
Abstract
The KCNQ2/3 channel has emerged as a drug target for a number of neurological disorders including pain and epilepsy. Known KCNQ2/3 openers have effects on two distinct biophysical properties of the channel: (1) a hyperpolarizing shift in the voltage dependence of channel activation (V1/2), and (2) an increase in channel open probability or peak whole-cell current. The current high-throughput screening assays for KCNQ2/3 openers measure changes of channel activity at sub-peak conductances and the output measure is a combination of effects on V1/2 shift and peak current. Here, we describe a medium-throughput electrophysiological assay for screening KCNQ2/3 openers using the QPatch HT platform. We employed a double-pulse protocol that measures the shift in V1/2 and the change in current amplitude at peak conductance voltage. Retigabine along with novel KCNQ2/3 openers were evaluated in this assay. Three classes of KCNQ2/3 openers were identified based on the hyperpolarizing shift in V1/2 and the change in peak current. All three classes of compounds caused a hyperpolarizing shift in V1/2, but they were differentiated by their respective effects on peak current amplitude (increase, decrease, or only modestly affecting peak current amplitude). KCNQ2/3 blockers were also identified with this assay. These compounds blocked currents without affecting voltage-dependent activation. In summary, we have developed a medium-throughput assay that can reliably detect changes in the biophysical properties of the KCNQ2/3 channel, V1/2, and peak current amplitude, and therefore may serve as a reliable assay to evaluate KCNQ2/3 openers and blockers.
Introduction
Known KCNQ channel openers have two biophysically distinct modes of action on the channel, a hyperpolarizing shift in V1/2 and an increase in channel open probability, or peak current amplitude. 11 –16 However, openers may vary in their relative effects on these two modes of action. For example, retigabine causes a large hyperpolarizing shift in V1/2 but has a modest effect on peak current amplitude on KCNQ2 homomeric channels; in contrast, zinc pyrithione causes a large increase in peak current but only a small hyperpolarizing shift in V1/2. 17 The current high-throughput assays that are used to screen for KCNQ2/3 openers like thallium flux 18,19 are unable to detect changes in V1/2. The assay 18,19 is designed to best detect changes in flux at or near the activation threshold voltage. It is sensitive to compounds that cause a hyperpolarizing shift in V1/2 but has weak sensitivity for identifying compounds that only increase peak current amplitude. The medium-throughput IonWorks electrophysiological assay for KCNQ2/3 openers, 20 though capable of measuring V1/2, has not been reported to measure V1/2. The electrophysiological assay 20 measures changes in current amplitude at predetermined voltages. The changes in current reflect a combination of shifts in V1/2 and/or alterations in the open probability of the channel. Thus, as configured, these assays have demonstrated limited utility to assess structure-activity relationships of KCNQ2/3 openers. Therefore, to accurately evaluate the activity of KCNQ2/3 openers, both a hyperpolarizing shift in V1/2 and changes in peak current amplitude must be measured.
The QPatch HT (Sophion Bioscience A/S) is an automated, high-resolution whole-cell patch clamp system capable of simultaneous and independent recording of up to 48 cells. 21,22 It facilitates gigaseal resistances and therefore produces high quality recordings that enable investigation of kinetics for voltage-gated ion channels. 22 In this study, we describe the development of a QPatch assay to examine the properties of KCNQ2/3 channel openers. The assay was validated with reference compounds and novel Abbott KCNQ2/3 openers.
Materials and Methods
Reagents
Retigabine was synthesized in house according to published methods, 23 and Abbott KCNQ2/3 channel openers/blockers were also synthesized in-house. Flupirtine, zinc pyrithione, diclofenac, meclofenamic acid, and XE991 were purchased from Sigma.
Transfection and Cell Culture
The full-length coding sequences of human KCNQ2 and KCNQ3 were cloned into the pcDNA3.1/V5-his TOPO and pcDNA3.1/Zeo(+) expression vectors (Invitrogen) respectively, as described previously. 24 Human embryonic kidney (HEK) 293 cells were cultured in Dulbecco's modified Eagle's medium (Invitrogen) supplemented with 10% fetal bovine serum (Invitrogen) and 1% penicillin-streptomycin (Invitrogen) in a humidified 5% CO2, 95% O2 incubator at 37°C. The generation of HEK293 cells stably expressing heteromeric KCNQ2/3 channel (KCNQ2/3 cells) was described previously. 24 Briefly, equal amounts of the expression construct for KCNQ2 and KCNQ3 were transfected into HEK293 cells using LipofectAMINE 2000 (Invitrogen) according to the manufacturer's instructions. The stable cell line was then generated by antibiotic selection (0.2 mg/mL zeocin and 0.5 mg/mL geneticin, Invitrogen).
