
Abstract
Primary hyperoxaluria is a severe disease for which the best current therapy is dialysis or organ transplantation. These are risky, inconvenient, and costly procedures. In some patients, pyridoxine treatment can delay the need for these surgical procedures. The underlying cause of particular forms of this disease is the misrouting of a specific enzyme, alanine:glyoxylate aminotransferase (AGT), to the mitochondria instead of the peroxisomes. Pharmacoperones are small molecules that can rescue misfolded proteins and redirect them to their correct location, thereby restoring their function and potentially curing disease. In the present study, we miniaturized a cell-based assay to identify pharmacoperone drugs present in large chemical libraries to selectively correct AGT misrouting. This assay employs AGT-170, a mutant form of AGT that predominantly resides in the mitochondria, which we monitor for its relocation to the peroxisomes through automated image acquisition and analysis. Over the course of a pilot screen of 1,280 test compounds, we achieved an average Z′-factor of 0.72±0.02, demonstrating the suitability of this assay for HTS.
Introduction
Primary hyperoxaluria (PH) is a rare autosomal recessive hereditary disorder responsible for an excessive production of oxalate that progressively accumulates as calcium salts and forms renal and/or bladder stones. 1 As it progresses, the disease can lead to chronic kidney failure and systemic oxalosis (deposition of calcium oxalate throughout the body). 2,3 Deficiencies in the liver-specific peroxisomal enzyme, alanine:glyoxylate aminotransferase (AGT, EC 2.6.1.44), have been linked to Type 1 PH. 4 Under normal circumstances, AGT catalyzes the transamination of glyoxylate to glycine within the peroxisomes of hepatocytes. In PH1 patients, who represent 80% of the PH population, the AGT deficiency causes the glyoxylate to diffuse from the peroxisomes into the cytosol, where it is oxidized to insoluble calcium oxalate by the lactate dehydrogenase. 1,5 Missense mutations in the AGT gene (AGXT) that lead to the mistargeting of AGT from its normal peroxisomal location to the mitochondria have been identified. 6,7 Although mistargeted, AGT is still catalytically active in the mitochondria, but metabolically ineffective in this location. The best therapies currently rely on dialysis or liver transplantation; both represent inconvenient, risky, and costly procedures. Hence, cellular rescue of AGT misrouting through small-molecule intervention is a promising and potentially safer therapeutic alternative.
Pharmacoperones are cell-permeable small molecules that serve as templates, which correctly fold and route otherwise misfolded/mistrafficked mutant proteins. In particular, G protein-coupled receptors (GPCRs) have been heavily exploited in HTS in terms of pharmacoperone discovery with promising lead scaffolds emerging for the vasopressin 2 receptor (V2R) and the gonadotropin-releasing hormone (GnRH) receptor (GnRHR). The functional rescue of these proteins by pharmacoperones has been reported by our groups and others. 8 –13 The discovery of target-specific pharmacoperones by high-throughput screening (HTS) requires the design of automation-friendly, microtiter plate-compatible assays. 14 Previously reported assays measuring AGT activity are not fully compatible with HTS requirements or are effectively too far removed from the phenotypic and physiological relevance of this target. 15 –17 In this study, we describe a phenotypic microscopy-based assay that detects the colocalization of AGT with the peroxisomes in a mammalian cell-based system expressing a pathophysiologically relevant mislocated mutant form of AGT, AGT-170. 16,18 In the present work, we have successfully miniaturized the assay to a 384-well plate format and demonstrate its HTS readiness through a small-scale pilot screen that paves the way for interrogating larger small-molecule libraries in a fully automated manner.
Materials and Methods
Cell Culture
The CHO-GO (glycolate oxidase) cell lines expressing the AGT-mi and -170 variants were described elsewhere. 6 Cells were routinely cultured in Ham's F12 medium supplemented with 10% fetal bovine serum (Hyclone), 100 units/mL penicillin, 100 μg/mL streptomycin, and 0.25 μg/mL amphotericin B (antibiotic-antimycotic mix; Gibco), as well as 400 μg/mL zeocin and 800 μg/mL G418 (Life Technologies) for selection.
