
Abstract
In small-molecule/protein interaction studies, technical difficulties such as low solubility of small molecules or low abundance of protein samples often restrict the progress of research. Here, we describe a quartz-crystal microbalance (QCM) biosensor-based T7 phage display in combination use with a receptor–ligand contacts (RELIC) bioinformatics server for application in a plant Brz2001/DWARF4 system. Brz2001 is a brassinosteroid biosynthesis inhibitor in the less-soluble triazole series of compounds that targets DWARF4, a cytochrome P450 (Cyp450) monooxygenase containing heme and iron. Using a Brz2001 derivative that has higher solubility in 70% EtOH and forms a self-assembled monolayer on gold electrode, we selected 34 Brz2001-recognizing peptides from a 15-mer T7 phage–displayed random peptide library using a total of four sets of one-cycle biopanning. The RELIC/MOTIF program revealed continuous and discontinuous short motifs conserved within the 34 Brz2001-selected 15-mer peptide sequences, indicating the increase of information content for Brz2001 recognition. Furthermore, an analysis of similarity between the 34 peptides and the amino-acid sequence of DWARF4 using the RELIC/MATCH program generated a similarity plot and a cluster diagram of the amino-acid sequence. Both of these data highlighted an internally located disordered portion of a catalytic site on DWARF4, indicating that this portion is essential for Brz2001 recognition. A similar trend was also noted by an analysis using another 26 Brz2001-selected peptides, and not observed using the 27 gold electrode-recognizing control peptides, demonstrating the reproducibility and specificity of this method. Thus, this affinity-based strategy enables high-throughput detection of the small-molecule–recognizing portion on the target protein, which overcomes technical difficulties such as sample solubility or preparation that occur when conventional methods are used.
Introduction
A biosensor-based screening of T7 phage-displayed random peptide is a useful strategy for detecting small-molecule/protein interactions, which could circumvent those technical limitations. 2 –6 This approach employs a high-sensitivity quartz-crystal microbalance (QCM) biosensor that can monitor the decrease of alternating voltage–induced intrinsic crystal vibrations in real time, which indicates a mass increase in the gold electrode deposited on the crystal of the QCM sensor chip (Fig. 1A). The discovery of QCM phenomena in the air phase, which is described as the Sauerbrey's equation, goes back to the original report in 1959. 7 Owing to the reiterated physical and engineering refinements over the past half-century, high-resolution QCM biosensors have been developed. They make it possible to weigh the pico- to nanogram scales of bound molecules even in aqueous solutions, and have been widely applied to studies in life and material sciences. 8 –10 The use of this platform in a T7 phage display enabled a rapid capture of small-molecule recognizing T7 phages on the gold by sub-minutes of monitoring the frequency decrease, with no need of repeated rounds of biopanning. 3,4 This also corresponds to the fact that the QCM-based selection eliminated hitherto inevitable problems associated with phage distribution bias during the amplification process on classical methods. Furthermore, subsequent use of a host Escherichia coli culture realized a rapid and direct recovery of DNA from the trace amount of bound T7 phage, without elute condition exploration. By sequencing the peptide-encoding region of phage DNA, the corresponding amino-acid sequence can be clearly determined. In other words, the T7 phage particle plays a role as a transducer for the highly sensitive biosensor to sequence the trace amount of peptide bound to the small molecule on the gold electrode. Significantly, unlike proteins, the use of random shorter peptides can allow sterical contact with any compounds of small molecular size fixed on the gold electrode (Fig. 1A). 3,4