Cell Preparations
KCNQ2/3 cells grew in T225 flasks and were maintained at <80% confluence. For conventional patch-clamp recordings, cells were plated on poly-D-lysine coated coverslips the day before experiments. On the day of QPatch experiments, cells were rinsed twice with Ca2+ and Mg2+ free phosphate-buffered saline (PBS), followed by addition of 5 mL Detachin (Genlantis), and incubated at 37°C for 90 s unless otherwise noted. At the end of incubation, 5 mL of serum containing culture medium was added and cells were gently triturated. Cells were then centrifuged in a Beckman G5-15 centrifuge (Brea, CA) at 1000 rpm for 1 min, the supernatant was removed, and the cell pellet was resuspended with serum-free medium (SFM II; Invitrogen) supplemented with 20 mM HEPES at a density of 3,000,000–5,000,000 cells/mL. Cells were transferred to the QPatch stirring station, and were allowed to recover for 5 min before the start of the experiment.
Electrophysiology
QPatch HT recordings
All experiments were conducted on single-hole QPatch HT plates in the unattended mode of operation. Briefly, cells were stored in the cell hotel and constantly stirred. After initiation of the experiment, a series of procedures were conducted that included sealing, whole-cell formation, liquid application, recording, and data acquisition. KCNQ2/3 currents were recorded with a double-pulse protocol that consisted of a tail protocol followed by a step protocol. The tail protocol contained step pulses at 10 mV intervals from −100 to +60 mV for 1 s then returned to −30 mV for 200 ms. The step protocol contained a step pulse to +20 mV from a holding potential of −100 mV for 1 s. A wash step was added at the end of the protocol. Cells that showed greater than 10%–15% reduction of currents after wash were discarded. Rundown was not corrected in the results. Extracellular solution contained (in mM) 140 NaCl, 5 KCl, 2 CaCl2, 1 MgCl2, 10 HEPES, and 5 glucose (pH adjusted to 7.4 with NaOH). Internal solution contained (in mM) 125 K-aspartate, 20 KCl, 10 EGTA, 1 MgCl2, 5 Mg-ATP, and 5 HEPES (pH adjusted to 7.2 with KOH).
Conventional patch-clamp recordings
Whole-cell currents were recorded at room temperature using the standard patch-clamp technique with an Axopatch 200B amplifier (Molecular Devices). External and internal recording solutions were identical to those used on QPatch. Pipette electrodes had a typical resistance of 2–3 MΩ when filled with the internal solution. KCNQ2/3 currents, filtered at 5 kHz using the built-in filter of the amplifier, were recorded with the identical protocol to that used for the QPatch assay.
Compound preparation and handling
All compounds were made at 10 mM stock in dimethyl sulfoxide (DMSO). The highest testing concentration was 10 μM, which contained a maximum of 0.1% DMSO. For conventional patch clamp, compounds were applied using a DAD-12 Superfusion System (ALA Scientific Instruments) that uses a computer-controlled micromanifold with an outlet bore of 100 μm placed about 100 μm from the cells. For the QPatch experiments, compounds were held in glass inserts placed in 96-well plates. The compounds were delivered to cells as liquid applications in 4 brief bursts of 5 μL each.
Data Analysis
The QPatch data were analyzed using Sophion's QPatch Assay Software in combination with Origin 8.0 (OriginLab). The voltage-dependent activation curve was determined from peak tail currents and fitted with the Boltzmann function [1+exp([V1/2–V]/k)]−1, where V is the step pulse potential, V1/2 is the potential that gives half-maximal peak current, and k is the slope factor). Half activation voltage (V1/2) and slope factor (k) were then determined. The concentration response curves were fitted to a function Y=Bottom + (Top ™ Bottom)/(1+10^[(LogEC50−X) × HillSlope]), where Y is percentage change in current amplitude at +20 mV, X is concentration, and Bottom is fixed at 100. Average data are presented as mean±SEM.