Compound Library
A small set of 1,280 compounds encompassing 4 compound plates from our library was used for the pilot screen. Details regarding Scripps' Drug Discovery collection can be found at
384-Well Plate Assay Protocol
A detailed stepwise protocol is presented in Table 1. Briefly, AGT-170 cells (and AGTmi cells that serve as a positive control) were seeded in black, square IQ™ 100 μ 384-well plates (Aurora; Brooks) at 500 cells/wells using a Flying Reagent Dispenser (FRD; Beckman Coulter). After an overnight incubation at 37°C, 95% relative humidity (RH), and 5% CO2, cells were treated with 100 nL test compounds or DMSO for the positive and negative wells with a pintool transfer unit (GNF). Plates were then placed in an incubator for 3 days at 37°C, 95% RH, and 5% CO2, immunostained, and acquired on a high-content imager as described below.
384-Well Plate Protocol for the AGT High-Content Assay
1. Uses Flying Reagent Dispenser (FRD) technology.
2. Uses PinTool compound transfer technology.
5. Uses FRD technology.
7. Plates were washed using Matrical's Squirt.
8. Uses FRD technology.
10. Plates were washed using Matrical's Squirt.
11. Uses FRD technology.
13. Plates were washed using Matrical's Squirt.
14. Uses FRD technology.
16. Plates were washed using Matrical's Squirt.
17. Uses FRD technology.
19. Acquisition on Cellomics CellInsight.
AGT, alanine:glyoxylate aminotransferase; D-PBS, Dulbecco's Phosphate-Buffered Saline.
Immunostaining
Following treatment and incubation as described above, cells were fixed by the addition of 2× fixation solution (8% paraformaldehyde) using an FRD. Plates were then washed thrice with phosphate-buffered saline (PBS) on the Squirt microplate washer using a 12° nozzle tilt and 15 PSI pressure according to the user's manual for washing normal binding cells in a 384-well plate format (Brook). Cells were then permeabilized by dispensing a solution of 0.1% Triton X-100 in PBS (PBS-TX) and incubated for 10–30 min. Next, cells were stained with a guinea pig anti-AGT A1 antibody (1:10,000; Danpure's Laboratory) and a rabbit anti-PMP70 antibody (1:500; Abcam) in PBS-TX containing 0.5% goat serum. After three PBS washes, anti-rabbit AF488 and anti-guinea pig AF546 secondary antibodies (both at 1:10,000; Molecular Probes) as well as Hoechst at 10 μg/mL were added to each well. Thirty minutes later, plates were washed thrice, filled with 50 μL/well of PBS, and sealed with black tape.
Image Acquisition and Analysis
Plates were read and images acquired with the CellInsight high-content reader (Thermo Fisher Scientific, Pittsburgh, PA) using a 20× objective. A detailed and illustrated description of the image acquisition and analysis is presented in Figure 1. A maximum of nine fields of view per well were acquired, yielding an average of ≈700 cells that were detected. The nuclear stain channel (Hoechst, 386 nm, 9.8 ms exposure time) was used to focus on the cell layer. Puncta labeled with the anti-PMP70 antibody (labeling