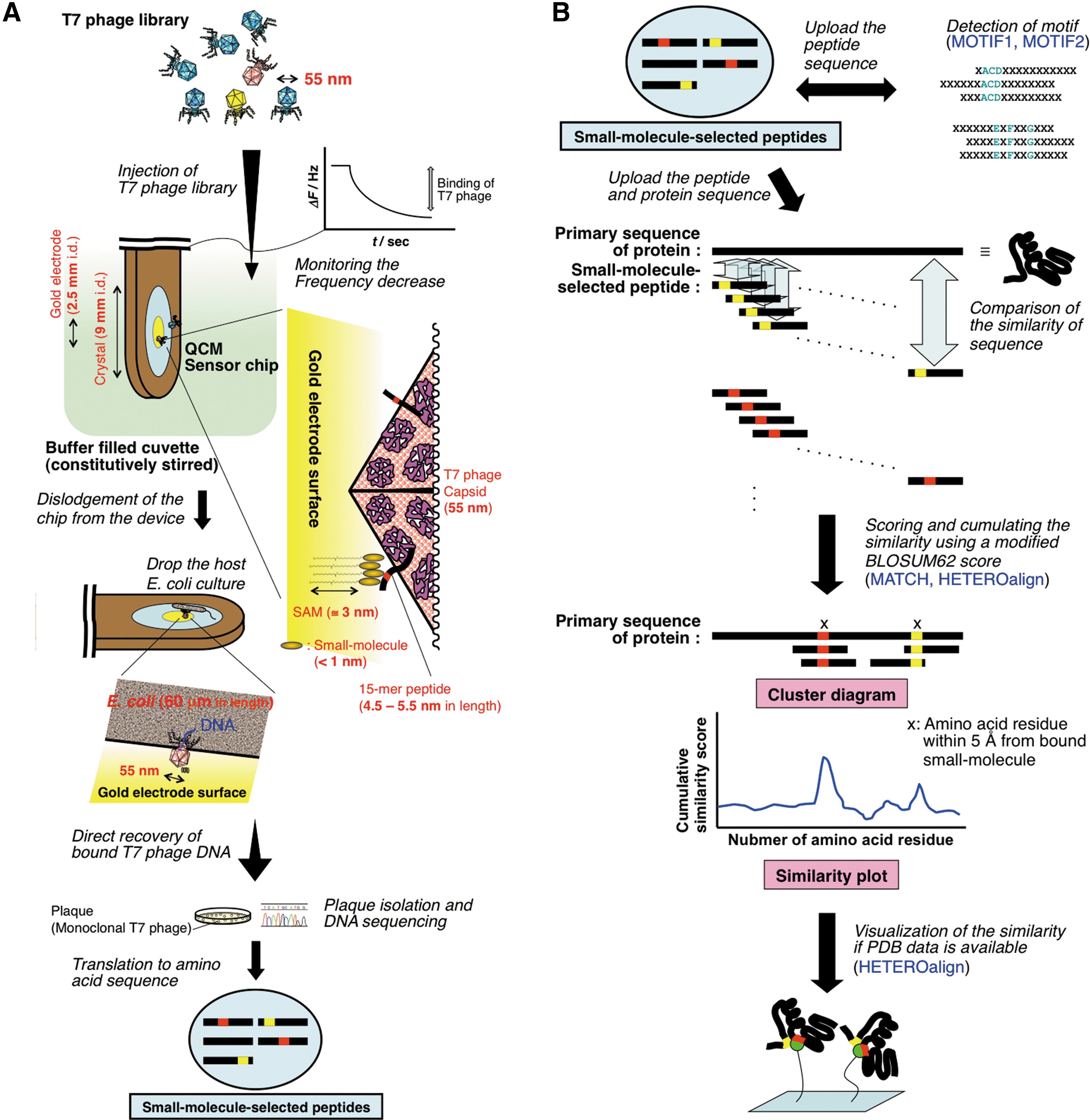
A schematic representation of a quartz-crystal microbalance (QCM) biosensor-based T7 phage display and an analysis of the small-molecule–selected peptide sequence using bioinformatics program in a receptor–ligand contacts (RELIC) suite.
In 2004, Rodi and Makowski and their coworkers launched the receptor–ligand contacts (RELIC) bioinformatics server, and they have since extensively studied the annotation of a small-molecule binding site on protein by a similarity search using affinity-selected random peptides. 11 –16 According to their earlier studies, phage-displayed short peptides, most of which are basically disordered, can mimic the disordered loops in naturally occurring proteins that are considered an essential portion in many small-molecule/protein interactions. 17 –19 Indeed, there are several examples of ligands that occupy sites on proteins which are not the classic “substrate binding site” as well as cases where analogous inhibitors do not bind analogously. 20 –22 Based on these observations, they have established a suite of bioinformatics programs designed for random peptide phage display. Once the amino-acid sequence of affinity-selected peptides is determined, the use of an appropriate program in RELIC with the sequence allows not only the qualitative and quantitative assessment of the affinity-selected peptide population, but also the annotation of small-molecule binding sites on protein that is dependent on the weak similarity with the limited short peptide sequence data (Fig. 1B). 11,13,23,24
In this work, the QCM-based one-cycle biopanning in combination use with RELIC bioinformatics analysis has been extended into plant molecular biology. 3,4 Brz2001 (Fig. 2A) is a specific DWARF4 inhibitor in plant hormone brassinosteroid biosynthesis. 25,26 Brz2001 belongs to the triazole series of compound that shows low solubility in solvents, as represented by antifungal agents such as fluconazole. In addition, recent studies have revealed that the target protein DWARF4 is a C22 hydrolase which is classified into plant cytochrome P450 (Cyp450) monooxygenase super family and contains heme and iron in an interior location of the enzyme where it catalyzes the hydroxylation reaction. 27 This fact implies that the DWARF4 protein requires these cofactors for folding into a functionally correct three-dimensional structure when genetically engineered. Furthermore, it may be also challenging to obtain the amount of purified plant enzyme for interaction analysis by NMR experiment or X-ray crystallography. Using this plant Brz2001/DWARF4 system, which may be time consuming and labor-intensive to refine the direct interaction properties on classical methods, we show that this rapid approach reveals disordered loops within catalytic sites on DWARF4 as an essential portion for Brz2001 recognition. This affinity-based strategy could be applicable for any small-molecule/protein systems under the identical protocol, overcoming the technical difficulties of conventional methods.