Results
Dissociation Methods
One of the keys to generate reliable recordings in the QPatch assay is to ensure that isolated cells are viable. Typically, enzymes are used to achieve ideal isolation of cells, 22 but particular dissociation reagents limit expression levels of some ion channels. 25,26 Several dissociation methods with different enzymes were evaluated to determine the effects on the current amplitude of KCNQ2/3 channel. A standard protocol with Detachin (Method 1, Table 1) was first tested. KCNQ2/3 cells were incubated with Detachin for 5–10 min followed by gentle trituation with a pipette. This method yielded cells that were visually well isolated, and successful whole-cell recordings were obtained in 66% of wells. However, the current expression rate was low (67%, n=64) as was the current amplitude (320 pA, measured at +50 mV. 250 pA was the minimal current amplitude to score as positive). Next, using the same procedure as Method 1, Detachin was diluted with PBS by a ratio of 1:1 (Method 2, Table 1) to determine whether this could improve current amplitude. Similar results to Method 1 were observed with no increase in current amplitude. Further dilution of Detachin yielded less ideal dissociation without apparent improvement of current amplitude. In Method 3, the Detachin incubation time was shortened from 5 to 10 min to 1.5 min. At the end of enzyme incubation, an equal volume of serum-containing culture medium was immediately added into the incubation solution before gentle trituation (Table 1). This method yielded a good level of isolated single cells with only a few cell clusterings. The whole-cell formation was similar to previous methods at 69%. However, the current expression rate (84%, n=66) and the current amplitude (1,010 pA) improved significantly.
Comparison of Cell Dissociation, Whole-Cell Configuration Formation, and Current Amplitude Between Cell Dissociation Methods
An alternative enzyme, trypsin, was also examined. KCNQ2/3 cells were incubated with 0.125% trypsin for 1 to 2 min before trituation (Method 4, Table 1). This method yielded an ideal isolation of single cells and comparable whole-cell formation (83%), but current expression rate (44%, n=68) and current amplitude (270 pA) were much smaller than Method 3. To determine whether cell recovery from trypsin dissociation could improve current expression rate and amplitude, the cells were placed on a shaker in an incubator at 37°C for up to 8 h (Method 5) or overnight (Method 6) after dissociation with 0.125% trypsin. Method 5 produced no improvement over Method 4 in terms of current expression rate and amplitude. Method 6 yielded many cell clumps. Whole-cell formation was about 42%, however, the current expression rate was high (84%, n=40) and current amplitude was large (950 pA).
Thus, short incubation with Detachin (Method 3) yields acceptable dissociation of isolated cells and decent current amplitude, and this is the method that was used in this study.
Recording Protocols
KCNQ2/3 openers are known to have two biophysically distinct modes of action on KCNQ channels: a hyperpolarizing shift in V1/2 and an increase in channel open probability or current at peak conductance voltage. 11 –16 To address both of these modes of action, a double-pulse protocol was designed to evaluate channel openers (Fig. 1A). Cells were held at −100 mV and depolarized by step pulses at 10 mV intervals from −100 to +60 mV for 1 s and then returned to −30 mV for 200 ms, followed by a step pulse to +20 mV for 1 s. Current recordings corresponding to the protocols are shown in Figure 1B and C. The first part of the protocol, the tail protocol, was used to construct voltage-dependent activation curves by plotting the tail current amplitudes versus the conditioning voltages. The second part of the protocol, the step protocol, was used to construct concentration response curves by plotting the changes of current amplitude measured at +20 mV against 3 escalating concentrations.

QPatch voltage protocol and representative recordings of KCNQ2/3 currents.
