
Abstract
The aim of this study was to evaluate the ability of the well-known radical scavenging compound edaravone (EDA) to entrap and detoxify reactive carbonyl species (RCS) derived from lipid peroxidation [4-hydroxy-trans-2-nonenal (HNE), acrolein and glyoxal], as well as its ability to prevent RCS-induced protein carbonylation, by using hemoglobin (Hb) modified by HNE as an in vitro model. Through a combined HPLC and high-resolution mass spectrometric approach, we confirmed the ability of EDA to scavenge precursors for either advanced glycation or lipoxidation end products (EAGLEs), such as glyoxal, and demonstrated for the first time that EDA is also a potent quencher of α,β-unsaturated aldehydes (providing mass spectral characterization of the adducts), being significantly more active than a series of well-known RCS sequestering agents. Direct infusion analysis of the intact protein and nano LC-ESI-MS/MS analysis of the tryptic digest, carried out on an LTQ-Orbitrap hybrid mass spectrometer, were used to study the modifications occurring on Hb after exposure to increasing HNE concentrations, providing evidence for Cys93 (Hb β-chain) involvement in covalent attachment, and to demonstrate the ability of EDA dose-dependently to inhibit Hb carbonylation. Computational studies allowed us to elucidate the mechanism of EDA-RCS interaction and to explain the preferential site of HNE adduction to Hb. The same combined approach indicated that EDA is not a selective RCS scavenger, being able to react also with nontoxic, physiologically relevant aldehydes, such as pyridoxal. Antioxid. Redox Signal. 12, 381–392.
Introduction
Taking RCS and carbonylation damage as a drug target, the first molecular strategy considered to neutralize/reduce these pathogenetic factors was based on free radical scavengers that prevent lipid peroxidation by capturing highly reactive oxygen species (ROS). As an alternative strategy to the antioxidant approach, several efforts have been made in the last few years to discover efficient RCS scavengers, nucleophilic compounds able to entrap highly electrophilic, highly cytotoxic lipid-derived RCS (identified as the chemical intermediates between hyperglycemia, hyperlipidemia, and their complications), and to detoxify them by forming covalent and unreactive adducts (RCS-sequestering agents) (1, 2, 17).
In other words, the first compounds might prevent the RCS formation, whereas the last ones might neutralize the RCS effects, avoiding their accumulation. The nonenzymatic antioxidants normally include reducing compounds (ascorbic acid, α-tocopherol, carotenoids, and flavonoids) which are able to capture one electron, whereas reactive carbonyl scavengers typically involve nucleophilic compounds (histidine-containing dipeptides, aminoguanidine, and hydralazine), able to form covalent products with RCS mainly through Michael-type addition. Usually, the investigated molecules do not combine both effects, even if a very important exception is represented by endogenous glutathione (GSH) whose sulfhydryl group can act as a nucleophile or reducing agent or both, being oxidized into the corresponding disulfide homodimer (GSSG). Another promising molecule that may combine the two mentioned mechanisms is edaravone (MCI-186, 3-methyl-1-phenyl-2-pyrazolin-5-one), whose activity is truly original, being based on the acidity of the methylene group in α to a carbonyl function.
Edaravone (EDA) is a well-known free radical scavenger that has been shown to have protective effects against cerebral ischemia/reperfusion injuries in a variety of experimental animal models. The clinical efficacy of edaravone against ischemic brain attack has been demonstrated by the presence of significant improvements in functional outcome in a randomized, placebo-controlled, double-blind study, and indeed, it has been approved by the Japanese health authorities as a neuroprotective agent for the treatment of acute cerebral infarction since 2001.
In addition to these antistroke effects, edaravone has been shown to prevent oxidative damage to various extracerebral organs (29) and to inhibit accumulations of HNE and 8-hydroxy-deoxyguanosine, which are oxidative byproducts, as well as neuronal cell death after transient focal ischemia in the murine brain (32). Systemic administration of edaravone attenuates the increase of malondialdehyde levels, the reduction of superoxide dismutase (SOD) activity, and suppresses the retinal dysfunctions after retinal ischemia/reperfusion in rats, as well as oxidative stress-induced endothelial damage and early atherosclerosis in ApoE-KO mice (30). More recently, the screening of a large chemical library disclosed that edaravone inhibits in vitro the advanced glycation end product (AGE) formation efficiently (14), by trapping both reactive carbonyl compounds (glyoxal and methylglyoxal) precursors for EAGLEs and AGEs, respectively.
Although the mechanisms for radical scavenging of EDA were deeply explored with several approaches, little or nothing has been done to investigate the precise mechanisms by which the compound can react with toxic and nontoxic physiologically relevant carbonyl compounds. Because generation of drugs sharing both antioxidant and carbonyl scavenger properties represents a new therapeutic challenge in the treatment of carbonyl stress-associated diseases, this study was undertaken to gain a deeper insight into the RCS trapping ability of EDA, and in particular, (a) experimentally to confirm, by an unequivocal mass-spectrometric approach, the reactivity toward carbonyl compound precursors of EAGLEs; (b) to evaluate its potency as a scavenger of highly reactive and cytotoxic lipid-derived α,β-unsaturated aldehydes (HNE and acrolein), precursor of ALEs (advanced lipoxidation end products), never considered; (c) to unveil mechanisms, stoichiometry, and reaction products of EDA with both toxic and endogenous (pyridoxal) aldehydes; and (d) to demonstrate the ability of the compound to prevent/inhibit protein carbonylation, by using RCS-modified hemoglobin as an in vitro model.
Materials and Methods
Chemicals and reagents
Human hemoglobin (Hb), hydralazine (HY), pyridoxamine (PYR), pyridoxal (PYAL), acrolein (ACR), and glyoxal (GO) were purchased from Sigma-Aldrich (Milan Italy). Because Hb was predominantly methemoglobin, before use, it was dissolved in 2 mM Tris–HCl (pH 7.5), reduced by 1,4-dithioerythritol (DTE, 0.5 mM), and then dialyzed overnight with Milli-Q water. Edaravone (3-methyl-1-phenyl-2-pyrazolin- 5-one; EDA) was from Tocris Bioscience (Bristol, U.K.). 4-Hydroxy-non-2-enal diethylacetal (HNE-DEA) was synthesized according to the literature (23). 4-Hydroxy-trans-2-nonenal (HNE) was prepared from HNE-DEA with 1 mM HCl hydrolysis (1 h at room temperature) and quantified with UV spectroscopy (λ max, 224 nm; ɛ, 13,750 l mol/cm). Carnosine (β-alanyl-
Protein carbonylation: MS studies
Sample preparation
HNE-induced carbonylation of Hb was studied by incubating 20 μM Hb solution with HNE at different molar ratios (Hb/HNE, 1:1; 1:5; 1:10) in 1 mM phosphate buffer (pH 7.4) for 120 or 240 min at 37°C. The dose-dependent effect of EDA on Hb carbonylation was studied by co-incubating for 120 min 20 μM Hb in the presence of 200 μM HNE at different EDA doses (40, 200, and 400 μM).