peroxisomes) and the anti-AGT antibody were acquired in the green (485 nm, 40 ms exposure) and red (549 nm, 15 ms exposure) channels, respectively. Nuclei were detected using the fixed threshold method with an intensity threshold of 300. Next, a cytoplasmic region of interest was created around each nucleus by creating a ring with a distance of one pixel from the nucleus and a width of eight pixels. Peroxisomes and AGT proteins were identified in the cytoplasmic region of interest using a fixed intensity threshold of 200 and 141, respectively. Colocalization of the two populations of puncta was determined using the Cellomics' Colocalization BioApplication; the output feature providing the Pearson's correlation coefficient between the two targets is called ROI_A(B)_Correlation Coef. This feature, which shows values ranging from −1 to 1, describes the degree of overlap of the two targets with each other independent of the intensity differences of the two targets, and is calculated using the following equation:
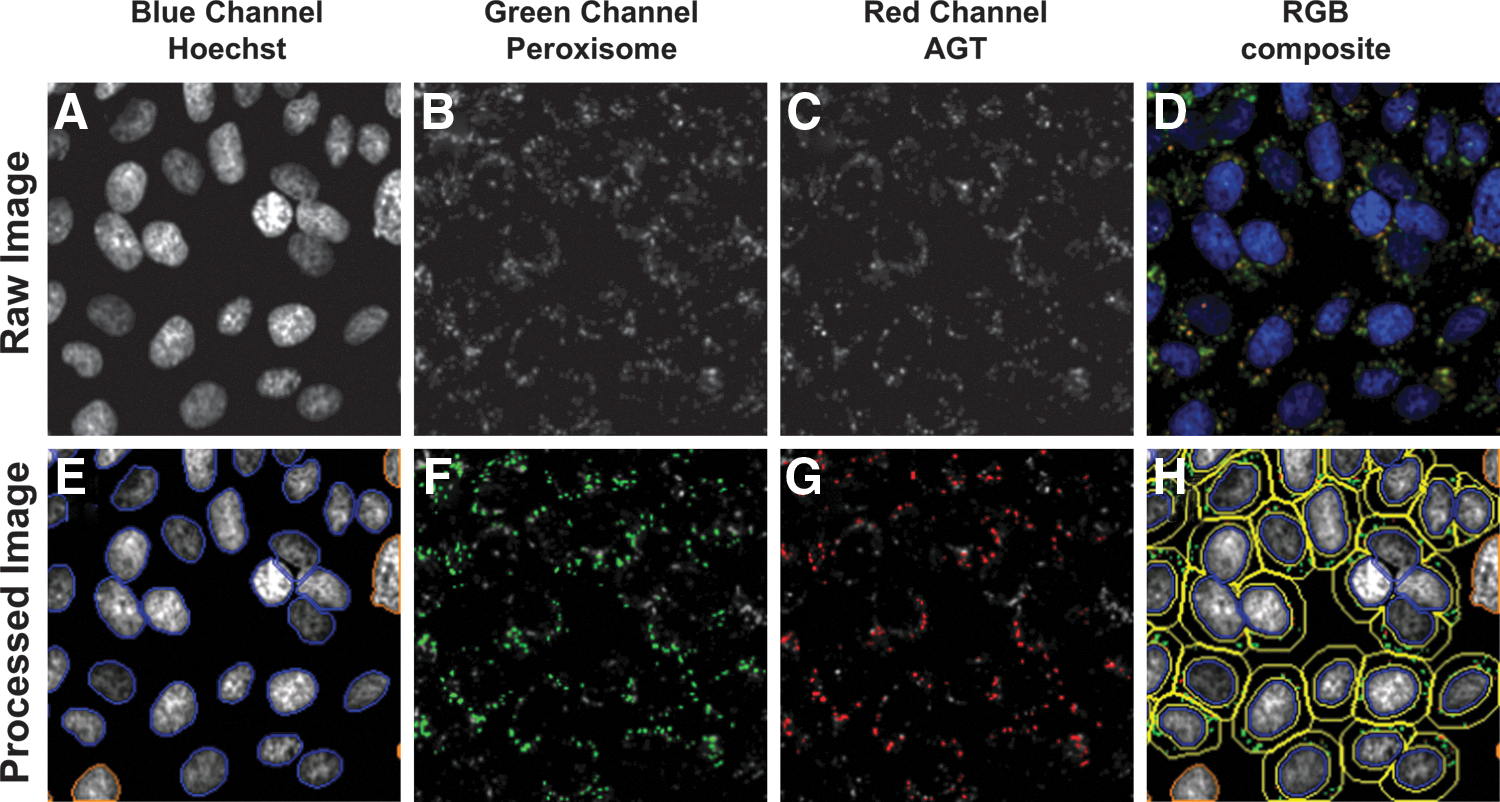
Automated image acquisition and analysis to evaluate the colocalization of AGT with the peroxisomes.
where rp
is the Pearson's correlation coefficient, x and y are pixel intensities of each pixel detected in the AGT and peroxisome channels, respectively, and
Images and data were automatically spooled to the STORE database (Thermo/Cellomics) hosted on Scripps's HTS server.
Data Management and HTS Data Analysis
The relevant well feature generated by the detection algorithm, named the MEAN_ROI_A_Correlation Coef, was exported from the STORE database as a tabulated file using a custom Simple Object Access protocol (SOAP) web service utilizing Thermo's high-content screening connect API. Reader files were then uploaded into Scripps' Drug Discovery database (Symyx Assay Explorer). Plate Z-factor, Z′-factor, and sample-to-background ratio (S/B) were automatically calculated as previously described.
19,20
Assay results were normalized on a per plate basis using the following equation:
where High Control represents the well containing AGTmi cells treated with DMSO (n=32) and Low Control represents the well containing AGT-170 cells also treated with DMSO (n=16).
Results
Development of an HTS-Compatible Microscopy-Based Assay to Monitor AGT-Peroxisome Colocalization
Previously, we described the use of stably transformed CHO cells to study the subcellular distribution of normal and various PH1 mutant AGT constructs by immunofluorescence microscopy. 6 Among these mutants, the AGT-170 variant presented the most severe peroxisome-to-mitochondrion mistargeting phenotype. Accordingly, we incorporated a cell line expressing this specific mutation to design a high-content high-throughput assay to identify potential pharmacoperones able to rescue AGT mistargeting by rerouting it to the peroxisomes. To be compliant with HTS requirements, the original assay had to be simplified, adapted to microtiter plates, and automated.