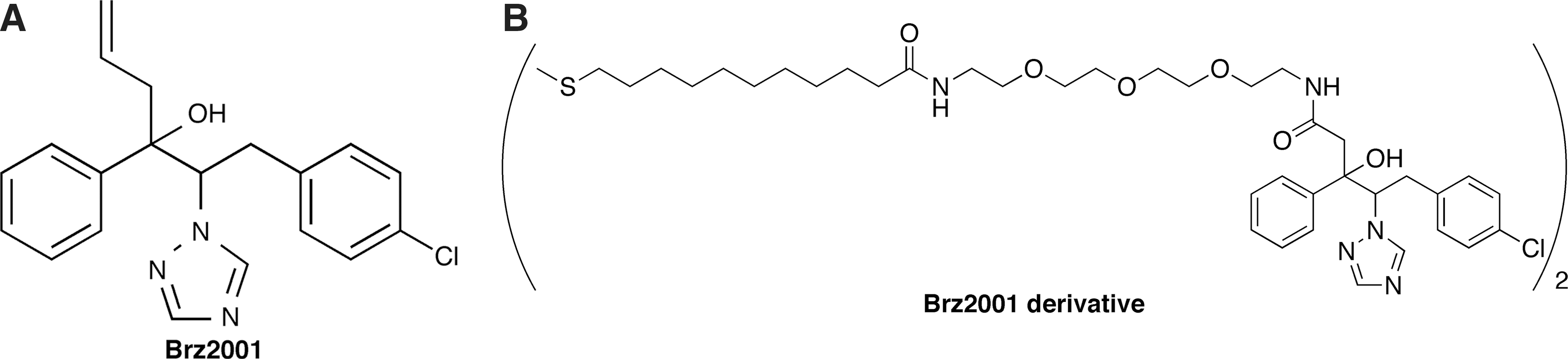
Structure of
Materials and Methods
QCM Apparatus and Reagents
The 27-MHz QCM apparatus (AffinixQ) and ceramic sensor chip (SiO2, 0.06 mm thick, 9 mm in diameter, 64 mm2; Au, 0.1 mm thick, 2.5 mm in diameter, 4.9 mm2) were purchased from Initium Inc. T7 select 10-3 OrientExpress™ cDNA Cloning System was from Novagen. The oligonucleotide and primer were purchased from Invitrogen™ Custom Oligonucleotides. Klenow DNA polymerase I and ExoSAP-IT were purchased from USB Corporation. Ex Taq DNA polymerase was obtained from TaKaRa. ABI PRISM® BigDye™ Terminator Cycle Sequencing Kit was from Applied Biosystems.
Construction of the T7 Phage-Displayed Random Peptide Library
For the preparation of a duplex DNA library, oligonucleotide GGG GAT CCG AAT TCT (NNK)15 TGA AAG CTT CTC GAG GG (0.056 pM) and CCC TCG AGA AGC TTT CA (0.56 pM) were mixed with Klenow buffer, heated to 95°C for 5 min, and annealed by slowly cooling the mixture to 37°C. The single-stranded regions were converted to duplex DNA by continuing the incubation at 37°C for 2 h in the presence of dNTPs (2.5 mM) and Klenow enzyme (0.5 mU/μL). After the reaction, double-stranded DNA was recovered by EtOH precipitation. The obtained DNA was then digested separately with EcoRI and HindIII restriction enzyme and inserted into the T7 select 10-3b vector according to the manufacturer's instructions. The primary titer of this T7 phage library was 1.6×107 pfu/mL. For the screening procedure, the phage library was amplified up to 1.7×1010 pfu/mL using E. coli (BLT5615) as the host strain.
The QCM Biosensor-Based T7 Phage Display
The protocol is summarized in Table 1 . A ceramic sensor chip was attached on the oscillator of a 27-MHz QCM apparatus (AffinixQ), and the intrinsic frequency in the air phase was recorded before compound immobilization. After detaching the chip, a 20 μL aliquot of a Brz2001 derivative solution (1 mM in 70% EtOH) was dropped onto the gold electrode of the ceramic sensor chip and left for 1 h under a humid and shaded atmosphere at room temperature. The surface of the electrode was washed for 10 min in buffer (10 mM Tris-HCl, pH 8.0, and 200 mM NaCl), which was stirred at 1,000 rpm at 25°C (AffinixQ, Initium Inc.). The sensor chip was set up for the QCM apparatus and recorded the decreased frequency in the air phase to measure the immobilized Brz2001 amount. The Brz2001 derivative was immobilized at ∼11,000 Hz (330 ng) on average. The QCM sensor was immersed into the cuvette containing 8 mL of buffer and then allowed to fully stabilize. An aliquot of 8 μL of a T7 phage library (1.7×1010 pfu/mL) was injected into the cuvette. Frequency changes, caused by binding to the Brz2001 immobilized on the gold electrode surface, were then monitored for 10 min. For the recovery of bound phages, 20 μL of log-phase host E. coli (BLT5615) solution, which was cultured for 30 min at 37°C in the presence of 50 μg/mL of carbenicillin and 1 mM of IPTG beforehand (OD600=1.0), was dropped onto the gold electrode and then incubated at 37°C for 30 min. Another 200 μL of LB medium was then added to the resulting solution, which was then subjected to plaque isolation. After that, the gold electrode surface was swabbed with 1% sodium dodecyl sulfate, treated with 5 μL of piranha solution (concentrated H2SO4: 30% H2O2=3: 1) for 5 min, washed with dH2O, and then dried.
