
Abstract
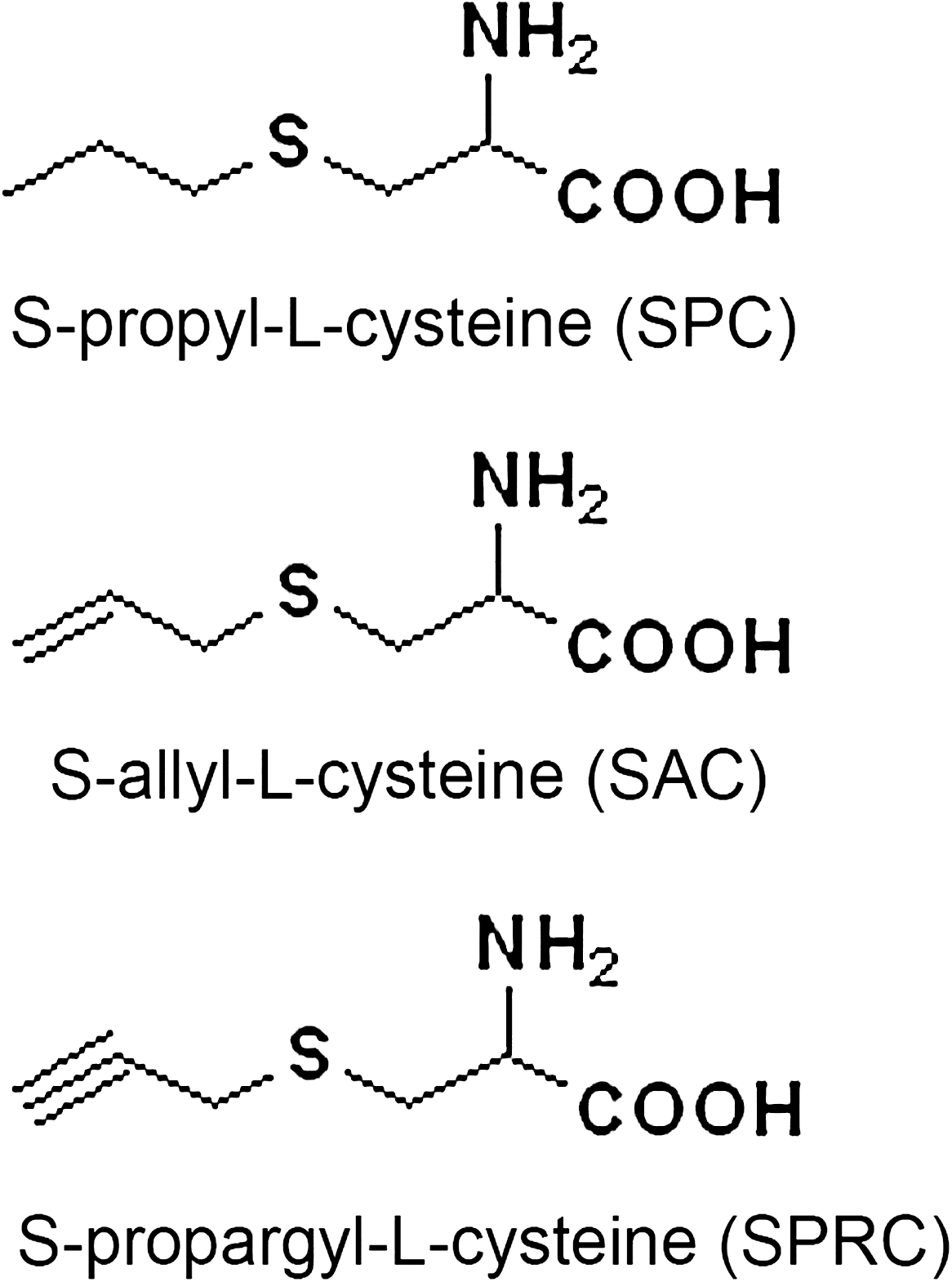
The current study was designed to evaluate the pharmacologic effects of three novel cysteine-containing compounds: S-propyl-
Introduction
Garlic (Allium sativum) has been prescribed for the treatment of several human diseases for centuries. Recent interest has focused on the role of garlic and its chemical constituents in the prevention of cardiovascular disease and cancer (14, 15). The mechanisms ascribed to garlic's biologic activities are believed to be partly due to its antioxidant action, although additional mechanisms also are likely to contribute (4). Recent work has shown that the cardioprotective effects of garlic may be attributed to its ability to modulate the endogenous levels of hydrogen sulfide (H2S). Interestingly, H2S was recently identified as a novel gaseous signaling molecule having a similar function to that of nitric oxide and carbon monoxide in the cardiovascular system of mammals (1, 25). Wei et al. (22) reported that H2S acts as an antioxidant by attenuating oxidative stress in hypoxic pulmonary hypertension. Geng et al. (7) also proposed that H2S protects the heart from isoproterenol-induced ischemic injury, at least in part by scavenging oxygen-free radicals and attenuating lipid peroxidation. Similarly, H2S has been shown to protect primary rat cortical neurons from oxidative injury by stimulating the synthesis of the antioxidant glutathione (9). In the heart, H2S is produced by the pyridoxal-5-phosphate–dependent enzyme, cystathionine-γ-lyase (CSE) with
Materials and Methods
All animal care and experimental protocols complied with the Animal Management Rules of local authorities and Care and Use of the Laboratory Animals of the Experimental Animal Center of Fudan University.