We first investigated our ability to detect and measure AGT/peroxisome colocalization with our HTS-compatible, wide-field high-content imager by labeling CHO cells expressing either the AGT-170 PH1 mutant or the AGT minor allele for nuclei, peroxisomes, and AGT in a 96-well plate format. Acquisition was made using a 20× objective. A dedicated detection algorithm was defined that automatically calculates and reports the Pearson's colocalization coefficient between the AGT- and peroxisome-labeled areas for each well (Fig. 1). Figure 2 shows images from a representative well of each of the two cell lines as acquired with the high-content reader. Upon automated analysis, the Pearson's colocalization coefficient was determined at 0.85±0.08 for AGTmi cells and 0.56±0.15 for AGT-170 cells, values that are in accordance with those reported in our previous work using a confocal laser-scanning fluorescence microscope, a 60×objective, cells fixed on glass coverslips, and the ImageJ JACop plugin. 6 This indicated that a lower magnification wide-field instrument geared toward higher throughput was also able to properly measure the biology of interest.
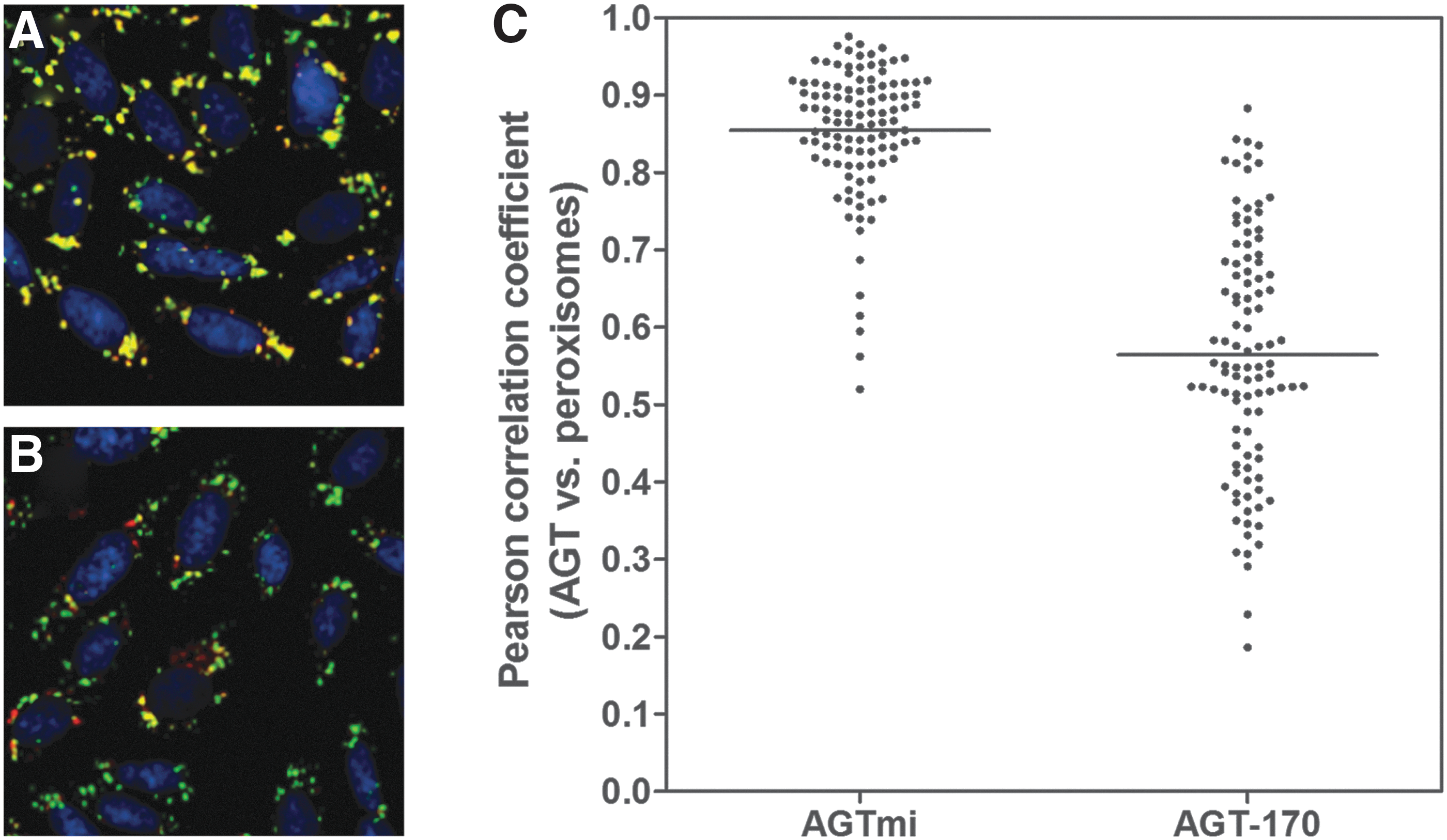
Differential localization of the AGTmi and AGT-170 proteins with regard to the peroxisomes. RGB composite images of CHO-K1 cells stably expressing the AGTmi
To assess our ability to induce and detect an increase in the Pearson's colocalization coefficient, we treated both AGTmi and AGT-170 cells with glycerol, a viscous polyol compound known to enhance the stability of proteins in solution. 21 This cosolvent shifts the native protein to more compact states and inhibits protein–protein aggregation while proteins are being refolded during biological synthesis. Accordingly, treatment with increasing concentrations of glycerol resulted in a marked increase of Pearson's colocalization coefficients for the AGT-170 cells changing from 0.41±0.01 to 0.80±0.04 (n=4), that is, a 1.91-fold increase as shown in Figure 3A. Notably, AGTmi cells were also showing an increase in Pearson's values upon glycerol treatment, although this increase was marginal (∼1.10-fold increase). Concentrations of glycerol higher than 5% affected cell viability prevented accurate determination of the Pearson's colocalization coefficient; for these reasons, they were avoided. To probe the robustness of the assay, we repeated the aforementioned glycerol titration assay on two separate plates. The plot presented in Figure 3B shows the Pearson's coefficient determined from multiple wells treated with varying glycerol concentrations ranging from ≈0.4 to ≈0.9 and indicates that data generated from the two separate plates were virtually identical, yielding an R 2 greater than 0.99. The Z′-factor, a statistical measurement indicative of HTS readiness, 20 was calculated between the AGT-170 cells in the absence and presence of 5% glycerol and found to be 0.58 and at 0.81 between untreated AGT-170 and AGTmi cells.