Direct infusion mass spectral analysis
To detect changes in the protein mass of the α- and β-globin subunits, samples prepared as described earlier were analyzed with direct infusion into a LTQ-Orbitrap Hybrid Mass Spectrometer (Thermo Scientific, Milan, Italy) equipped with an Electrospray Finnigan Ion Max source. Aliquots (50 μl) of each sample were mixed with 150 μl of H2O:CH3CN:HCOOH (50:50:0.1, vol/vol/vol), infused into the mass spectrometer at a flow rate of 3 μl/min and analyzed in full-scan mode, under the following instrumental conditions: AGC target, 5 × 104; 500-ms maximum inject time; one microscan, scan time, 1.9 s; resolving power, 100,000 positive ion mode; capillary temperature, 270°C, spray voltage applied to the needle, 3.5 kV; capillary voltage, 37 V; nebulizer gas (nitrogen) flow rate set at 5 (a.u.); acquisition time, 1 min. Direct infusion ESI-MS spectra were deconvoluted by using Xtract for Qual Browser v. 2.0.7 (ThermoQuest).
Nano LC-ESI-MS/MS analysis
The HNE-induced structure modifications of Hb were characterized with nano LC-ESI-MS/MS analysis of incubated samples after reduction with NaBH4, an established procedure for adduct stabilization (7, 12), followed by enzymatic digestion, by using trypsin as proteolytic enzyme and iodoacetamide as an alkylating agent. Peptide mass mapping provided identification of the peptides, accounting for ∼96% of the protein sequence. All digested peptide mixtures were separated with online reversed-phase (RP) nanoscale capillary liquid chromatography (nanoLC) and analyzed with electrospray tandem mass spectrometry (ESI-MS/MS). Chromatography was performed by using a Surveyor LC system (ThermoFinnigan Italia, Milan, Italy) on a 180-μm × 10-cm column packed with a 5-μm, Biobasic-18 stationary phase (Thermo, Superchrom, Milan, Italy). The pump flow rate was split 1:75 for a column flow rate of 1 μl/min. The column effluent was directly electrosprayed by using the silica emitter source without further splitting. Solvents A (water containing 0.1% HCOOH) and B (methanol containing 0.1% HCOOH) were used as a mobile phase. The separation of tryptic peptides was achieved by gradient elution from 0 to 100% B over a 65-min period. Before the next analysis, both the precolumn and the column were first washed with 100% solvent B for 10 min and then equilibrated with 100% solvent A for 20 min. For the identification of peptides, an LTQ XL-Orbitrap mass spectrometer was used (Thermo Scientific, Milan, Italy) and the electrospray interface (dynamic nanospray probe; Thermo Scientific) was set as follows: spray voltage, 1.6 kV; capillary temperature, 220°C; capillary voltage, 30 V; tube lens offset, 120 V, and no sheath or auxiliary gas flow. The analyses were performed in positive polarity enabling the data-dependent scan mode. The MS spectra were acquired in profile mode by Orbitrap in the following conditions: scan range, 250-2,000 m/z in full-scan mode with full-scan injection waveforms enabled; AGC target setting, 5 × 105; maximum inject time, 500 ms; scan time, 1 s (60,000 resolving power at 400 m/z, FWHM). A list of 14 protonated phthalates and siloxanes including dibutylphthalate (plasticizer, m/z 279.159086); bis(2-ethylhexyl)phthalate (m/z 391.284286) and dodecamethylcyclohexasiloxane [(Si(CH3)2O)6 + H]+; m/z 445.120025] were used for real-time internal mass calibration (19). Tandem mass spectra were recorded by the linear ion trap in centroid mode for the three most intense ions (isolation width, 2 m/z; normalized collision energy, 35 CID arbitrary units; minimum signal threshold, 5 × 104), and dynamic exclusion was enabled (repeat count, three; repeat duration, 10 s; exclusion-list size, 25; exclusion duration, 120 s; relative exclusion mass width, 5 ppm). Charge-state screening and monoisotopic precursor selection was enabled, singly and unassigned charged ions were rejected.
HNE adduct identification
The acquired MS/MS spectra were searched by using the Bioworks software (rev. 3.3.1 sp1; Thermo Scientific) and by using a database containing only the protein of interest. The following settings were used for the search engine: mass type, monoisotopic precursor and fragments; enzyme, trypsin (KR); missed cleavage sites, 5; peptide tolerance, 5 ppm; fragmentations tolerance, 0.5 AMU; modifications, 5 PTMs per peptide; the variable modifications considered were as follows: HNE reduced Michael adduct to His, Lys, and Cys (+158.13067 Da as monoisotopic mass); reduced HNE Schiff base with the ɛ amino group of Lys (+140.12011 Da as monoisotopic mass); and fixed modification: carbamidomethylation on Cys. Results were filtered by setting the peptide probability to 0.005.
Reactivity and selectivity
HPLC studies
The reactivity of EDA and of the reference compounds toward different RCS (scavenging activity) was evaluated by measuring their consumption (50 μM in 10 m
Selectivity was determined in a similar manner after 180-min incubation, by using PYAL as a target physiologically relevant aldehyde. PYAL consumption was determined with HPLC by using a fluorimetric detector, as previously reported (16). The results are reported as percentage of the free aldehyde remaining with respect to a blank, incubated in the absence of the tested compound.
Direct-infusion MS experiments
Structure characterization of the adducts was determined with MS studies (direct infusion) on a triple-quadrupole (TQ) mass spectrometer (Finningan TSQ Quantum Ultra, ThermoQuest) equipped with an Electrospray Ion Max source at a flow rate of 10 μl/min. Samples were prepared by incubating for 180 min 0.5 mM EDA in 1 mM phosphate buffer (pH 7.4) with an equimolar concentration of the target aldehyde (HNE, ACR, and PYAL in 1 mM phosphate buffer, pH 7.4). An aliquot of the mixture was then diluted 1:4 (vol/vol) with H2O:CH3CN (70:30; vol/vol) and analyzed with direct-infusion MS. Mass spectrometric analyses were performed in positive ion mode. ESI interface parameters were set as follows: middle position; capillary temperature, 270 °C; spray voltage, 4.0 kV. Nitrogen was used as the nebulizing gas at the following pressure: sheath gas, 30 psi; and auxiliary gas, 5 a.u. For each compound tested, the most suitable instrumental conditions were searched for by using the Quantum Tune Master software (Thermo Scientific, Milan, Italy). Elemental composition of each reaction product was determined on the basis of exact mass measurements (resolution, 100,000) obtained with direct infusion on an LTQ XL-Orbitrap hybrid mass spectrometer (Thermo Scientific) equipped with an Electrospray Finnigan Ion Max source by using a list of 14 protonated phthlates and siloxanes, including dibutylphthlate (plasticizer, m/z 279.159086); bis(2-ethylhexyl)phthalate (m/z 391.284286) and dodecamethylcyclohexasiloxane [(Si(CH3)2O)6 + H]+; m/z 445.120025] for real-time internal mass calibration (19).