Materials
SAC (99%) was supplied by Wakunaga Pharmaceutical Co. (Hiroshima, Japan); cysteine (99.5%) was purchased from Hengbai Chemical Co. (Shanghai, China). Propyl bromide and propargyl bromide were purchased from Yancheng Kelida Chemical Co. (Zhejiang, China). PrimeScript 1st Strand cDNA Synthesis Kits were from TaKaRa (Dalian, China).
Synthesis of SPC and SPRC
SPC was synthesized from
Treatment protocols
Adult male Sprague–Dawley rats (200 to 250 g) were randomly assigned into five treatment groups: sham vehicle (saline) (n = 6), MI treated with vehicle (n = 10), MI treated with SPC (50 mg/kg/day) (n = 10), MI treated with SAC (50 mg/kg/day) (n = 10), and MI treated with SPRC (50 mg/kg/day) (n = 10). In addition, five separate groups (n = 6) representing each treatment group as described earlier were treated with the cystathionine-γ-lyase (CSE) inhibitor, propargylglycine (PAG, 10 mg/kg/day).
Experimental induction of acute myocardial infarction
Rats were injected intraperitoneally on a daily basis for 7 days before occlusion of the coronary artery to induce MI. The MI model was induced on day 7 by ligating the coronary artery, as previously described (27). After the surgery, all treatment regimens were continued for an additional 2 days until the animals were killed 48 h after surgery. Hearts were also excised, washed in ice-cold saline, and stored at −80°C for subsequent experimental assays.
Glutathione levels and antioxidant enzyme analysis
Oxidized glutathione (GSSG) and reduced glutathione (GSH) levels were measured by using a commercially available kit according to the manufacturer's instructions (Beytime Institute of Biotechnology, Nantong, China). For the determination of antioxidant enzyme activities, 0.05 g LV tissues were homogenized in 50 mM ice-cold potassium phosphate buffer (pH 6.8). Superoxide dismutase (SOD) activity was measured as described by Dieterich et al. (6). In brief, total SOD activity was determined by monitoring the inhibition of pyrogallol autooxidation at 420 nm. To measure Mn-SOD activity, Cu Zn-SOD was inhibited by using diethyldithiocarbamic acid. SOD activity is expressed as units per milligram protein, where 1 U is the amount required to inhibit pyrogallol oxidation by 50%. GPx and catalase (CAT) activities were determined by the modified method of Alvarez (2). Lipid peroxidation was measured in terms of tissue MDA contents by using the thiobarbituric acid (TBA) assay, with 1,1,3,3-tetramethoxypropane as an external standard for the construction of standard curves. MDA levels are expressed as nanomoles MDA per milligram protein (24). Protein content was determined by using a BCA protein assay kit (Beytime Institute of Biotechnology).
Measurement of CSE activity
CSE activity was assayed as described (26). In brief, 0.1 g of left ventricular myocardial tissue was thawed on ice and homogenized in 2 ml of 100 mM ice-cold potassium phosphate buffer (pH 7.4). Tissue homogenates were centrifuged at 24,000 g for 5 min at 4°C, and the supernatant was used for the assay. The reaction mixture contained 20 μl of 10 mM
Real-time PCR assay
Total RNA from either the left ventricle (n = 4 each group) or cultured cardiomyocytes was isolated by using Trizol reagent (Invitrogen, Carlsbad, CA). RNA was quantified spectrophotometrically by measuring the optical density of samples at 260/280 nm. Two micrograms of RNA were reverse-transcribed to cDNA with the PrimeScript 1st Strand cDNA Synthesis Kit (TaKaRa). Gene expression was performed by using real-time PCR with the SYBR Green PCR Master Mix (Bio-Rad) on a BIO-RAD IQ5 system. Expressions of SOD1, SOD2, CSE, GPx, and CAT mRNA were determined. Results were normalized to β-actin. Three duplicates were performed. The primers used for real-time PCR are shown in Table 1.