Effect of glycerol on the colocalization of AGT with the peroxisomes. CHO-K1 cells expressing either AGTmi (gray bars) or AGT-170 (white bars) were cultivated for 4 days in 96-well plates in the presence of different concentrations of glycerol ranging from 0 to 5%. Images were then acquired and the Pearson's correlation coefficient calculated
Miniaturizing and Automating the AGT-170 Cell-Based Assay to the 384-Well Plate Format
With Z′-factor values greater than 0.5, the assay warranted further miniaturization and automation to the 384-well plate format. Whereas the majority of HTS assays that are homogeneous in format typically rely on three dispense steps (cell addition, compound delivery, detection reagents dispense), and immunostaining-based assay protocols usually require a minimum of eight dispenses, including aspiration and/or wash steps. A primary concern was that automated liquid handling devices will disrupt the cell layer during this procedure. To prevent this from occurring, we chose to fix the cells as early as possible in the protocol by replacing the initial media aspiration step with a dispense of a 2× concentrated fixation solution. In addition, we implemented a noncontact plate washer that does not rely on aspiration to empty the wells, but instead utilizes an air blade that ejects liquids from the well with little to no residual volume (Squirt; Brooks). To verify that cells remained in the wells and were not being dislodged during the plate preparation process, Hoechst staining was added during the fixation step of a mock immunostaining protocol and nuclei were counted at each step. The results presented in Figure 4 indicate that the optimized dispensing and washing conditions assured little to no cell detachment over the course of the plate preparation process. These results also indicated that cells seeded at densities higher than 600 cells per well reached total confluence; a cell seeding density of 500 cells per well was considered optimal and used for the rest of this study. Volumes, concentrations, and incubation times were optimized to offer the best balance between cost per well, time, and assay performance. The resulting miniaturized assay protocol is presented in Table 1.