Effects of Retigabine and Known KCNQ Openers and Blockers on KCNQ2/3 Channel
Retigabine was first tested on KCNQ2/3 cells with the double-pulse protocol (Fig. 2). Retigabine caused a hyperpolarizing shift in V1/2 and an increase in current amplitude in a concentration dependent manner. At 1, 10, and 100 μM, retigabine caused a shift in V1/2 of −5.8±2.1, −13.0±1.1, and −35.8±2.8 mV (control, V1/2: −11.3±1.6 mV, slope: −9.6±0.7; retigabine 0.1 μM, V1/2: −17.1±2.1 mV, slope: −9.1±0.5; 1 μM, V1/2: −24.3±2.7 mV, slope: −9.2±0.4; 10 μM, V1/2: −47.1±3.4 mV, slope: −9.5±0.9, n=4), respectively, (Fig. 2A–C). Retigabine's effects were reversible (data not shown). Application of 10 μM retigabine increased current amplitude 159±17% at +20 mV (Fig. 2D). The estimated EC50 was 1.31±0.14 μM (n=4; Fig. 2F). In addition, retigabine clearly slowed the deactivation of the tail currents (Fig. 2E). The results are generally consistent with a previous report using conventional patch clamp on recombinant KCNQ2/3 channels expressed in Chinese hamster ovary (CHO) cells. 27 The shift in V1/2 at 10 μM is also similar to that observed either in DRG neurons (−21 mV) 27 or sciatic nerve (−24 mV). 28 Other KCNQ openers flupirtine, zinc pyrithione, diclofenac, meclofenamic acid, and KCNQ blocker XE991 were also tested (n=3–6; Table 2). The IC50 for XE991 was 1.4 μM. The results are consistent with our unpublished data and previous reports using conventional patch clamp. 13,15,20
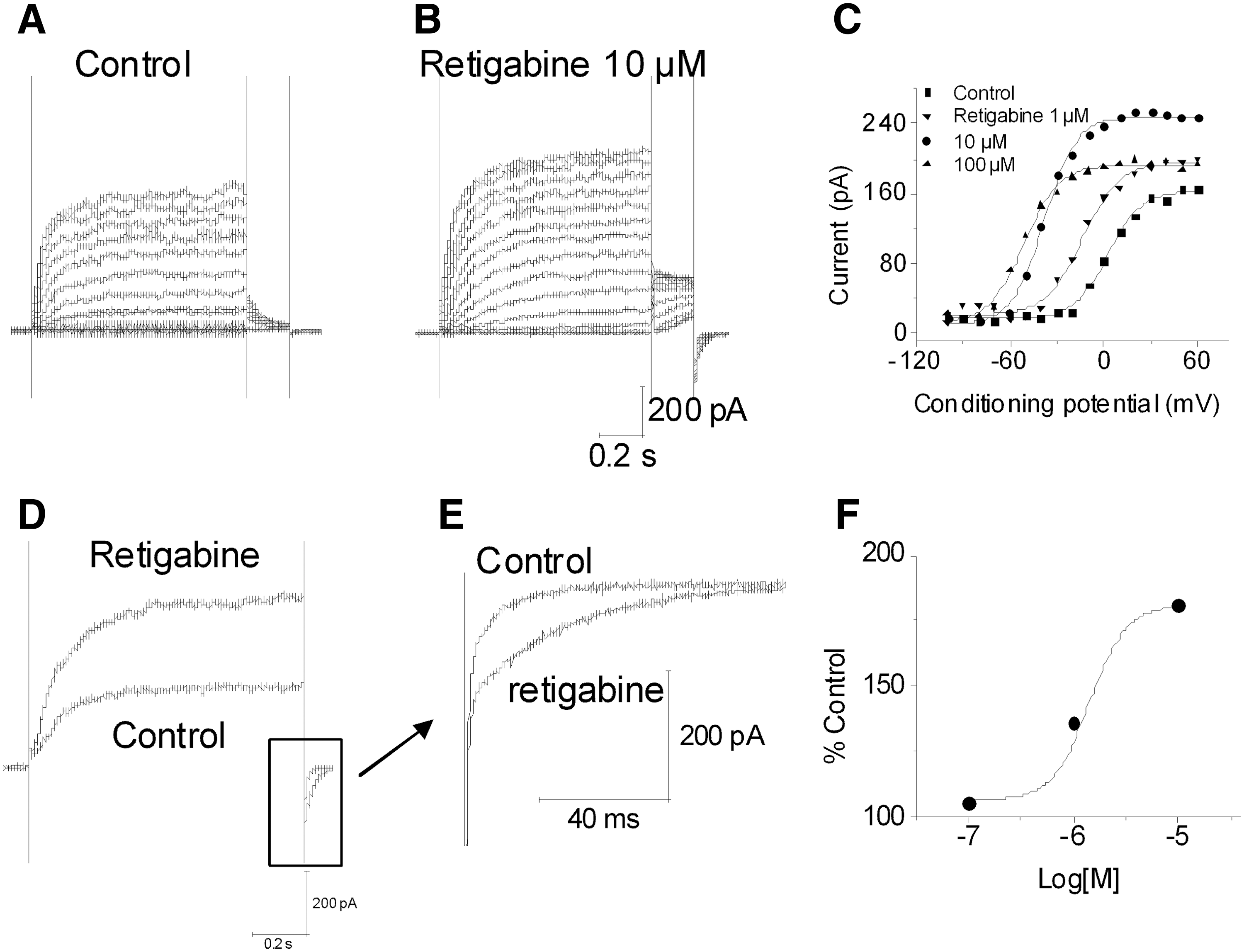
Retigabine enhances KCNQ2/3 currents. Shown is a representative cell in the absence
Effects of KCNQ Openers and Blockers on KCNQ2/3 Channel
ΔV1/2, hyperpolarizing shift in V1/2 (mV);% Current, maximum changes in current amplitude at +20 mV (% control); NV, not determined.