Molecular formulas were generated by the Elemental Composition feature included in Xcalibur Qual Browser 2.0 (Thermo Scientific) by following the rules described by Kind and Fiehn (26). The maximum error of mass measurements was set at 5 ppm; the nitrogen rule and a restriction for carbon atoms in each formula were applied, considering a 5% relative error in the 12C/13C isotope ratio measurements.
Computational details
Docking studies
The different abundance of the HNE-Hb adducts was investigated in silico by docking the HNE molecule into the experimentally resolved structure of hemoglobin (PDB Id: 1 HGC). Specifically, the retrieved protein structure was completed by adding the hydrogen atoms. At physiologic pH, Arg, Lys, Glu, and Asp residues were preserved ionized, whereas His and Cys residues were considered neutral by default. The structure underwent a global minimization with backbone atoms and heme groups fixed to preserve the experimental folding. The obtained hemoglobin structure was then exploited in docking calculations by using the Biodock program (22), which performs a fully blind search, randomly rototranslating the HNE structure over the entire hemoglobin surface. In detail, Biodock produced 1,000,000 possible poses clustered in ∼5,000 nonredundant solutions that were scored according to the CVFF non–bond-interaction energy. The best docking result around Cys β93 was finally minimized, keeping the atoms fixed outside 15 Å around the docked HNE molecule to preserve the hemoglobin structure. All mentioned minimizations were carried out with Namd2.51 by using the conjugate gradient algorithm until RMS = 0.01.
Reactivity analyses
To rationalize the scavenging activity of edaravone toward ACR and HNE, the reactivity of the reported adducts was analyzed by calculating the molecular softness that is a quantum mechanical index of the global reactivity toward nucleophilic reactions. According to Koopmann approximation, molecular softness (S) was computed by HOMO/LUMO energies [i.e., S = 1/(ELUMO – EHOMO)], as derived by semiempiric calculations by using MOPAC2009 (Keywords: “PM6,” “PRECISE,” “GEO-OK,” and “MULLIK”). When more adducts were possible, the potential regioselectivity of monitored additions was investigated by calculating local reactivity descriptors, as expressed by condensed atom Fukui indices (3).
Results
HNE-induced carbonylation of Hb
The first step of this work was to evaluate the ability of EDA to inhibit protein carbonylation, by using Hb as a model protein covalently modified by HNE, the highly reactive α,β-unsaturated aldehyde generated by lipid peroxidation. To do this, high-resolution mass spectrometry was initially used to study the stoichiometry of the Hb-HNE reaction, and to identify the target nucleophilic sites for HNE adduction on the protein, never reported in the literature. Figure 1a shows the ESI-MS spectrum of human Hb recorded at a resolution of 100,000, characterized by several multicharged ions relative to the beta (Hbβ) and alpha (Hbα) chain (see the deconvoluted spectrum in Fig. 1b). When Hb was incubated in the presence of HNE at a molar ratio 1:1 for 120 min at 37°C (Fig. 2a), no detectable adducts of both α and β chains were observed. By increasing the HNE concentration (Hb/HNE molar ratio, 1:5), a very detectable HNE adduct of the β chain was observed at 16,022.41 Da, shifted by + 156 Da with respect to the native form (15,866.26 Da), and attributed to the HNE Michael adduct (Fig. 2b). By further increasing the HNE concentration (1:10 molar ratio; Fig. 2c), the relative abundance of the adduct increased in a dose-dependent manner, to reach a relative abundance greater than 10%. No adducts on the α chain were observed at all the HNE concentrations tested (Fig. 2).
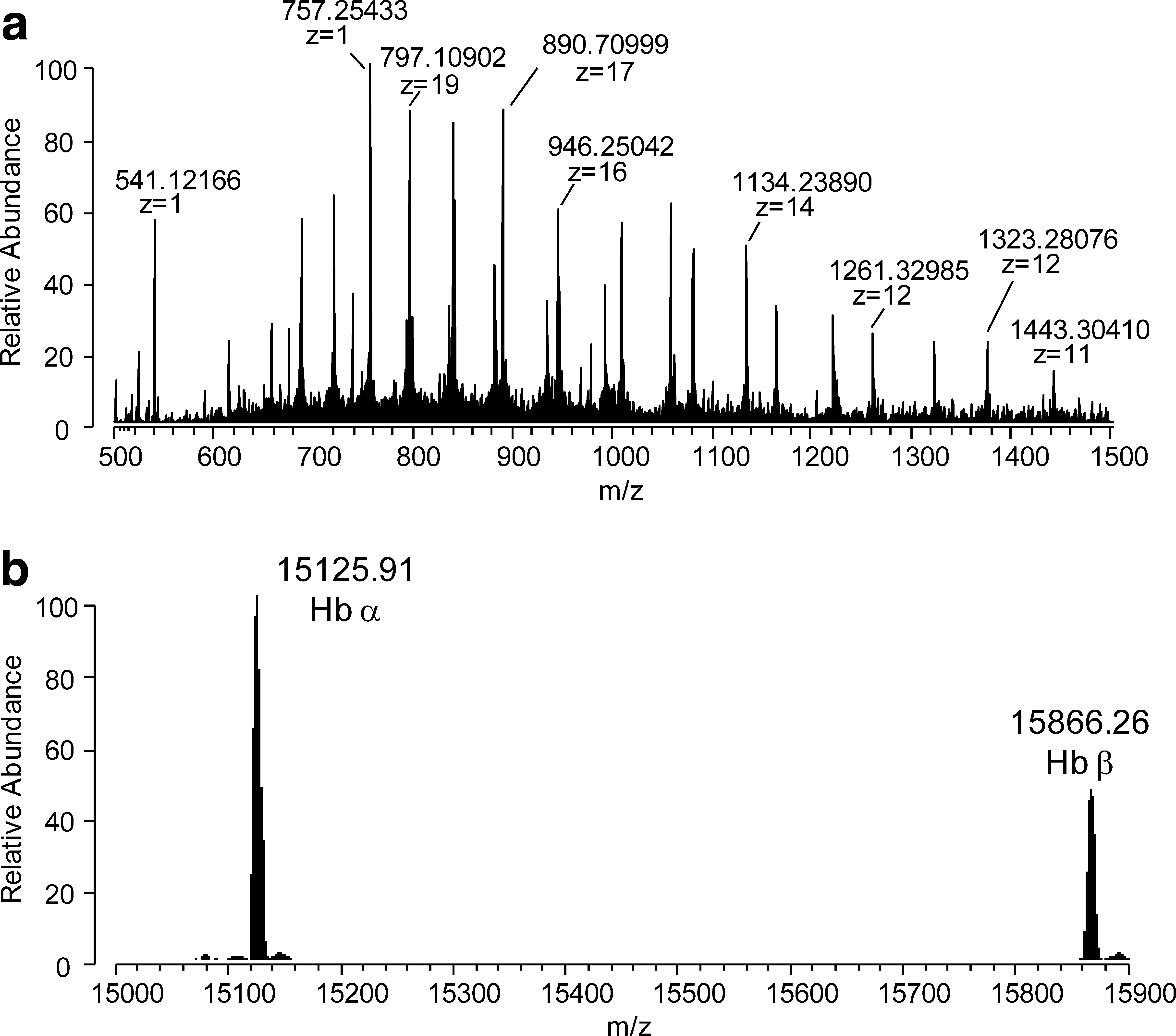
In a second set of experiments, the incubation time was extended to 240 min. In these conditions, HNE (Hb/HNE molar ratio, 1:10) induced a weak, but significant HNE-covalent modification of the α chain, which was detected at 15,282.08, consistent with a shift of 156 Da compared with the native α chain. The abundance of the Michael adduct on the β chain significantly increased with respect to the native chain, and, in addition, a double HNE Michael adduct appeared at 16,177.53 Da (Fig. 3).