Protocol for the QCM Biosensor-Based T7-Phage Display and Analysis of RELIC Bioinformatics Server
1. A 20 μL aliquot of a Brz2001 derivative solution (70% EtOH) that forms SAM was dropped onto the gold electrode of the ceramic sensor chip and left for 1 h under a humid and shaded atmosphere at room temperature. The immobilized amount on gold was measured as the decreased frequency in the air phase after immobilization.
2. The sensor chip was attached on oscillator and immersed; the buffer-filled cuvette was constitutively stirred at 1000 rpm at 25°C (AffinixQ, Initium Inc.).
3. After stabilizing the QCM sensor, the 8 μL of T7 phage library (final >108 pfu) was injected into the cuvette.
4. Frequency decrease by mass increase on binding of T7 phage to Brz2001 on the gold electrode was monitored for 10 min.
5. The sensor chip was detached from the apparatus, and 20 μL of log-phase and T7 phage-infective host Escherichia coli (BLT5615) solution was dropped onto the gold electrode to incubate for 30 min at 37°C with shaking. The resulting solution was added to 200 μL of LB medium.
6. The 100 μL of each diluted solution (101 to 106 times) was plated onto LB/carbenicillin plate with 3 mL of prewarmed top agarose and 100 μL of host E. coli solution, and incubated for 3 h at 37°C.
7. Each plaque was isolated and suspended into 50 μL phage extraction buffer. Peptide-encoding region on T7 phage DNA was amplified by PCR using specific primers and then sequenced.
8. Resulting amino-acid sequence of peptide was listed in .txt file and uploaded to execute an appropriate program in RELIC bioinformatics server according to the instructions.
9. After swabbing the gold electrode surface by 1% SDS, 5 μL of piranha solution (concentrated H2SO4: 30% H2O2=3: 1) was dropped and left for 5 min, and then washed by dH2O.
QCM, quartz-crystal microbalance; SAM, self-assembled monolayer; RELIC, receptor ligand contacts; SDS, sodium dodecyl sulfate; PCR, polymerase chain reaction.
Plaque Isolation
The recovered phage solution, which contains 5×105–5×106 pfu/mL of T7 phages on average, was diluted from 101 to 106 times. A 100 μL aliquot of each solution was mixed with 200 μL of log-phase host E. coli solution and 3 mL of prewarmed top agarose [10 g/L Bacto tryptone, 5 g/L yeast extract, 5 g/L NaCl, 6 g/L agarose], and then seeded onto an LB/carbenicillin plate. The plate was incubated at 37°C for 3 h to form individual phage plaque.
PCR and DNA Sequencing
Plaques were randomly picked from LB plates, and each was dissolved in 50 μL of phage extraction buffer (20 mM Tris–HCl, pH 8.0, 100 mM NaCl, 6 mM MgSO4). The phages were disrupted by heating the extract to 65°C for 10 min. To amplify the peptide-encoding region, PCR was performed according to the following PCR mixture: 0.3 μL phage solution, 0.05 μL of the 100 μM forward primer 5′-TGC TAA CTT CCA AGC GGA CC-3′ and the 100 μM reverse primer 5′-AAA AAC CCC TCA AGA CCC GTT TA-3′, 0.25 mM dNTP, 1 μL of 10× Ex Taq DNA polymerase buffer, and 1.25 U of Ex Taq DNA polymerase. The mixture was diluted up to 10 μL using double-distilled water. PCR condition was 25 cycles each of 94°C for 60 s, 50°C for 30 s, and 72°C for 30 s. The 5 μL of products were treated with 2 μL of ExoSAP-IT (digested at 37°C for 15 min and then inactivated at 80°C for 15 min) and 70% EtOH precipitated. The products were then sequenced using ABI Prism BigDye Terminator Cycle Sequencing Kit and on an ABI Prism3100 Genetic Analyzer (ABI) according to the manufacturer's protocols.
