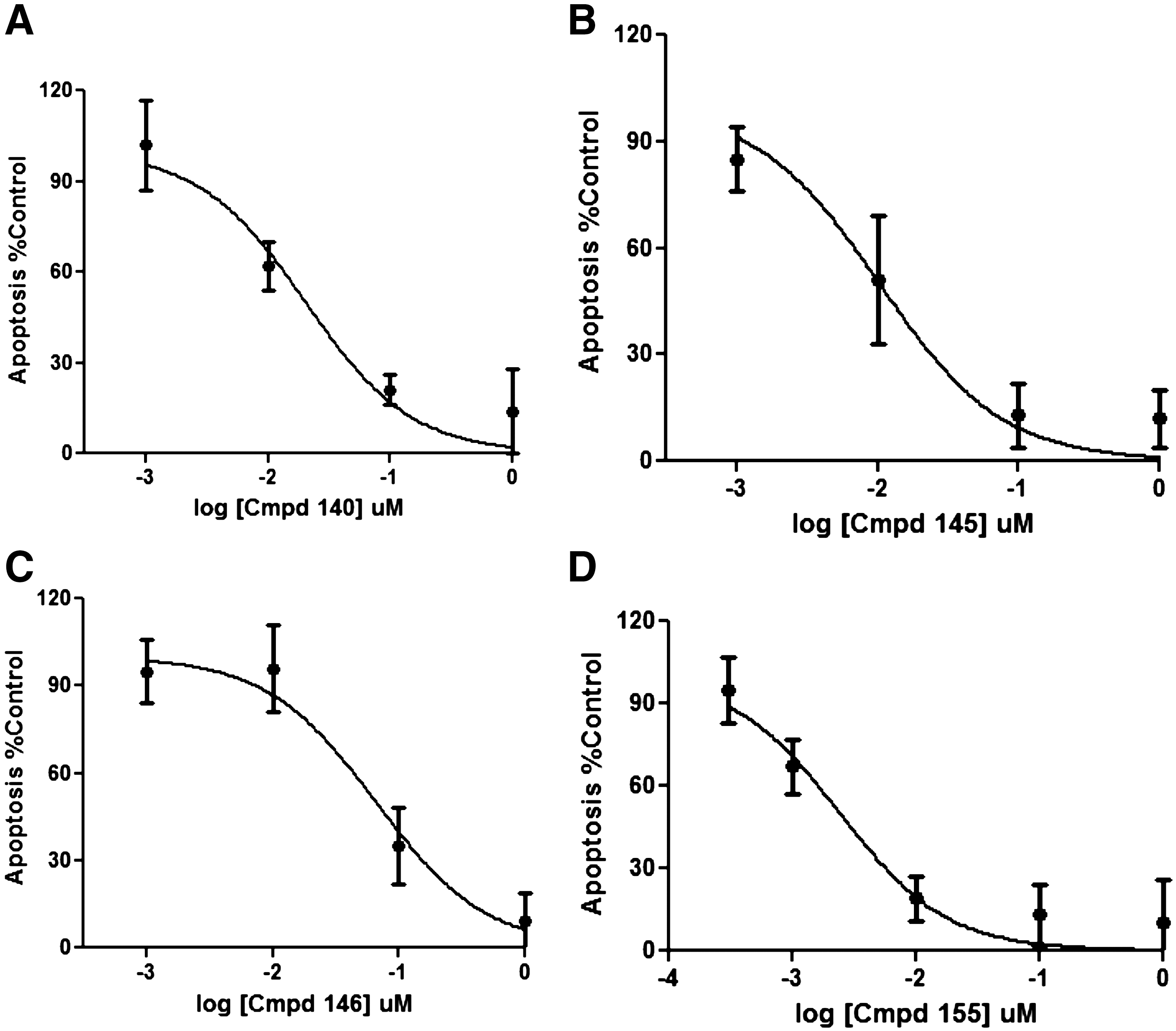
Dose–response curves for inhibition of apoptosis (as assessed using caspase-3 activity) by representative active compounds from the pilot screen.
High-Throughput Screening Data
We also undertook efforts to ensure that the activity of our active compounds was due specifically to an effect on p75NTR signaling, not nonspecific cytotoxicity. An established cell-health assay (see Materials and Methods) was utilized to demonstrate the lack of toxic effect of the 4 representative compounds characterized for IC50 above. All cells treated with these compounds showed similar cell counts/field as untreated cells, and all compounds also did not change nuclear morphology. Hence, the data suggest that the anti-apoptotic effects of the 4 tested compounds are not due to cytotoxic effects. Finally, we also tested whether these same set of 4 compounds could simply be caspase inhibitors or otherwise artifacts that interfere with the fluorescent read-out. To do this, we again used epoxomicin and staurosporine to induce caspase activation in a manner that is not mediated through p75NTR (Fig. 3A). We observed that each of the 4 compounds fails to block both epoxomicin-mediated and staurosporine-mediated caspase activation (data not shown), confirming that the activity of the compounds is not due to caspase inhibition or interference with the fluorescent read-out.