The adducted amino acids were then characterized with LC-ESI-MS analysis of trypsin-digested Hb, previously incubated for 240 min with HNE (Hb/HNE, 1:10 molar ratio) and then reduced by NaBH4. HNE is well known to react through a Michael adduction with the nucleophilic sites of His, Lys, and Cys, as well as to form a Schiff base with the ɛ-amino group of lysine (2). Based on these well-known reaction mechanisms, the covalent modifications of the nucleophilic sites (His, Lys, Cys) with HNE were predicted and, for each adduct, the monoisotopic mass shift of the corresponding reduced form was calculated and used as a variable modification in the Sequest search. By using this approach, several HNE Michael adducts were identified on both Hb chains. Table 1 summarizes the covalently modified sites involving the Michael adduct formation on His, Lys, and Cys residues, and the Schiff base on the Lys residue. Besides a qualitative pattern, we also were interested in comparing the chemical reactivity of the target amino acids toward HNE, to identify the most reactive and nucleophilic sites. To do this, the chromatographic peak of each modified peptide was reconstituted by selecting the most intense multicharged ion as filter, and the peak areas were calculated and compared with those of the corresponding unmodified peptides. The results reported in Table 1 indicate Cysβ93 as the most reactive site, because the peptide containing the HNE modification accounts for more than 40% of the Cysβ93-containing peptides. Figure 4 shows the reconstituted LC-ESI-MS chromatogram by using as filter ions the m/z 739.85082 and 790.40543 relative to the [M + 2H]2+ of the native and adducted peptides, respectively.

Hb was digested with trypsin following incubation with HNE (Hb:HNE 1:10 molar ratio) for 240 min at 37°C.
MA, Michael adduct; SB, Schiff base.
Delta (ppm) with respect to the calculated [M+H]+ values.
Displays the probability of finding a match as good as or better than the observed match by chance.
Peak area (%) of the adducted peptides with respect to the peak area of the corresponding unmodified peptides.
Molecular-modeling studies were then performed with a view to explaining the different abundance of the monitored adducts. As compiled in Table 2, three parameters were considered in this computational analysis: (a) the accessibility of each considered residue, as defined by its SAS value; (b) the predicted ionization constants, as computed by pKaTool (18); and (c) the interaction energy, as derived by docking simulation for the closest pose. Table 2 clearly indicates that all monitored residues are significantly accessible, even if the His residues are, on average, more exposed than the Lys and Cys residues. Again, Table 2 shows that all residues possess a pK a constant quite similar to that of the corresponding free residue, meaning that the protein microenvironment does not vastly affect the ionization state of such residues. This result can justify the marked abundance of the Cysβ93 adduct because, at physiologic pH, a significant fraction of reactive thiolate anion is found, as well as the modest abundance of Lys adducts, because, at physiologic pH, the unreactive protonated forms are largely predominant. Additionally, the docking scores, as computed for the closest pose of each residue, are in line with the adduct abundances, showing that the complex with HNE close to Cysβ93 yields the vastly lowest interaction energy. Figure 5 depicts such minimized complex unveiling that HNE is accommodated in a polar crevice at the interface between the two subunits, where it can stabilize some significant contacts. Specifically, the aldehydic carbonyl group forms an H-bond with Hisβ97, whereas the C4 hydroxyl function stabilizes a network of H-bonds that involves Lysα40, Aspβ94, and Hisβ146. Modeling analyses also can explain why Cysβ93 is the only Cys residue yielding adducts with HNE. The other two Cys residues (Cysβ104 and Cysβ112) are markedly less acid (pK a > 10) and totally buried, as confirmed by the fact that no docking pose exists where HNE is reasonably close to such cysteines.