Analysis Using RELIC Server
The stand-alone type RELIC programs were kindly provided by Dr. Lee Makowski (Northeastern University, Boston, MA). AAFREQ, MOTIF1 and 2, and MATCH programs in RELIC were used according to their manuals for analysis of Brz2001-selected peptides and detection of the Brz2001-binding site on DWARF4. 13
Results and Discussion
To immobilize Brz2001 (MW: 353.85) on the gold electrode of the QCM sensor chip, we used a derivative that forms a self-assembled monolayer on gold (
Fig. 2B
, Supplementary Fig. S1; Supplementary Data are available online at
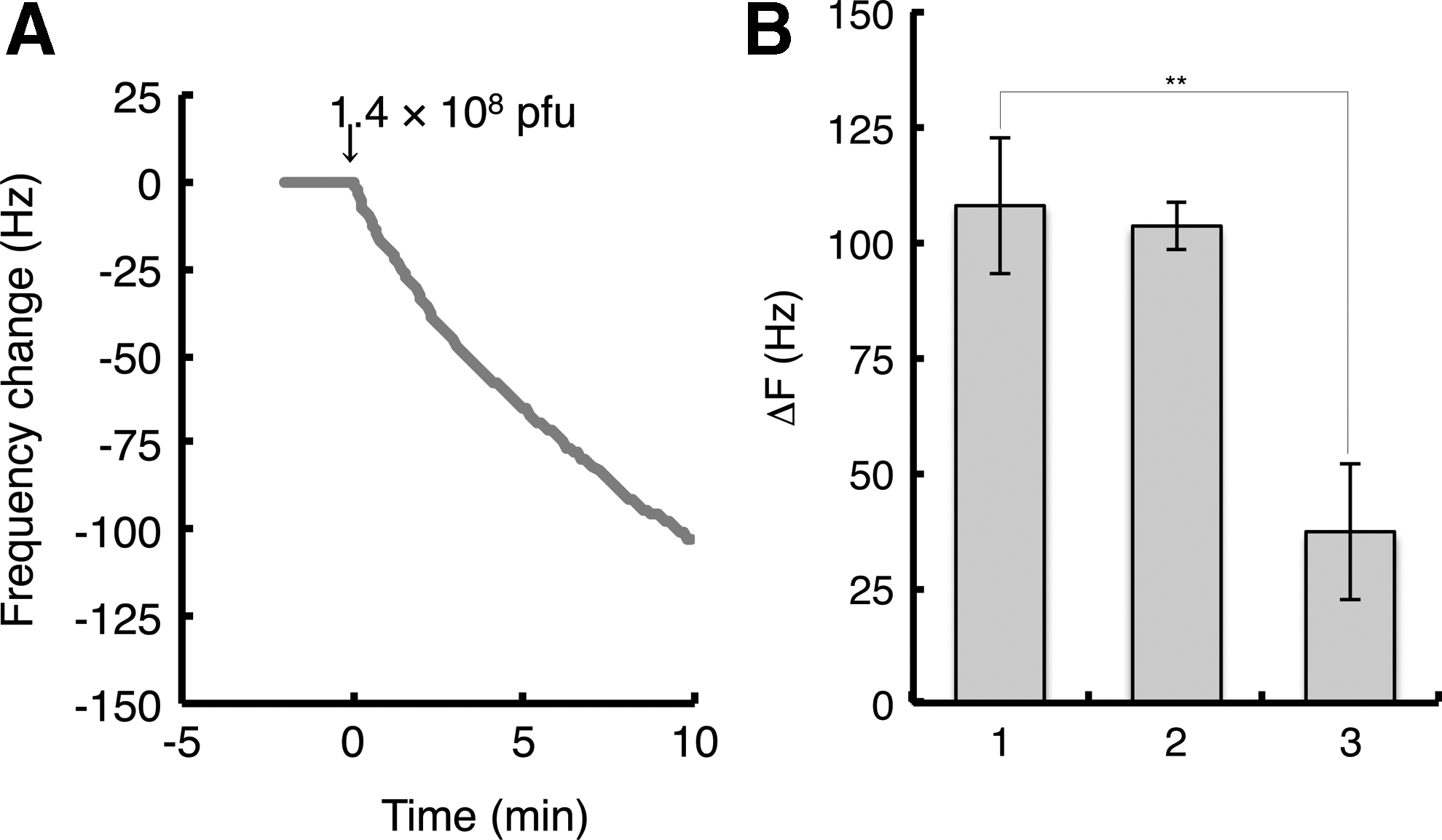
The QCM biosensor-based one-cycle biopanning.
Brz2001-Selected 15-mer Peptide Sequences After Four Sets of Individual One-Cycle Biopanning
T7-phage particles were arbitrarily extracted from the resulting solution and then analyzed by PCR amplification of the DNA encoding the fusion peptide. The PCR products were then sequenced. aThe peptide directly highlights a disordered portion on DWARF4 using MATCH program ( Fig. 5A ).