KCNQ2/3 Openers Show Different Profiles
In an effort to identify novel KCNQ2/3 channel openers, a high-throughput screening assay was conducted using a thallium flux assay. 19 A subset of the hits (∼10% representing multiple diverse chemical series) identified in this assay were further evaluated in a medium-throughput QPatch assay, which could screen ∼30 compounds/day for a three point concentration response curve with successful recordings on all 3 concentrations over 85% of the cells. Based on their effects on the hyperpolarizing shift in V1/2 and changes in peak current amplitude, KCNQ2/3 openers were classified into three classes (Table 3). The first class of compounds (n=173, examples are Compounds A–C in Table 3) was characterized by causing a hyperpolarizing shift in V1/2 and an increase in peak current amplitude (>159%–615%). The second class of compounds (n=57, examples are Compounds D–F in Table 3) was characterized by causing a hyperpolarizing shift in V1/2 and a decrease in peak current amplitude (49%–75%). The third class of compounds (n=27, examples are Compounds G–I in Table 3) was characterized by causing a hyperpolarizing shift in V1/2 but had minimal effects on peak current amplitude (93%–97%). In addition, 15 compounds (examples are Compounds J and K in Table 3) appeared to block KCNQ2/3 current by 50% or more at 10 μM at +20 mV.
Different Profiles of KCNQ2/3 Openers
Compound C, an example of the first class compound, showed a strong hyperpolarizing shift in V1/2 and an increase in current amplitude (Fig. 3A–C). At 0.1, 1, and 10 μM, Compound C caused a shift in V1/2 of −5.93±2.18, −18.74±2.79, and −50.38±5.72 mV (n=5), respectively. Compound C (10 μM) increased current amplitude 615±105% (n=5) at +20 mV (Fig. 3D). Compound C had an estimated EC50 of 0.27±0.09 μM (n=5). It also slowed the deactivation of the tail currents (Fig. 3D).