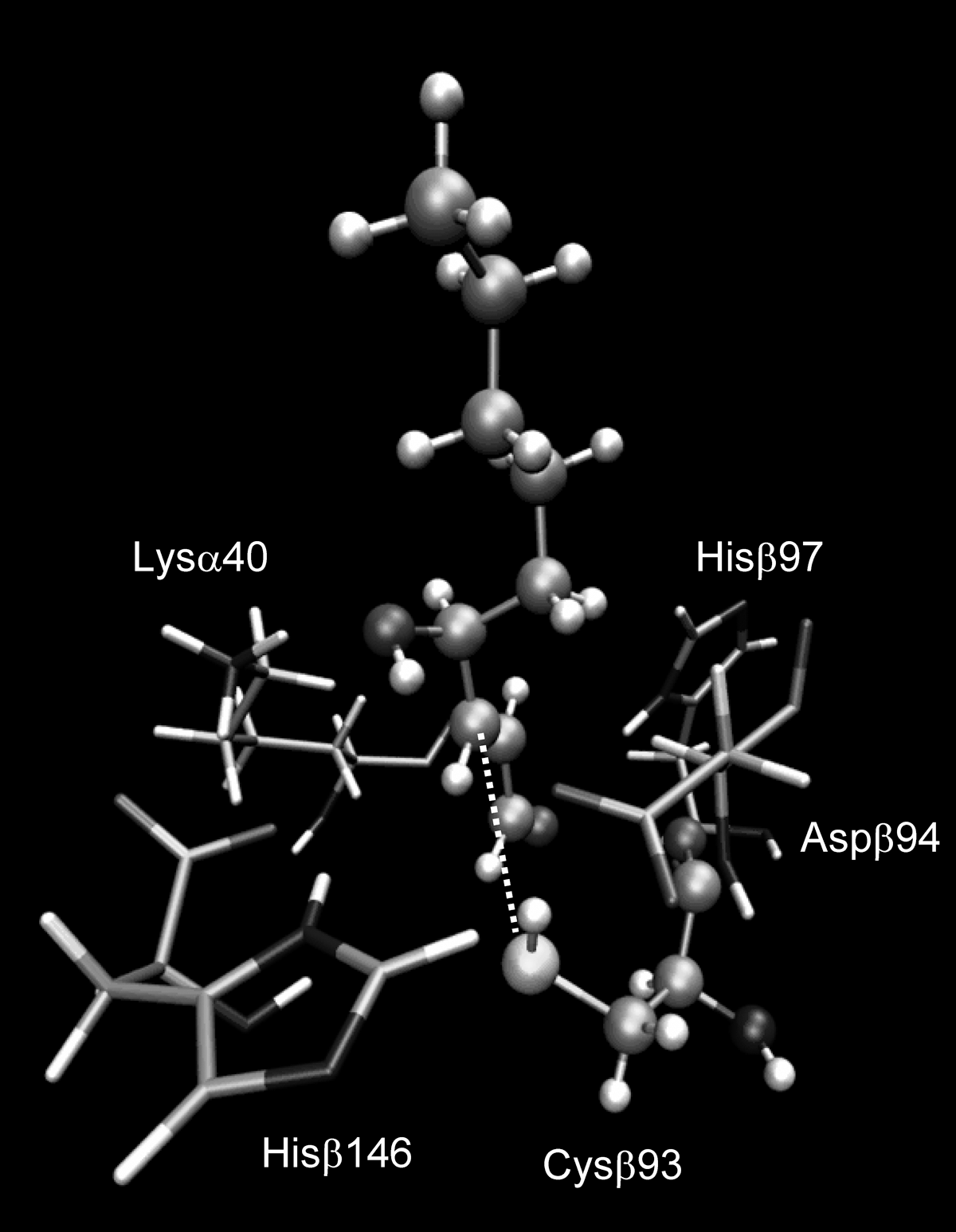
Reactivity was parameterized in terms of ionization state (expressed by predicted pK3 values), solvent accessibility (as computed by SAS surface, Å3), and interaction capacities (evaluated by docking score for the closest pose, Kcal/mol).
Peak area (%) of the adducted peptides with respect to the peak area of the corresponding unmodified peptides.
Edaravone inhibits Hb carbonylation
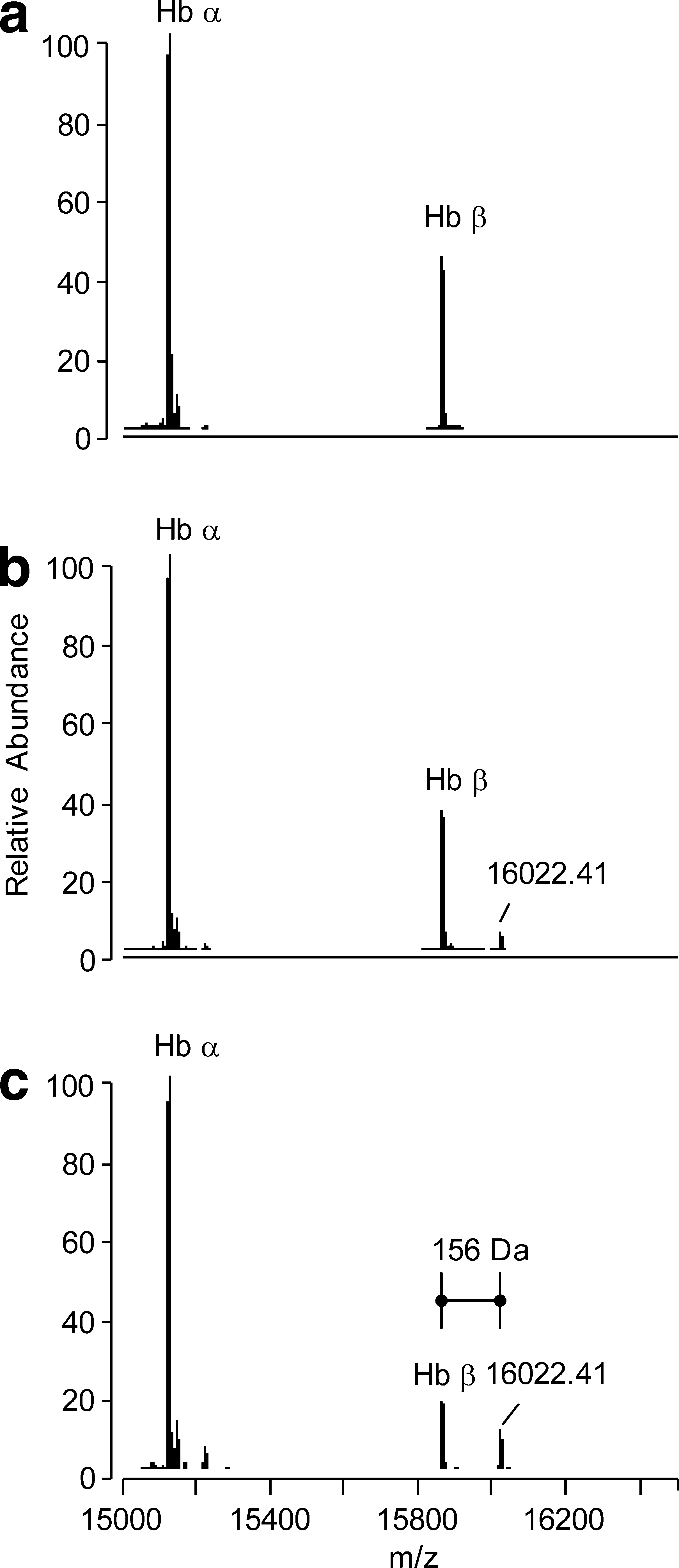
When EDA was co-incubated for 120 min in the presence of fixed concentrations of Hb and HNE (1:10 molar ratio), it was found dose-dependently to inhibit Hb carbonylation (Fig. 6). The protective effect started at 40 μM (Fig. 6b), a concentration able to reduce by 24 ± 4% the Hb-HNE adduct formation, taking as 100% formation that found in samples incubated in the absence of EDA (Fig. 6a). The protective effect dose-dependently increased, reaching a 92 ± 2% inhibition at 200 μM (Fig. 6c). The signal for the Hb-HNE adduct was undetectable in samples incubated with 400 μM EDA (100% inhibition) (Fig. 6d).