In general, small molecules appear to show weak interactions with peptides via surface–surface contacts of continuous or discontinuous motifs of several amino-acid residues. Testing the interaction between individual peptides with small molecules should be experimentally limited to some extent, because there may be a large number of potential small-molecule–recognizing peptides within a library. Furthermore, affinity between immobilized small molecules and short peptides is too weak (K D: 10−4–10−5 M range) to characterize each affinity status. Nonetheless, algorithmic and heuristic approaches using the RELIC program enabled rapid extraction of sequential information that is potentially associated with the small-molecule recognition from a subset of affinity-selected peptides, without checking the affinity status of individual peptides. 11,13,23,24 Table 3 shows continuous motifs that were detected from 34 Brz2001-selected peptides by the use of the RELIC/MOTIF1 program. In addition to the exact match of short amino-acid stretches, conservative motifs are efficiently extracted from the limited peptide population. Indeed, a pair of six-continuous motifs, 6 kinds of five-continuous and 25 of four-continuous motifs, were detected in the subset of peptides (Table 3). Furthermore, the RELIC/MOTIF2 program, which searches for patterns of three amino acids and does not allow conservative amino-acid substitutions, allows identical gap lengths, highlighted VXXFXF (among five individual peptides), CXXVXL, FXVXXG, SXXXVXXXXS, VXLXV (four peptides; Table 4), and others (two or three peptides; data not shown). These motifs were not significantly detected within the 103 peptide sequences (Supplementary Table S1) arbitrarily selected from the unscreened parent library. In addition, frequency of amino acids C F G S V, which are the components of the continuous and discontinuous motifs, also increased 1.1- to 1.23-fold through the selection, although it decreased or did not change in the other amino acids (Supplementary Tables S2 and S3). Collectively, these results indicate that the information content for Brz2001 recognition significantly increased through the affinity selection for Brz2001.
Four to Six Continuous Motifs Extracted from Brz2001-Affinity-Selected Peptides Using the MOTIF1 Program in the RELIC Suite
aThe peptide directly highlights a disordered portion on DWARF4 using MATCH program.
Three Discontinuous Motifs Extracted from Brz2001-Affinity-Selected Peptides Using the MOTIF2 Program in the RELIC Suite
Motifs conserving more than 4 peptides are presented in bold.
aThe peptide directly highlights a disordered portion on DWARF4 using MATCH program.
Although the mode of docking of Brz2001 to DWARF4 has still been unsolved by NMR experiment or X-ray crystallography, the subsequent use of the RELIC/MATCH program with the Brz2001-selected peptide sequences enabled the prediction of polypeptide portions involved in Brz2001 binding. This program calculates a weak similarity between small-molecule–selected short peptides and a longer target polypeptide, even in the absence of a clearly identifiable consensus sequence motif or a three-dimensional structure of the target ( Fig. 1B ). 3,4,11,13 Figure 4A shows a similarity plot between 34 Brz2001-selected random peptides and the DWARF4 amino-acid sequence (M1-L513). This plot was calculated by a modified BLOSAM62 amino-acid matrix within each window of five amino acids in length across the entire length of the protein sequence. 12,14 –16 The similarity was cumulatively scored and plotted at each amino-acid residue. Furthermore, scores calculated using 103 unscreened parent random peptides (Supplementary Table S1) were subtracted as a background. As a result, a portion containing S438 in DWARF4 was detected as a maximal similarity score by employing 34 Brz2001-selected peptide sequences. As seen in Figure 5A, this analysis also generated a cluster diagram, indicating that this portion is essential for Brz2001 recognition. A similar trend was noted in the analysis using another 26 Brz2001-selected peptides ( Figs. 4B and 5B; Supplementary Table S4). In this case, other portions containing N379, P407, and P454, respectively, were also pinpointed, in addition to S438. By contrast, such maximal scores were not observed by analysis using 27 gold electrode–recognizing control peptides ( Fig. 4C; Supplementary Table S5). These results indicate the reproducibility and specificity of this method. Furthermore, a part of the highlighted amino-acid residues in 15-mer peptides overlapped with the potential Brz2001-recognizing motifs detected in analysis using RELIC/MOTIF-1 and -2 programs ( Tables 3 and 4 ). Thus, this strategy could be applicable to annotate a small-molecule–recognizing portion on proteins in the case when the three-dimensional structure is unavailable. Significantly, the small-molecule binding site that is internally located on protein can be targeted by this approach. Furthermore, this method is free from both naturally and experimentally occurring protein mutations, deletions, and other modifications that may influence the protein folding and fail to target identification in some cases.