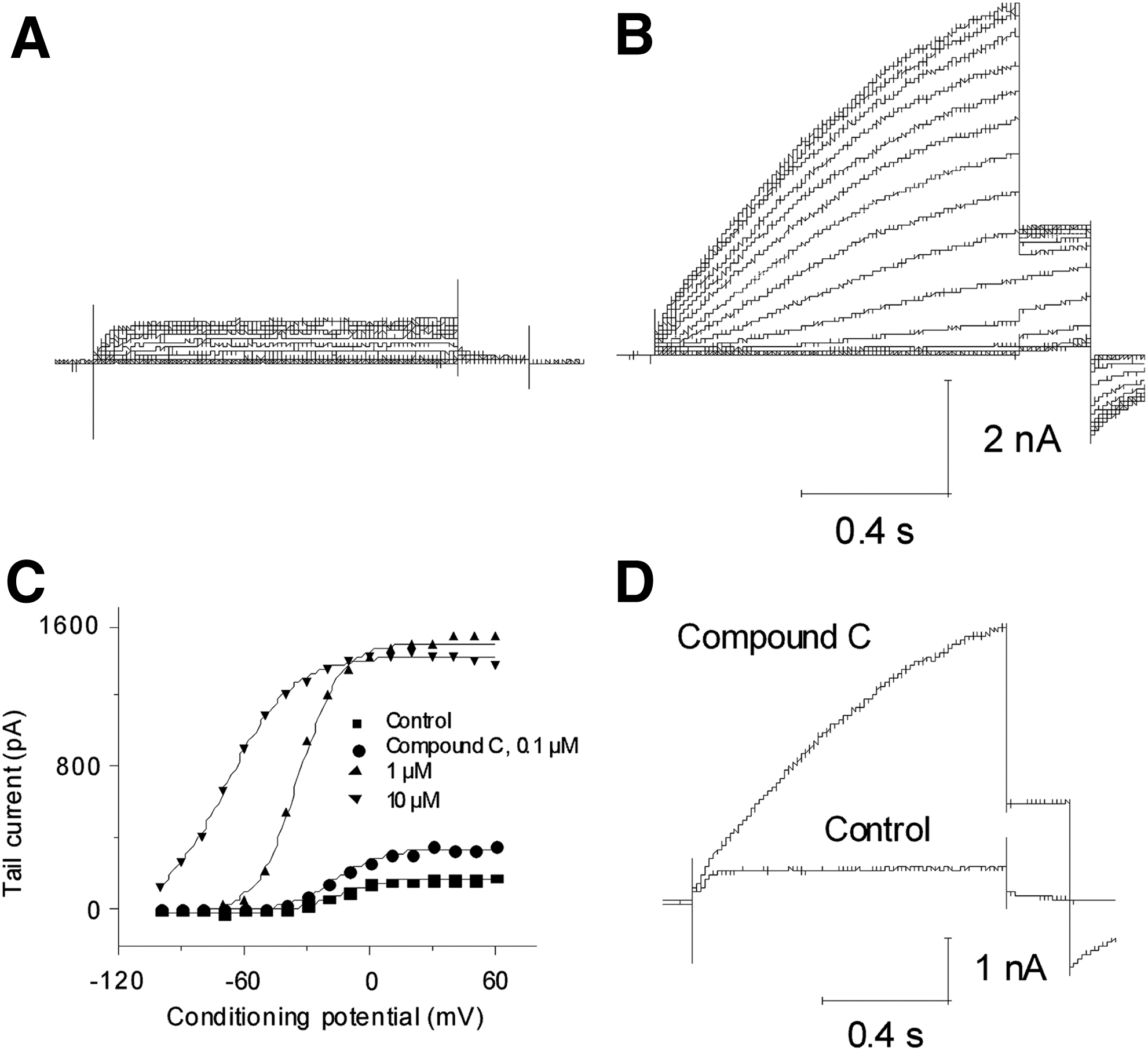
Properties of novel KCNQ2/3 opener Compound C. Shown is a representative cell in the absence
Compound D, an example of the second class compound, concentration dependently caused a hyperpolarizing shift in V1/2 but reduced current amplitude at higher concentrations when testing at +20 mV (Fig. 4A–C). At 0.1, 1, and 10 μM, Compound D caused a shift in V1/2 of −2.50±1.93, −15.70±3.63, and −32.23±3.16 mV (n=4), respectively. Application of 10 μM Compound D decreased current amplitude 49±10.4% (n=5) at +20 mV (Fig 4D). Like retigabine and Compound C, Compound D slowed the kinetics of the tail currents (Fig. 4E).

Properties of novel KCNQ2/3 opener Compound D. Shown is a representative cell in the absence
KCNQ2/3 blockers were also identified. Compound J blocked the current by 82.1±3.8% at 10 μM (n=4) at +20 mV with IC50 about 0.47 μM (Fig. 5A, B). At 0.1 μM, Compound J caused a −3 mV shift in V1/2 and a small change in slope (control, V1/2: −9.2±1.7 mV, slope, −9.1±0.4; compound J, V1/2: −11.9±1.2 mV, slope, −6.9±1.2, n=4), but the shift was not significant (P>0.05; Fig. 5C). The change in slope may reflect changes in the current components. When KCNQ2/3 currents were reduced, the proportion of the endogenously expressed Kv currents in the total currents likely increased. So, the V1/2 shifted to more depolarized potential. 28,29 Compound J did not slow the deactivation of the tail currents (Fig. 5D, E).