RCS-scavenging activity (reactivity)
To gain a deeper insight into the mechanism of protein-carbonylation inhibition, the direct HNE-scavenging ability of EDA was then evaluated, by measuring the consumption of the target aldehyde with HPLC analysis. Table 3 includes the scavenging activities of EDA toward HNE, and the results are compared with those of selected reference compounds (PYR, L-CAR, and HY), which are well-known RCS-sequestering agents, as already reported (28). EDA was found to be the most reactive compound toward HNE, leading to an almost total disappearance of the aldehyde after 180 min of incubation. The scavenging activity of EDA was then extended to other RCS (ACR and GO), and also with these target aldehydes, the EDA potency was far greater than that of the other sequestering agents.
Results are reported as percentage of RCS and PYAL remaining with respect to a blank incubated in the absence of the tested compound. Values are the mean ± S.D. of 4 determinations.
N.D., not determined.
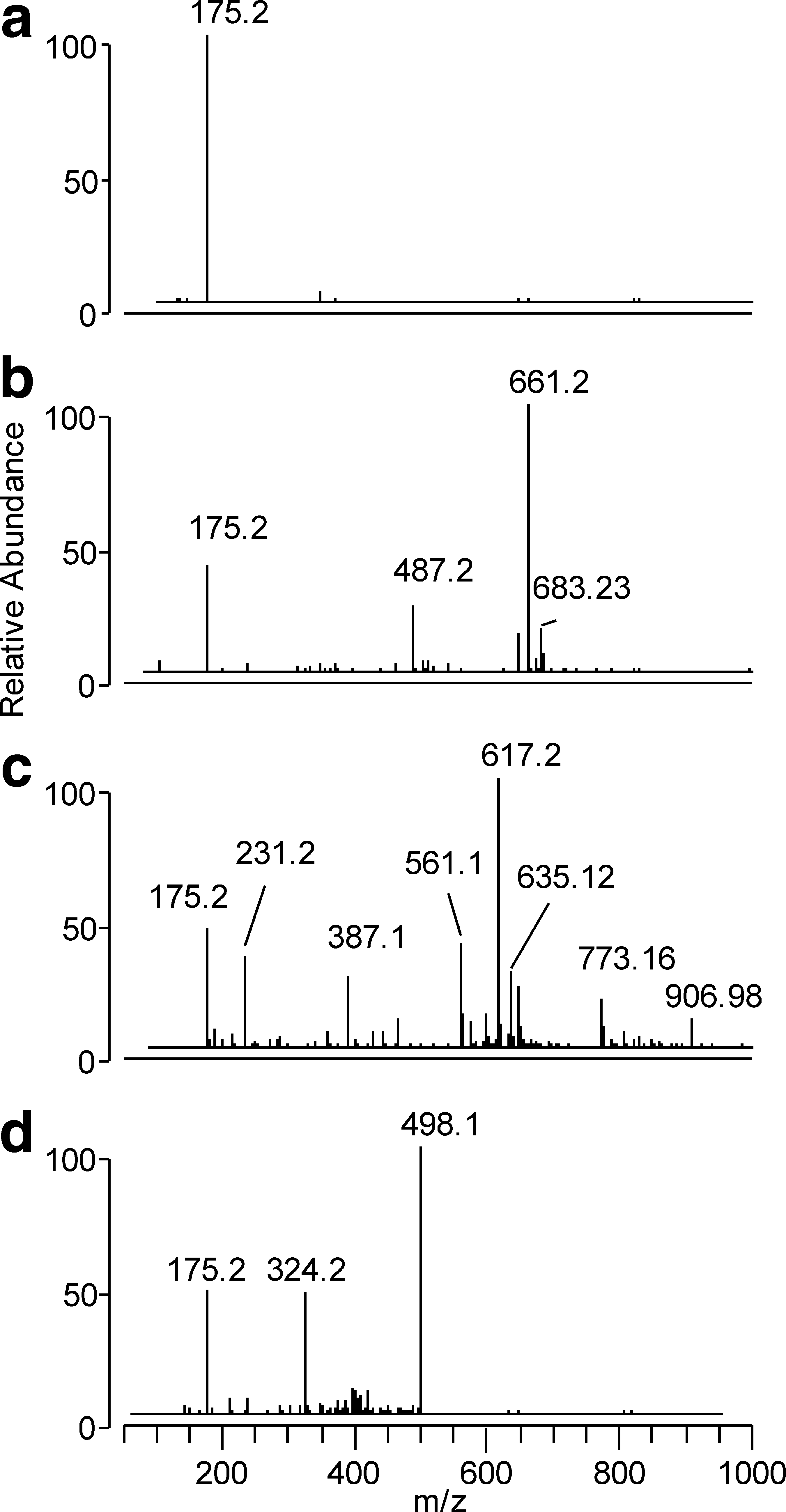
The entrapping capacity of EDA was further confirmed by identifying the RCS-EDA reaction products with direct-infusion MS analysis in full-scan mode. As a preamble, it is worth mentioning that several theoretic and experimental studies showed that the most electrophilic center in an α,β-unsaturated aldehyde is the C3 atom (13), meaning that when it reacts with a carbonion (as in the case of edaravone), the Michael addition is largely prevalent on the mixed aldolic condensation. Hence, only the reaction mechanisms starting with a Michael addition were monitored here. Figure 7a shows the MS spectrum of EDA, characterized by a base peak at m/z 175.2 relative to the [M + H]+ ion. The MS spectrum recorded after 180-min incubation was superimposable, to indicate the stability of EDA in the experimental conditions used (data not shown). When

Elemental composition of each compound was calculated by Elemental composition feature included in Xcalibur Qual Browser 2.0. Molecular softness for the monitored adducts was computed by semiempirical calculations. Notice that the molecular softness of EDA is equal to 0.116 au.
When EDA was incubated with ACR for 180 min, besides the [M + H]+ ion relative to the unreacted EDA, several other ions appeared, among which were those at m/z 617.2 (base peak), m/z 561.14 (35%), 387.1 (30%), and 231.2 (34%) (Fig. 7c). The accurate masses of the reaction products were then determined by using Orbitrap as a mass analyzer (resolution, 100,000) (Table 4). The proposed reaction mechanism of EDA with ACR is schematically summarized in Fig. 9. The first step involves formation of a Michael adduct, forming compound 1d. Compound 1d further reacts with a second EDA molecule through an aldolic condensation and dehydration, giving 2d. This intermediate, being characterized by an α,β-unsaturated moiety, reacts with another EDA molecule, yielding compound 3d, which in turn reacts with ACR to form two 4d derivatives, which differ in adduct position. Again, 4d isomers can yield a mixed aldolic condensation with a fourth EDA molecule to form 5d positional isomers.