Discussion
It is well known that ligation of p75NTR by NGF or pro-NGF can result in activation of caspase-3. 11 Previously, it has been shown, using Western blotting techniques, that when p75NTR is transfected into HEK293 cells and subsequently stimulated by with NGF, cleavage of caspase-3 resulted. 5 It should be emphasized that assays reported to date for measuring p75NTR activation are low-throughput and not amenable for screening of small-molecule inhibitors. In this work, we set out to develop a p75NTR receptor-signaling assay that was homogeneous and sufficiently robust to permit screening of small-molecule p75NTR modulators. We reasoned that increased caspase-3 activity, arising from p75NTR ligation, could directly be assayed by measuring cleavage mediated by caspase-3. A commercially available caspase-3 substrate was employed that is nonfluorescent in its uncleaved state, but becomes highly fluorescent after cleavage by caspase-3. In this work, we successfully (1) demonstrated that HEK293 cells transfected with p75NTR and stimulated with NGF show increased caspase-3 activity, (2) defined optimal experimental conditions for measuring the increase in caspase-3 activity in these cells, (3) validated that the increase in caspase-3 was p75NTR-mediated using a signaling-incompetent p75NTR mutant, and (4) utilized the assay to conduct a small-molecule pilot screen.
The development of a p75NTR-signaling assay in a tractable cell system such as HEK293 cells should greatly facilitate future dissection of the signaling events downstream from the p75NTR receptor. Many of the established methods to study p75NTR signaling have to date utilized primary neurons, which are more biologically relevant, but that require greater time and effort to prepare. For instance, one of the well-characterized model systems typically used for analysis of p75NTR-mediated apoptotic signaling employs sympathetic neurons from the superior cervical ganglion. 29,30 As demonstrated by K. Teng and others, 29 –31 treatment of rat primary superior cervical ganglion cell cultures with NGF suppresses apoptosis via a p75NTR-mediated mechanism, while treatment with the pro-neurotrophin increases apoptosis. It has been demonstrated that the kinase JNK appeared to mediate the pro-apoptotic effects. 30 The p75NTR/caspases-3 assay that we have described here provides a good model system to follow-up on these studies and further explores the signaling pathways triggered by neurotrophins and pro-neurotrophins. Future studies could focus on whether neurotrophins and pro-neurotrophins trigger independent signaling events in HEK293 cells (as they appear to induce in many primary neuronal systems).
In addition to having the advantage of utilizing the easily cultured HEK 293 cell line, the p75NTR/caspases-3 assay described here has several other advantages in comparison to many previously described p75NTR assays. For instance, the assay demonstrates an equally good or better signal to noise the ratio compared to other p75- signaling assays that are performed in primary culture systems. Many of these assays demonstrate a signal window less than twofold, 30,32 while the final optimized assay conditions for the p75NTR/caspases-3 assay described here demonstrated a signal change of twofold over background (Fig. 6). In addition, the assay described here also demonstrated very strong reproducibility; a nearly identical increase in caspase-3 activity was consistently observed with the optimized assay conditions on at least 10 different experimental occasions over many weeks (data not shown). Furthermore, compound 1 and the JNK inhibitor SP600125 yielded very consistent IC50 values on a number of test occasions. From our pilot screen, we observed 13 hits that decreased pro-NGF-mediated caspase-3 activation at 5 SDs from the mean (7.7% hit rate). Many of these 13 compounds were of a similar scaffold, which accounted for the high hit rate at this stringency. Also, compounds did not exhibit cytotoxicity. The relatively low screening concentration of 1 μM was chosen to minimize the potential for cytotoxicity in this assay. In addition, the compounds chosen for follow-up did not exhibit activity in other caspase-3 activation paradigms such as preventing epoxomicin- or staurosporine-induced caspase-3 activation. Thus, these compounds were specific for blocking p75-mediated caspase-3 activation. Overall, we have found this assay to be highly enabling in supporting hit to lead and lead-optimization screening efforts, where the Z′ values are sufficient for driving traditional medicinal chemistry SAR optimization. In contrast, a lead-optimization campaign using primary superior cervical ganglion cells (p75-expressing neurons) would be untenable due to the technical challenges of working with these specific primary neurons.