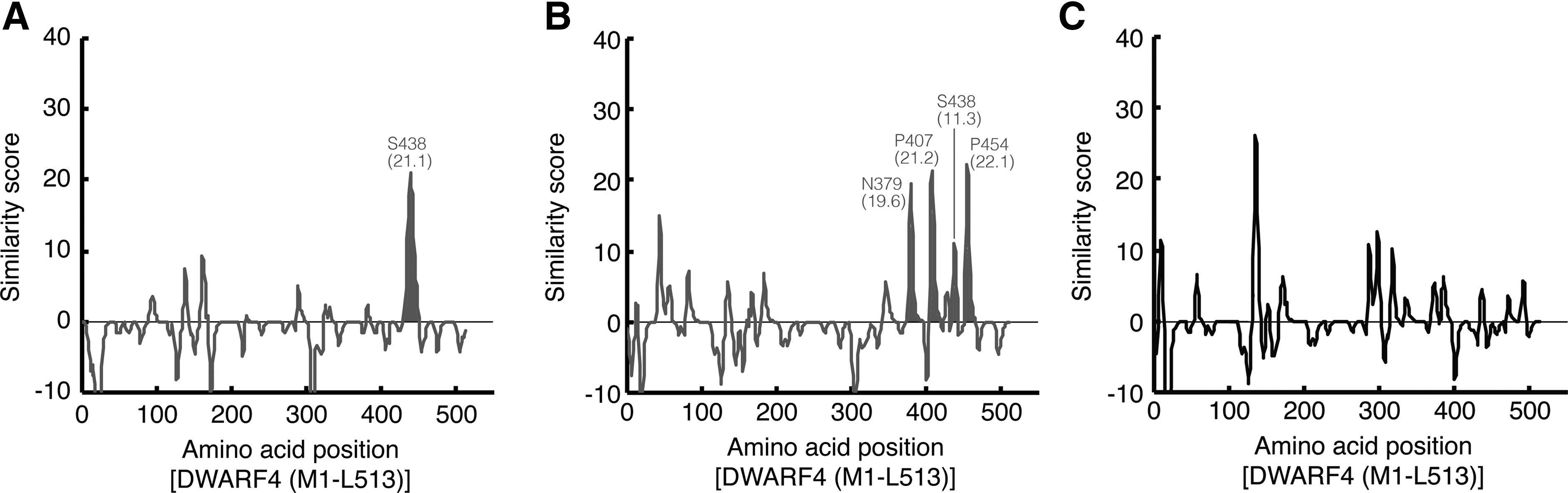
Analysis of the similarity of amino-acid sequence between Brz2001-selected peptides and DWARF4 using the MATCH program in the RELIC suite: similarity plot. Scores for the sequence of DWARF4 (M1-L513) against the sequences of 34
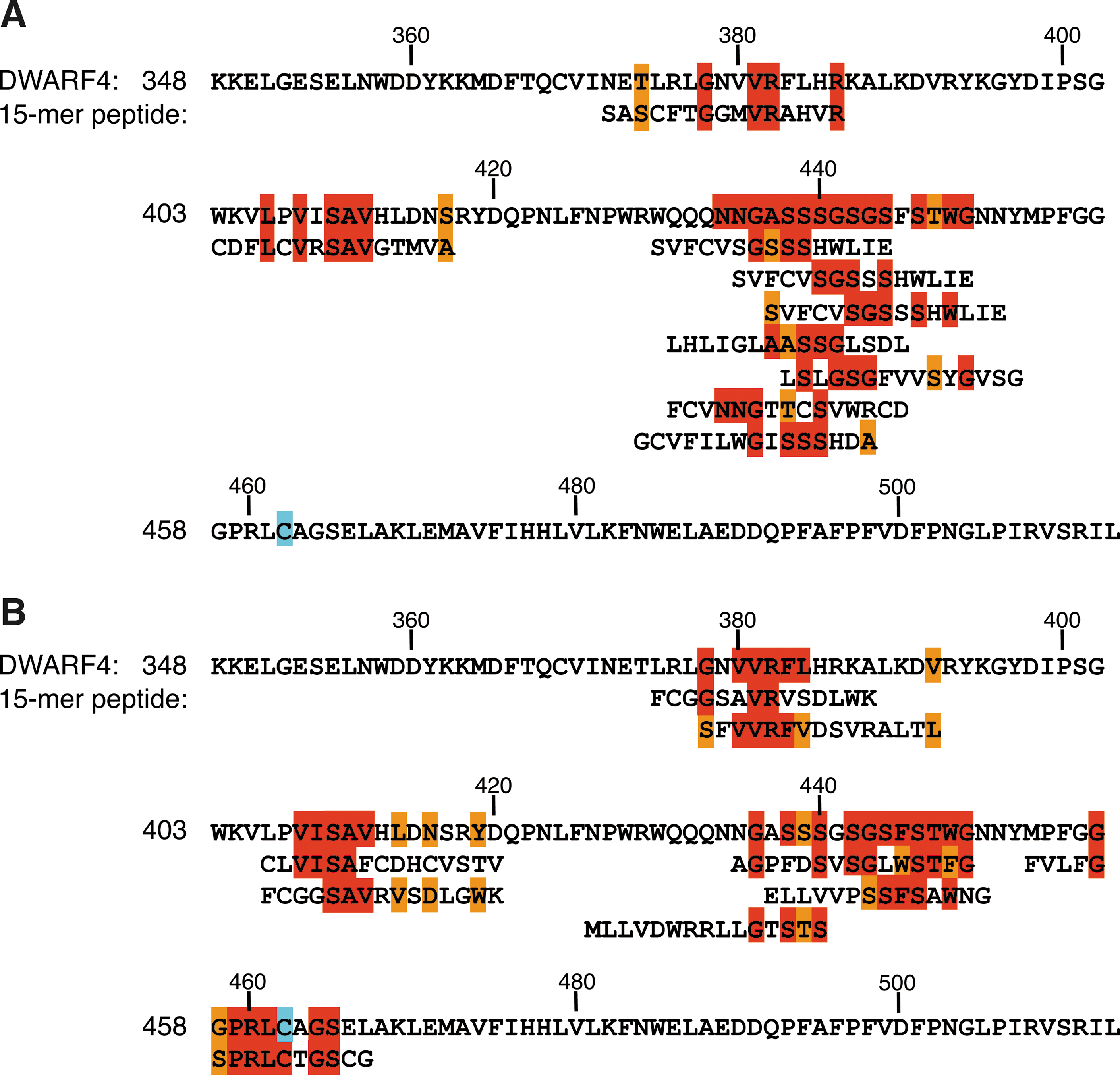
Analysis of the similarity of amino-acid sequence between Brz2001-selected peptides and DWARF4 using the MATCH program in the RELIC suite: cluster diagram. A highlighted mapping of amino-acid sequence between Brz2001-affinity-selected 34
According to the information about human cytochrome 2C9, one of a Cyp450 monooxygenase homologous to DWARF4 (E value: 6e-12 on standard blastp) and being available in the three dimensional structure [PDB ID: 1R9O], the primary sequence of the highlighted portion is potentially disordered flexible loops that comprise a substrate-binding site within the vicinity of the catalytic site that is mediated by heme, iron, and a cysteine of the heme axial ligand ( Fig. 5A, B; Supplementary Fig. S2). 34,35 Indeed, DWARF4 inhibitors having a triazole ring interfere with the redox activity via this portion, which result in the inhibition of brassinosteroid biosynthesis and growth suppression of plants ranging in concentration from 10−4 to 10−6 M (K D: 10−5–10−6 M). 25 –27,36,37 Dynamic mobilization of the flexible portions that have been first detected in this experiment seems to be essential during docking of Brz2001 to the catalytic site. In addition, several other amino-acid residues in DWARF4 were detected by this analysis ( Fig. 4A, B ). These residues could contribute to the binding of Brz2001to DWARF4, or might just be false positives, as can be seen in the control experiment ( Fig. 4C ). Conversely, undetected portions or residues involved in the binding of Brz2001 might be present. The introduction of the position of linker on Brz2001, or genetically modified diversity or length of library peptides, might improve these properties. Along with precise docking studies, further experiments make it possible to expand the utility of this approach.
Since brassinosteroid has been discovered as a plant growth–promoting hormone, extensive studies have been conducted about its biosynthesis as well as the production of inhibitors as a tool for biosynthesis regulation. 