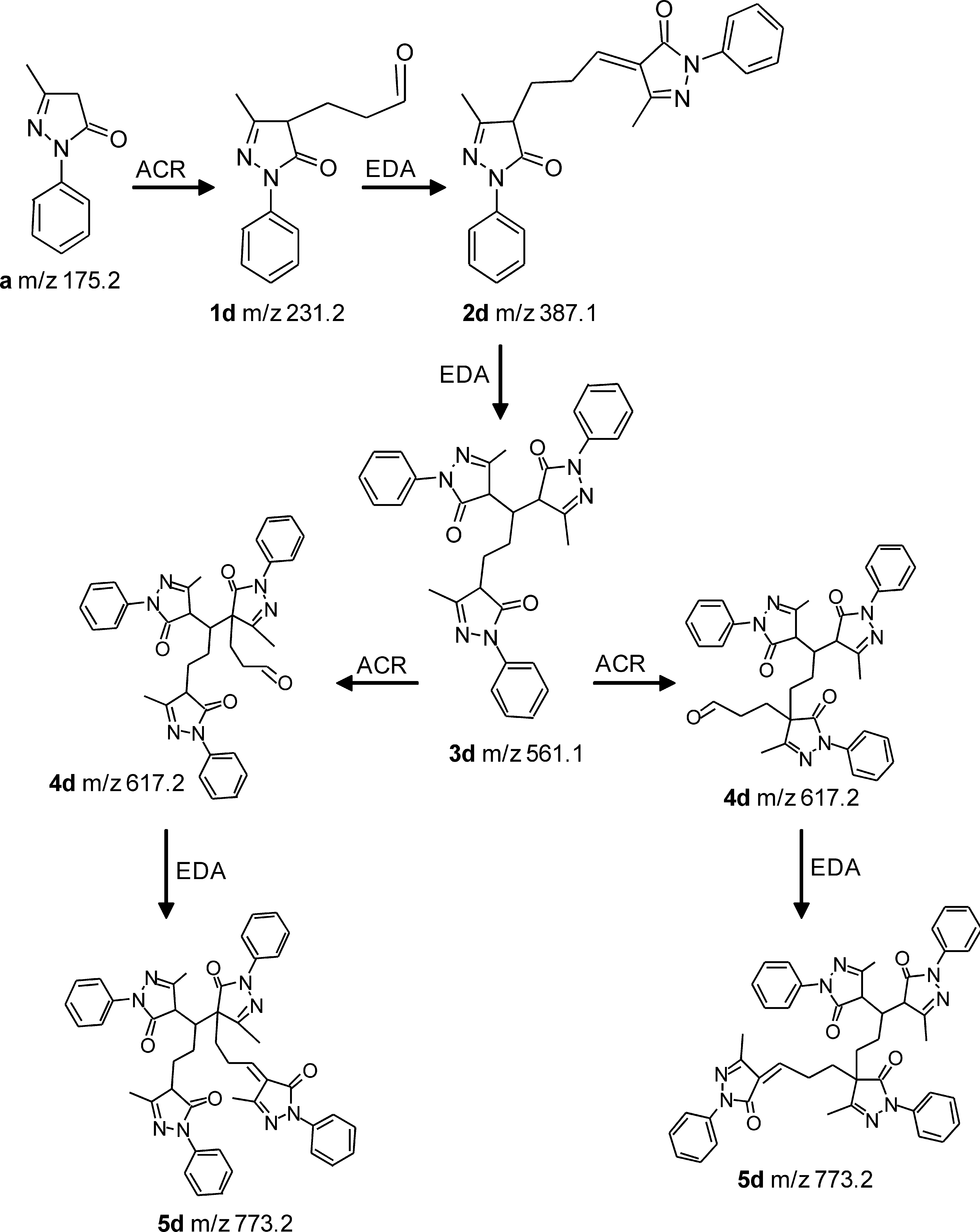
The molecular softness values for monitored adducts (Table 4) explain the particular reactivity of EDA toward α,β-unsaturated aldehydes. In particular, the molecular softness values allow two relevant considerations: (a) the adducts being characterized by α,β-unsaturated moieties are markedly more reactive than are those having an acid methylene, confirming that the Michael addition is largely preferred, compared with aldolic condensation; and (b) all considered adducts show a molecular softness similar to or greater than that of EDA. This is understandable, considering that all adducts conserve either an α,β-unsaturated moiety or an acid methylene, and suggests that EDA can react with α,β-unsaturated aldehydes, acting as a polymerizing agent and forming adducts characterized by very high molecular weights. It should be noted that local reactivity descriptors (as expressed by condensed atom Fukui indices) were computed when more adduct positions were possible to unveil whether the examined adduction could have a certain degree of regioselectivity. Nonetheless, the very similar Fukui indices for 4d and for 5d derivatives suggest that such positional isomers are almost equally probable.