Western blot analysis
Frozen tissues (n = 4 per group) were cut into small pieces, homogenized in 0.5 ml of RIPA buffer (150 mM NaCl, 1% NP-40, 0.5% deoxycholate, 0.1% sodium dodecylsulfate, 50 mM Tris-hydrochloric acid, 2 mM phenylmethylsulfonyl fluoride, pH 7.4), and incubated at 4°C overnight. The dissolved proteins were collected after centrifugation at 10,000 g for 30 min, and the supernatant was then collected. Protein concentrations were determined by using the enhanced BCA protein assay kit (Beytime Biotechnology). To detect CSE, CuZn-SOD, and Mn-SOD protein expression levels, cell lysates were subjected to SDS-PAGE analysis on a 12% (wt/vol) acrylamide gel and then electrotransferred onto a PVDF membrane (Millipore Corporation). Membranes were then incubated with the appropriate secondary horseradish peroxidase–conjugated anti-rabbit IgG antibodies at a 1:10,000 dilution (Jackson ImmunoResearch Laboratories Inc., West Grove, PA). Immunoreactive proteins were visualized by using the ECL Western blotting detection kit (Alpha Innotech, San Leandro, CA).
Histologic analysis and immunofluorescence assays
Heart tissues (2 mm) were embedded in OCT medium immediately. Frozen sections (6 μm thick) were used for hematoxylin-eosin staining and immunofluorescence assays.
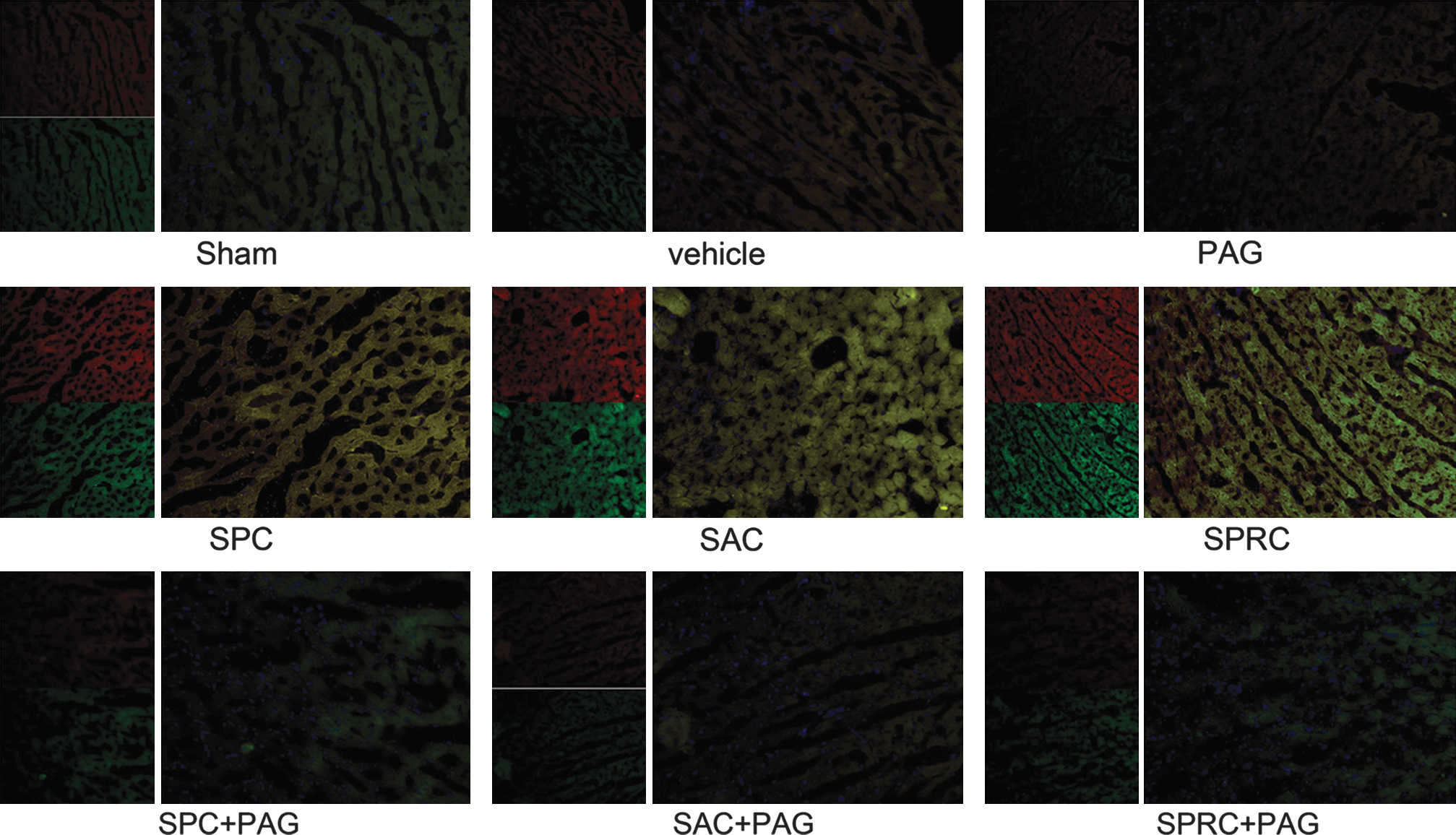
By using antibodies for CSE and Mn-SOD, we performed immunofluorescence assays to measure the relation with the endogenous H2S production and the expression of the antioxidant enzyme Mn-SOD. For immunofluorescence assays, sections were washed (0.1 M Tris, pH 7.6, 15 min), denatured (2N HCl, 37°C, 30 min), rinsed (0.1 M PBS, 10 min), incubated with 1% H2O2 in 0.1 M Tris for 30 min, rinsed, blocked (10% normal goat serum, 37°C, 30 min), and incubated with anti-CSE antibody and anti-MnSOD antibody (4°C overnight). Sections were rinsed (0.1 M PBS, 10 min), incubated with tetraethyl rhodamine isothiocyanate (TRITC)-conjugated goat anti-rabbit IgG and fluorescein isothiocyanate (FITC)-conjugated anti-mouse IgM (room temperature, 2 h), and rinsed, mounted, dried, and coverslipped by using DAPI.