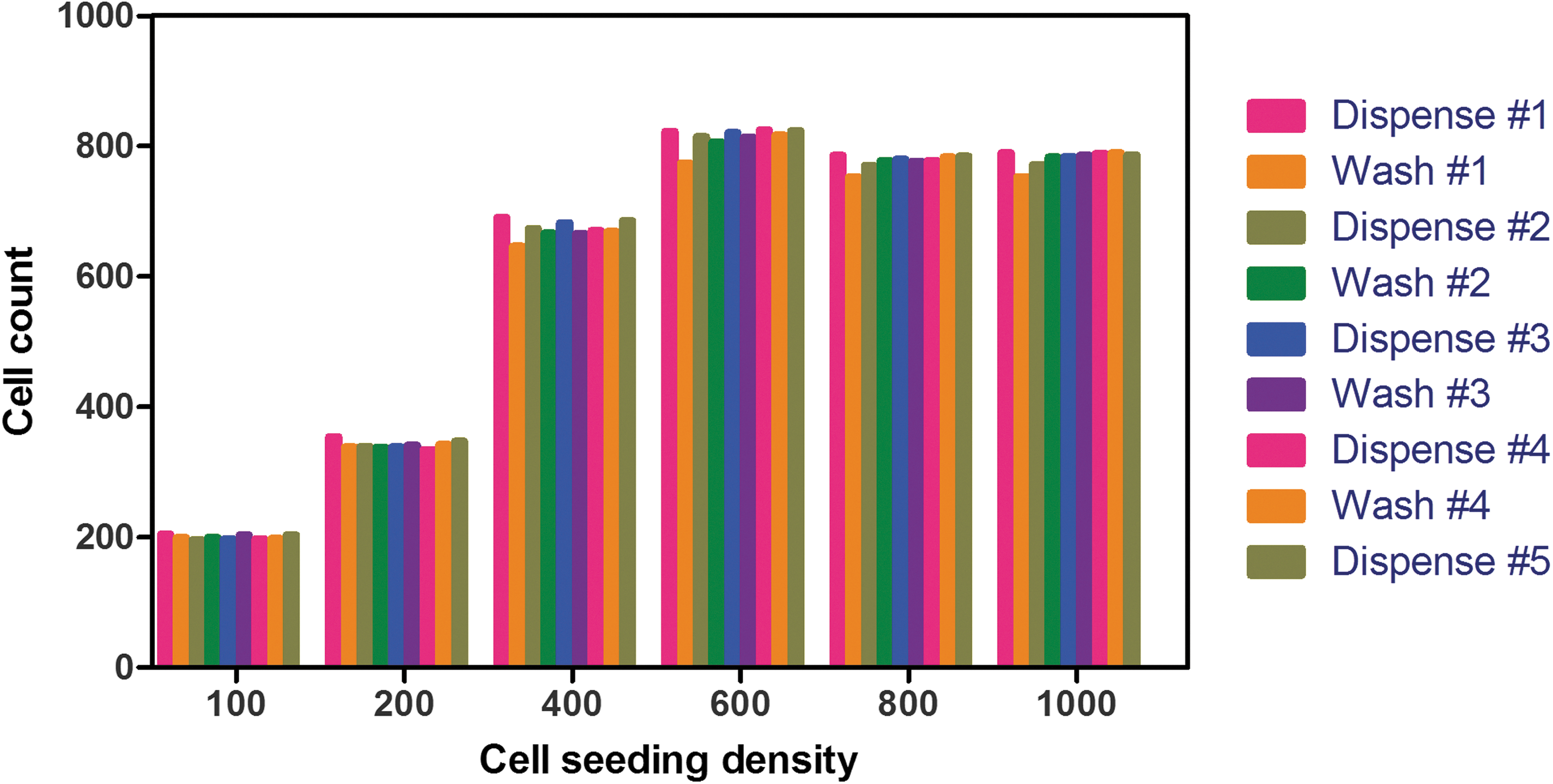
The effect of multiple dispense/wash steps using noncontact, automated instruments on the cell layer integrity of CHO-K1 cells cultivated in a 384-well plate. CHO-K1 cells were plated in 384-well plates at different cell densities ranging from 100 to 1,000 cells per well (64 wells per condition) and incubated for 4 days. In this case, cells were immediately fixed and their nuclei labeled by the addition of 2× fixation solution (8% paraformaldehyde) containing 20 μg/mL Hoechst using an FRD. After each step, as indicated in the stepwise protocol in Table 1, the cell number was determined using a CellInsight high-content reader after imaging a single field of view with a 5× objective. FRD, Flying Reagent Dispenser.