Pyridoxal-scavenging activity (selectivity)
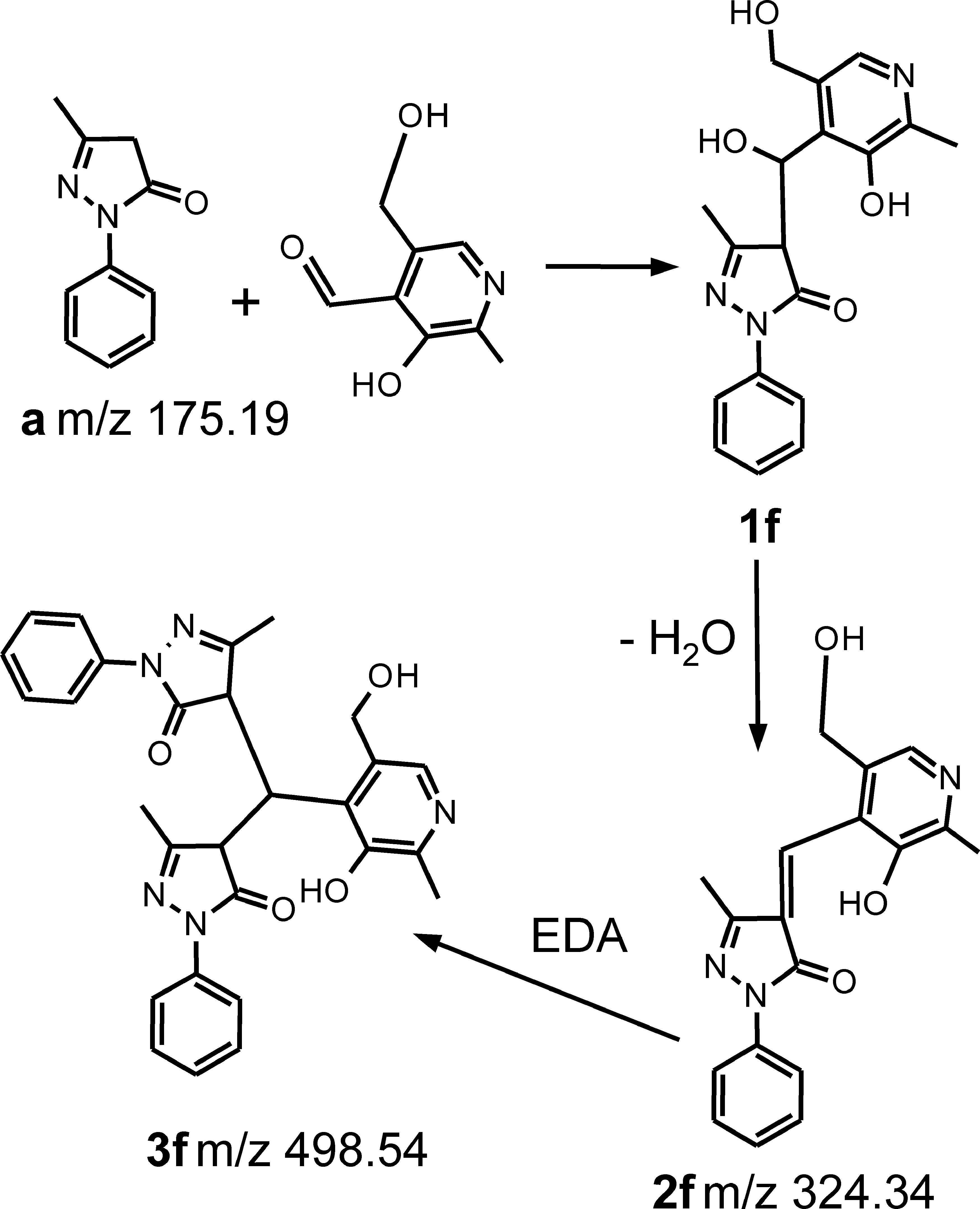
The ability of EDA, compared with that of the reference compounds, to react exclusively with cytotoxic RCS (no reaction with physiologically relevant aldehydes), was determined by using pyridoxal as target aldehyde. HPLC analyses indicated that EDA and HY, unlike L-CAR, induce a significant depletion of pyridoxal, after 180-min incubation (Table 2). The lack of selectivity of EDA was further confirmed with MS analysis of the incubation mixtures. The ESI-MS spectra are indeed characterized by the reduction of the [M + H]+ signal relative to the scavenger at m/z 175.2, accompanied by the formation of the reaction products at m/z 498.15 and 324.2 (Fig. 7d). Figure 10 summarizes the proposed EDA-PYAL reaction mechanism, involving as a first step a mixed aldolic condensation to yield 1f. Dehydration of this intermediate gives 2f, which, bearing an α,β-unsaturated moiety, reacts with a second EDA molecule to give 3f.
Discussion
In view of the growing need to find new molecules sharing both antioxidant and carbonyl scavenger properties to restrain lipid-derived oxidative/carbonyl stress associated with a variety of pathologic conditions, we focus our study on edaravone, a compound already known as a potent radical scavenger, whose carbonyl-sequestering ability, however, has been poorly investigated. The only report available limited the evaluation to the trapping ability toward glyoxal and methylglyoxal, dialdehydes that are precursors of EAGLEs and AGEs, respectively (14). Edaravone has been shown to be effective in a variety of oxidative stress–related diseases owing to its antioxidant potency. Because intervention strategies based on a direct radical-scavenging (antioxidant) approach, which provides a first line of defense against free radicals, such as vitamins E and C, failed to show beneficial effects in ROS-dependent diseases (2), we thought that edaravone intervention as a free radical scavenger could be questionable, and not solely accountable for the beneficial effects demonstrated in both pharmacologic and clinical studies (29). This consideration, associated with the emerging role of RCS derived by ROS-induced decomposition of polyunsaturated fatty acids as pathogenic factors, and to the concept of carbonylation damage as a new drug target (1, 2, 17), prompted us to investigate in detail the RCS-entrapping ability and potency of EDA.
With Hb as a protein model, the results of the first part of this study demonstrate that EDA inhibits Hb carbonylation induced by HNE through a direct trapping mechanism. The covalent modification of Hb by HNE, never reported, involves Cys93 of the β chain, as demonstrated by a combined high-resolution mass-spectrometric and computational approach. Our results are quite different from those reported by Yocum et al. (31), who studied the covalent modifications of Hb after incubation with 4-oxo-nonenal as a lipid-derived target aldehyde, by using MALDI-TOF/MS analysis of the intact protein and a combination of LC-ESI-MS/MS and MALDI-TOF/MS/MS analysis of the tryptic peptides. Although incubation conditions were different from those reported in this study, they found covalent modifications on both Hb chains and identified Hys20 (α chain) and His63 (β chain) as the target nucleophilic sites for 4-oxo-nonenal adduction, a finding supported by molecular modeling, indicating these two residues as the most solvent accessible in intact Hb.
Nonetheless, it should be noted that our modeling analyses confirmed the marked exposition of His20, but unveiled that His63 is quite inaccessible (SAS = 7.2 Å2), thus explaining why the adduct on His63 was not detected in the present study.
Our findings indicate that Cysβ93 Hb modifications can be used not only as a reliable in vitro model to screen rapidly the ability of newly developed RCS sequestering agents to inhibit protein carbonylation, but also as a potential biomarker in vivo of lipid-derived macromolecule damage. The ability of EDA to prevent Hb modification by HNE, the prototype of cytotoxic α,β-unsaturated aldehydes derived from lipid peroxidation, served as the first convincing evidence to undertake a detailed investigation of the RCS-entrapping capacity of the compound, providing new information on its reactivity (a ranking of potency within a series of well-known RCS-sequestering agents has been established), its ability to spare physiologically relevant aldehydes, such as pyridoxal (selectivity), and on the mechanisms involved in EDA interaction with both α,β-unsaturated aldehydes and pyridoxal.
The results of this part of the study confirm that EDA traps glyoxal and demonstrate for the first time that the compound, unlike
The high reactivity with α,β-unsaturated aldehydes (EDA is far more potent than the other structurally unrelated RCS scavengers considered) and the ability to inhibit protein carbonylation shed light on a new mechanism by which the compound can exert its efficacy in vivo and open new perspectives to start a rational design of EDA analogues, maintaining the same RCS-scavenging potency, but characterized by an increased selectivity.
Footnotes
Acknowledgments
This research was supported by funds from University of Milan (PUR 2007, 2008) and from MIUR (PRIN 2007).
Author Disclosure Statement
No competing financial interests exist.