Light-microscopy and electron-microscopy observation
For transmission electron microscopic study, specimens were prefixed in 2.5% glutaraldehyde solution, diced into 1 mm3, followed by three 15-min rinses with 0.1 M phosphate buffer (pH 7.4). Postfixation was in cold 1% aqueous osmium tetroxide for 1 h. After a rinse with phosphate buffer again, the specimens were dehydrated in a graded ethanol series of 50 to 100% and then embedded in Epon 812. Ultra-thin sections were sliced with glass knives on a LKB-V ultramicrotome (Nova, Trelleborg, Sweden), stained with uranyl acetate and lead citrate, and examined under a Hitachi H-600 electron microscope.
Statistical analysis
All values are presented as mean and standard deviations. One-way analysis of variance (ANOVA) was used to examine statistical comparisons between groups. The statistical significance of differences between two groups was determined by using a two-tailed Student's t test. A χ2 (chi-square) test was used for calculating the significance of the mortality data. A probability value of <0.05 was taken to indicate statistical significance. All analyses were performed by using SPSS 12.0.
Results
GSH and GSSG levels
The GSH content of left ventricular tissues of rats in the MI vehicle group were lower than those of the sham group (9.5 ± 0.7 μM/mg protein vs. 24.3 ± 1.6 μM/mg protein, p < 0.01). In rats treated with the three cysteine derivatives, GSH levels were preserved. In the SPC-, SAC-, and SPRC-treated animals, GSH levels were increased by 67%, 77%, and 80%, as compared with the MI vehicle group (p < 0.05). Interestingly, in the MI + PAG group, the GSH level was reduced to 7.8 ± 0.7 μM/mg protein. PAG also was found to abolish the observed increases in GSH levels in the SPC-, SAC-, and SPRC-treated groups (Fig. 2A). In addition, GSSG levels in the MI vehicle rats were higher than those in the sham-treated animals (p < 0.05), and all three cysteine analogues tested in the current study were found to decrease GSSG levels in MI animals (p < 0.05). The decrease in GSSG levels in the cysteine analogues–treated animals is suggestive of a reduction in oxidative stress.