Validation for HTS Purposes
To verify that the assay was compatible with HTS requirements, we first investigated the potential for position effects by running plates treated with DMSO only. A heat map of a representative plate treated with DMSO is shown in Figure 5; this plate did not show any edge effect or position effect within the sample field, indicating that despite an extended incubation time (4 days), all variables that can lead to well-to-well variability (such as temperature gradient, evaporation, and contamination) were tightly controlled. The coefficients of variation of 6.12%, 1.17%, and 7.47% for the sample field, high control, and low control, respectively, were well under an empirically accepted maximum of 10%. The Z′-factor was 0.62, demonstrating the miniaturized AGT assay's robustness and readiness for drug screening purposes. In addition, the lack of observable outliers within the sample field in general indicates this assay should be fairly devoid of false positives or negatives. The assay does have at least some indication of DMSO sensitivity as observed in column 24 of Figure 5 with the normalized data displaying a negative % rescue for nontreated wells.

Example of raw and normalized data from a representative DMSO plate. Five hundred CHO-K1 cells expressing either AGTmi or AGT-170 were plated in a 384-well plate and incubated overnight. The following day, all wells, but the last column, were treated with DMSO (0.9% final concentration) using a pintool transfer unit. After 3 days, plates were prepared for automated high-content imaging.
Pilot Screen
To assess the performance of the AGT assay under HTS conditions, a pilot screen against a set of 1,280 diversified molecules was conducted. The final concentration at which the compounds were tested was 4 μM, which corresponds to a final DMSO concentration of 0.4%. A total of six plates were used for this assay, comprising four separate compounds plates and two DMSO plates, one at the beginning and at the end of the run. The S/B, Z-, and Z′-factors of each plate are reported in Figure 6A. With an average Z′-factor of 0.72±0.02 and a steady S/B at 1.81±0.09, the pilot screen exhibited satisfactory assay statistics and the control wells excellent plate-to-plate reproducibility, which warranted further analysis. To identify compounds, which caused a significant relocation of AGT-170 to the peroxisomes (i.e., significant percent rescue), two values were calculated: (1) the average percent rescue of all test compounds and (2) three times the standard deviation of the percent rescue of all test compounds. The sum of these two values, (1) and (2), was used as a cutoff parameter and calculated at 29.43% rescue [with (1)=3.75% and (2)=3×8.59%=25.68%]. Compounds showing a percent rescue that was higher than the hit cutoff were considered as hits (4 compounds were identified out of 1,280, that is, a 0.31% hit rate).
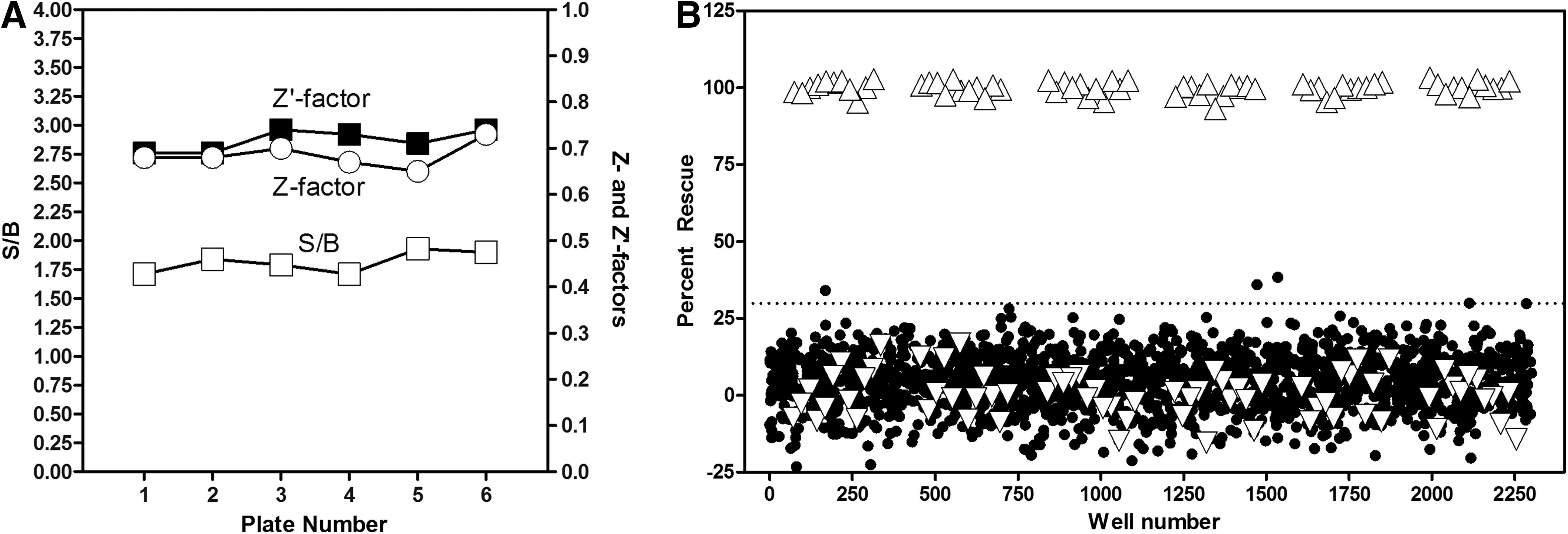
AGT assay pilot screen results.