Experiments aimed at optimizing the p75NTR/caspase-3 activation assay revealed that several aspects of the procedure were important for facilitating activation of caspase-3 by neurotrophins in p75NTR-transfected cells. Our attention focused on two aspects of the procedure in particular: the effect of cell density and the presence/absence of serum. First, it was found that NGF stimulation of p75NTR- transfected cells caused caspase-3 activation when cells were plated at a density of 2,000 cells/well, but not when cells were plated at a density of 6,000 cells/well (Fig. 4C). Second, caspase-3 activation was only observed after neurotrophin stimulation if cells were deprived of serum during the neurotrophin incubation period (Fig. 4D). Together, these two findings suggest that p75NTR signaling in HEK293 cells is most robust when cells are under mildly stressful conditions. Our observations are also consistent with other literature findings indicating that p75NTR function is upregulated under conditions of stress. 33 –36 Specifically, it has been found that NGF binding to p75NTR induces nuclear translocation of NFκB in a cell line overexpressing p75NTR only after the cells have been subjected to the previous stress of serum withdrawal. 33 Serum withdrawal has also been demonstrated to be key for p75NTR-mediated apoptotic signaling in R2 cells. 35 It has also been previously demonstrated that cell density plays an important role in regulating p75NTR function, 36 which is consistent with our results.
Important data validating our p75NTR/caspase-3 activation assay was acquired using a signaling-incompetent p75NTR point mutant (Figs. 5, 6). The point mutation was introduced at residue Cys 256 in human p75NTR (corresponding to residue Cys 257 in rat p75NTR). Biochemical studies have shown that this cysteine residue is located within the transmembrane domain of p75NTR and appears to mediate the formation of constitutive, disulphide-linked p75NTR dimers. 5,37 Specifically, it has been observed that p75NTR can run at a molecular weight consistent with a dimer on nonreducing gels, but runs as a monomer on reducing gels. 5 Mutation of Cys 256 to Ala has been demonstrated to hinder the functionality of p75NTR dimers by blocking downstream signaling events but not blocking binding of p75NTR to neurotrophins. These studies led to the proposal of a snail-tong model for p75NTR receptor activation, whereby neurotrophin binding induces a conformational change in preformed p75NTR dimers. 5
The p75NTR/caspase-3 activation assay described here may be most useful for discovering and characterizing small-molecule modulators of p75NTR. Specifically, our assay has been designed to contain all the attributes desired for a cell-based, small-molecule screening: homogeneousness, use of an easily cultured and manipulated cell line, and compatibility with 96-well and 384-well plates. Discovery of compounds that directly bind p75NTR would appear to be feasible since several recent articles report identifying small-molecule modulators of p75NTR. 21,22 For instance, molecules based on loop 1 NGF have been synthesized and utilized to investigate p75NTR signaling and neuronal survival. 21 These molecules have been reported to promote survival of hippocampal neurons as measured by GAP-43 immunostaining and also using terminal deoxynucleotidyl transferase dUTP nick end labeling staining. 21 The compounds also have been reported to induce signaling events mediated by p75NTR in a manner that is comparable to neurotrophins such as NGF and brain-derived neurotrophic factor. Furthermore, the same compounds have been described to antagonize many of the detrimental effects of Aβ on dissociated hippocampal cultures and organotypic hippocampal slice cultures, in a manner that is presumably mediated by p75NTR. 22 In addition to the compounds described above, a small molecule has been reported to block the interaction of p75NTR with NGF and may modulate p75NTR signaling through caspase-3. 38 Another inhibitor was identified through in silico drug design and has been reported to have an inhibitory effect on NGF in an adhesion assay. 39 Hence, there now appears to be proof of concept for the discovery of p75NTR modulators.
In the p75NTR field, there has been an absence of assays that have the throughput to drive a traditional medicinal chemistry SAR effort. The caspase-3 activation assay described here has the throughput, reproducibility, and sensitivity to make data-driven decisions in the hit to lead and lead-optimization phases of a drug discovery program. IC50 values were highly reproducible from different scaffolds, including a JNK inhibitor and compound 1. Furthermore, we also demonstrated that the assay is useful for the screening of smaller focused libraries. Hence, this assay would be highly enabling for a p75NTR drug discovery program, where the alternative of working with primary neurons (dorsal root ganglion neurons) will not allow for the throughput to drive a program in the hit to lead or lead-optimization phases. The implementation of this assay will hence allow for the opportunity to identify novel scaffolds that would not be identified using other assays.
Conclusions
The p75NTR has long been recognized to regulate many cellular processes, including neuronal survival and apoptosis. 6,7 p75NTR has also been strongly linked to diseases of neurodegeneration, including Alzheimer's disease. 16,17 In particular, p75NTR expression has been localized to brain regions associated with Alzheimer's disease, and p75NTR has been functionally linked with signaling via Aβ peptides. The association between p75NTR and Alzheimer's disease has led to great interest in better understanding p75NTR-signaling mechanisms and identifying small molecules that might modulate p75NTR function. In this work, we have described an advance toward targeting p75NTR with the development of a novel protocol for a homogeneous p75NTR cellular assay that is amenable to small-molecule screening. This assay can now be applied to further study the signaling network triggered by p75NTR ligation and to further identify compounds that might regulate p75NTR for therapeutic benefit in Alzheimer's disease.
Footnotes
Acknowledgments
The authors gratefully thank the numerous members of the Elan community who played a role in this work.
Disclosure Statement
All authors were affiliated with Elan Pharmaceuticals, a biopharmaceutical company.