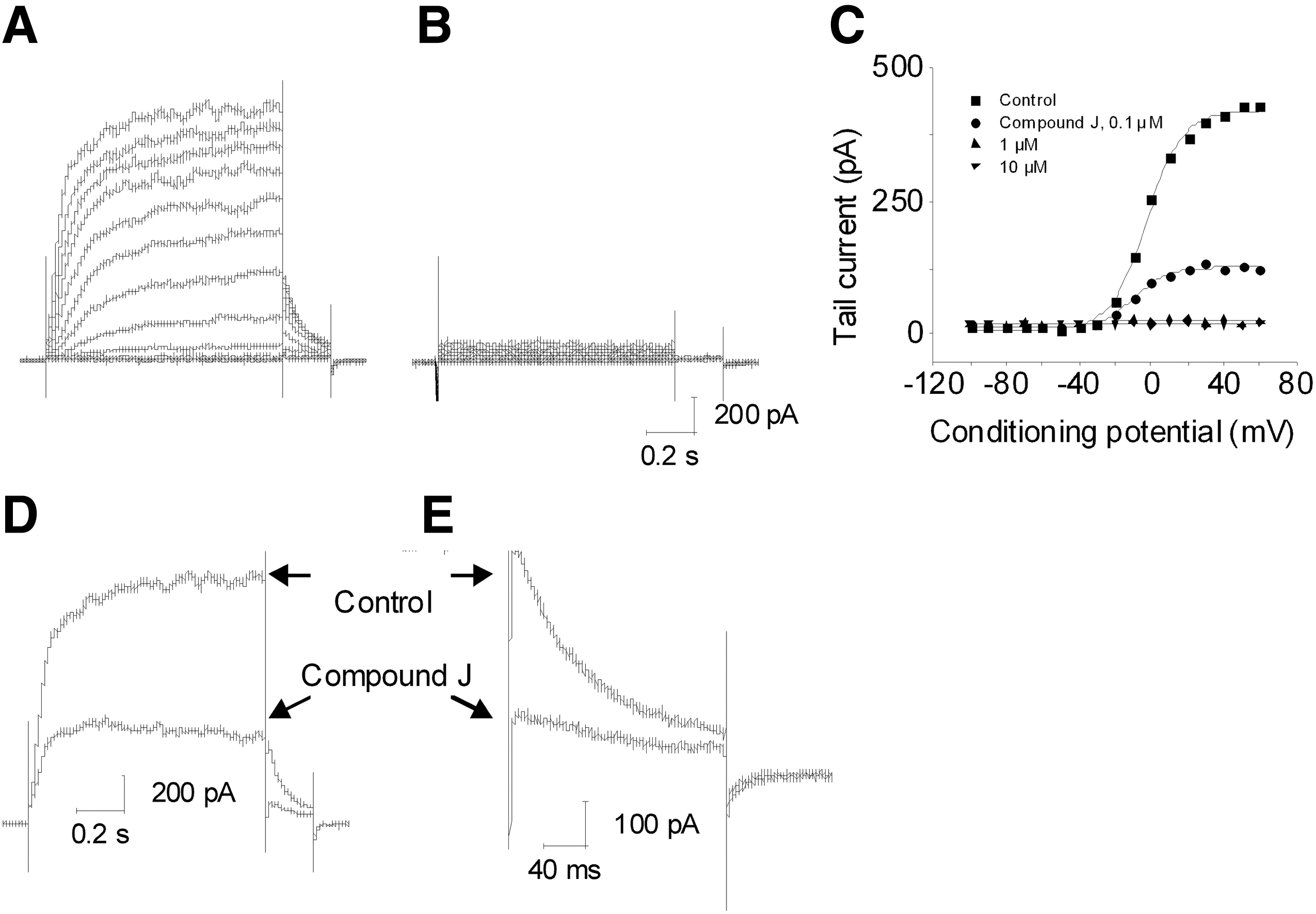
Properties of Novel KCNQ2/3 blocker Compound J. Shown is a representative cell in the absence
Correlation Between QPatch and the Conventional Patch-Clamp Recordings
A total of 14 compounds were tested (n=2–5) in both the conventional patch clamp and the QPatch assays using the same protocol (Fig. 1). As shown in Figure 6, the shift in V1/2 at 0.1, 1, and 10 μM correlated well between the two platforms (r 2=0.57, 0.76, and 0.87 at 0.1, 1, and 10 μM, respectively). The EC50 values (r 2=0.61) and changes in peak current amplitude (r 2=0.63) were also similar between the two assays.