25 –27,36 –41 Brz2001 is one of the synthetic compounds that specifically inhibits DWARF4 activity. These findings in our work will greatly contribute to better understanding the precise mode of docking as well as further designing the DWARF4 inhibitor for studies in brassinosteroid biosynthesis. Moreover, experimental methodology in this work, which could be beyond the technical difficulties of the conventional approach, will be a valuable tool for detecting a small-molecule binding site on a target protein as well as expanding the druggable proteome in a wide range of small-molecule–related studies, including plant molecular biology.
Conclusion
In this work, the QCM biosensor-based T7 phage display in combination use with bioinformatics analysis has extended into plant molecular biology. A screening of a library of T7 phage-displayed 15-mer random peptides was undertaken to identify 34 Brz2001-recognizing peptides. Subsequent qualitative and quantitative assessment of affinity-selected peptides, which is based on an algorithmic and heuristic approach using RELIC programs, predicted the increase in the information content of Brz2001 recognition through the selection, highlighting continuous and discontinuous peptide motifs. Furthermore, a similarity search using the RELIC/MATCH program with Brz2001-selected peptides has identified a potential disordered portion of DWARF4 as a scaffold during Brz2001 docking, even though the three-dimensional structure of this enzyme has been unavailable. A similar trend was also noted by an analysis using another 26 Brz-selected peptides and not observed using 27 control peptides, demonstrating the reproducibility and specificity of this method. Beyond the technical limitations associated with physicochemical properties of small-molecule or protein samples, this affinity-based methodology could be applicable to any small-molecule/protein systems under the identical protocol as a high-throughput method for the scanning of small-molecule-recognizing portion. Experimental methodology in this work will greatly contribute to the wide range of small-molecule–related studies, including plant molecular biology.
Footnotes
Acknowledgments
The authors thank Dr. Tadao Asami (University of Tokyo, Tokyo, Japan) for providing Brz2001, and Dr. Lee Makowski (Northeastern University, Boston, MA) for providing the stand-alone type RELIC programs. This work was partially supported by a Grant-in-Aid for Scientific Research (The Ministry of Education, Culture, Sports, Science, and Technology of Japan, Japan Society for the Promotion of Science).
Disclosure Statement
The authors declare no conflict of interests.