Discussion
The PH1-triggering AGT-170 mutation has been shown to be responsible for the misrouting of the enzyme from the peroxisomes to the mitochondria. Although functional in the mitochondria, the enzyme must reside in the peroxisomes to properly convert toxic glyoxylate into glycine. Failure to do so results in the accumulation of glyoxylate in the peroxisomes, which is then transported to the cytoplasm where it is converted to oxalate by the lactate dehydrogenase, potentially leading to kidney failure. Pharmacoperones are small molecules that serve as molecular templates to help mutant misfolded proteins to properly fold and route correctly within the cell. Since the enzymatic activity of AGT-170 is maintained despite the mutation, its correct rerouting to the peroxisome is considered a promising therapeutic option. The use of small-molecule pharmacoperones would be preferred to more controversial strategies such as gene therapies aiming at providing a nonmutated version of AGXT 22 or those aiming at pharmacologically modulating the activity of protein chaperones such as the HSPs 23 that are involved in the proper folding of hundreds of other proteins.
In this report, we describe the design, miniaturization, and validation of a cell-based HTS assay able to monitor the rerouting of the AGT-170 mutant protein to its correct location, the peroxisomes. Automated high-content microscopy was considered the technology of choice to accurately measure AGT rerouting; it was indeed favored over other technologies relying on reporter systems that usually require creating fusion or tagged proteins and can potentially lead to assay artifacts. 24 In contrast, using immunodetection allowed us to use AGT proteins that are devoid of any modifications and hence represent exact carbon copies of those found in patients.
Surprisingly, it appears that the use of a wide-field microscope yields enough spatial resolution to be able to clearly map the AGT location with regard to the peroxisomes. Even though our previous work relied on four acquisition channels (nuclear stain, anti-AGT and peroxisome immunostaining, and MitoTracker Red), we reasoned that for HTS purposes, the MitoTracker channel could be removed since it is now well documented that the AGT-170 mutant protein is predominantly located in the mitochondria. The goal is indeed to confirm its rerouting to the peroxisome upon treatment with a hit compound rather than its departure from the mitochondria.
In addition to the biology of interest and without any additional plate preparation, high-content imaging offers the opportunity to capture additional cell features, such as cell count, nuclear intensity, morphology, and condensation, which can help identify undesirable cytotoxic compounds early during the screening process.
As noted, DMSO itself was shown to have an effect on the % rescue in our assay (Fig. 5A). The fact that all test compounds were prepared in DMSO and that the low control 0% rescue wells were also receiving an equivalent DMSO concentration demonstrates that DMSO should not interfere with our ability to detect compounds able to reroute AGT to the peroxisomes.
An explanation for the low hit rate as well as the relatively low % rescue of the four hits identified is that the exposure time to compounds was potentially too long. We may reasonably assume that potentially active compounds will degrade over such an extended period of time, masking their effect. Another important consideration is that AGT already synthesized and mistargeted to mitochondria may not be affected by pharmacoperones; the latter can indeed only affect newly synthesized AGT before it has been imported. Therefore % rescue values are most likely underestimated, and the effect of potential pharmacoperones might be more effective in a real-life situation than they first appear. Regardless, time-course experiments with DECA, a mitochondria translocation blocker we recently identified during a chemical genetic screen on yeast, 17 should help us determine in an unbiased way the optimal incubation time that is relevant to the biology we are interrogating. In addition, we plan on further miniaturizing the assay to the 1,536-well plate format, as routinely performed in our laboratory and by others, 25 to reduce reagent and consumable costs by roughly four-fold.
In conclusion, we demonstrated the implementation of a novel, cost-effective, and robust, miniaturized high-content assay for the discovery of pharmacoperones that can rescue an enzyme-trafficking defect involved in
Footnotes
Acknowledgment
This work was supported by the Oxalosis and Hyperoxaluria Foundation (OHF) grant, Development of HTS Assays for Drugs for Hyperoxaluria, to P.M.C.
Disclosure Statement
No competing financial interests exist.