Correlation between QPatch and the conventional patch clamp. The hyperpolarizing shifts in V1/2 at
Discussion
The QPatch HT platform can be applied as a medium-throughput patch clamp electrophysiological assay for evaluating KCNQ2/3 openers. Using a double-pulse protocol, two biophysically distinct modes of KCNQ2/3 channel activation were examined, a hyperpolarizing shift in V1/2 of voltage-dependent activation and changes in peak current amplitude. Three classes of KCNQ2/3 openers were identified based on changes in peak current amplitude. One class (Type I) increased peak current amplitude, another decreased this amplitude (Type II), and the third class only modestly affected peak current amplitude (Type III). All three classes of openers caused a hyperpolarizing shift in V1/2. KCNQ2/3 blockers were also identified with this assay. These compounds blocked currents without affecting voltage-dependent activation.
QPatch HT is a 48-well automated electrophysiology instrument that provides gigaseal whole-cell configuration enabling investigation of kinetics of voltage-gated ion channels and providing quality data that are similar to a conventional patch clamp. 21,22 In this study, we observed that the shift in V1/2, EC50, and percent increase in current amplitude values for KCNQ2/3 openers on QPatch correlate very well with those from the conventional patch clamp. This good correlation validates QPatch as a reliable higher throughput alternative to the conventional patch clamp for the evaluation of KCNQ2/3 openers.
The advantage of the QPatch assay is its ability to measure both the hyperpolarizing shift in V1/2 of voltage-dependent activation and the changes in peak current amplitude, which is clearly superior to the thallium flux assay 18,19 because the latter lacks the readout of hyperpolarizing shift in V1/2. KCNQ openers could increase 16,27 or decrease 12 peak current, however, the common feature of the openers is a hyperpolarizing shift in V1/2 and slow deactivation. 12,16,27 The properties of Compound C and Compound D support this notion. In fact, the hyperpolarizing shift in V1/2 correlates well with KCNQ2/3 openers' potency and selectivity observed in thallium flux assay. 19 Therefore, the data set that QPatch assay generated provides more valuable information for structure-activity relationships than the thallium flux assay.
Despite the advantages of the described QPatch protocol, there are still inherent experimental details that need to be scrutinized when conducting this assay. First, the current expression level is critical to the accurate measurement of V1/2. Most host cell lines, like HEK 293 cells, endogenously express Kv1.x, Kv3.x, and Kv4.x channels, 29 and the V1/2 of voltage-dependent activation for these channels is ∼3–21 mV. 29,30 The V1/2 of KCNQ2/3 channel is around −17 mV. 27 Since the recorded currents are the combination of KCNQ2/3 and endogenously expressed Kv currents, the V1/2 value would be significantly influenced by endogenous Kv channels if KCNQ2/3 expression is too low, and thus give an inaccurate readout on the effects of KCNQ2/3 openers. Second, when a compound strongly blocks the peak current, like Compound J, the V1/2 value may also be inaccurate since the contribution from Kv channels to the V1/2 value is significantly enhanced. Thus, the interpretation of compound-related changes to the V1/2 becomes difficult to interpret under these particular circumstances. Alternatively, CHO cells express little endogenous K currents 31 and have been used to express KCNQ2/3 currents. 17,27 Therefore, CHO cells are another option for the QPatch assay and may provide a better substrate for studying KCNQ2/3 channels.
Poor current expression level can hamper both throughput and the accurate assessment of compound effects. Improper cell dissociation methods can contribute to small current amplitudes. 25,26 Proteases can cause selective proteolysis within S5-pore extracellular linker of KCNH2 channel, which is structurally unique among KV channels, leading to small current amplitudes. 26 Shortening exposure to cell dissociation regents can improve current amplitudes. 25 Our dissociation method of using Detachin and limiting exposure time helped boost KCNQ2/3 current amplitudes to levels that are sufficient for QPatch study.
Based on the modes of action, three classes of KCNQ2/3 openers were identified. The first class of compound, represented by retigabine and Compound C, causes a hyperpolarizing shift in V1/2, increases the peak current amplitude, and slows deactivation. These properties resemble the majority of the known KCNQ2/3 openers. The second class of compound, represented by Compound D, causes hyperpolarizing shift in V1/2, slows deactivation, but it decreases the peak current amplitude. They represent a novel class of KCNQ2/3 openers. Retigabine has been known to dampen neuronal excitability by hyperpolarizing membrane potential and blocking action potential firing. It is effective in various animal models of epilepsy and chronic pain. 10 It is conceivable that the class of compounds with retigabine-like effects would be effective in animal models of pain. However, it remains to be seen what effects the second class of compound produces on neuronal excitability, and ultimately animal models of pain.
Footnotes
Author Disclosure Statement
All authors are employees of Abbott Laboratories.