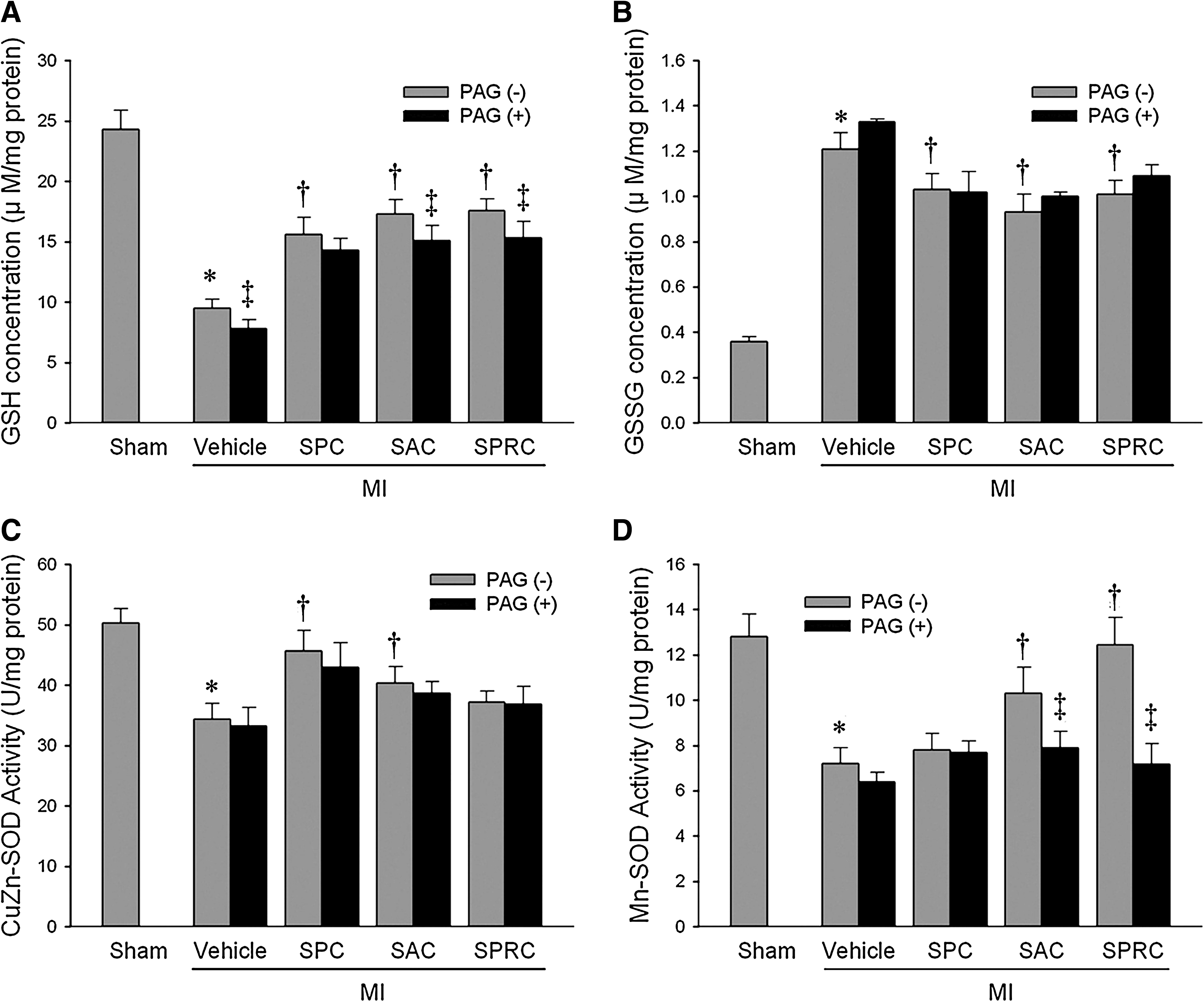

Antioxidant enzyme activities and MDA formation in MI tissues
The activities of tissue antioxidant enzymes are shown in Table 2. In our study, all three cysteine derivatives were found to increase the enzymatic activities of GPx as compared with the MI vehicle group. Similarly, the free radical–scavenging enzymes CuZn-SOD and Mn-SOD also were preserved. In the SPC group, CuZn-SOD activity was increased to 45.7 ± 3.4 U/mg protein, as compared with the MI vehicle group of 34.4 ± 2.6 U/mg protein (Fig. 2C). However, no noticeable changes were observed for Mn-SOD. In contrast, SAC increased both the Cu-Zn SOD activity to 40.3 ± 2.8 U/mg protein and the Mn-SOD activity to 10.32 ± 1.12 U/mg protein as compared with the MI vehicle group of 34.4 ± 2.6 and 7.21 ± 0.68 U/mg protein, respectively (p < 0.05). Interestingly, in the SPRC-treated group, only Mn-SOD was induced to 12.45 ± 1.20 U/mg protein and was significantly different from the MI vehicle group (p < 0.01; Fig. 2D).
Data are shown as mean values ± SD of five animals per group.
p < 0.05 when compared with the sham group.
p < 0.05 when compared with the MI group.
p < 0.05 when compared with the PAG (-) group.
In the PAG-treatment groups, Mn-SOD activity was significantly decreased to 7.68 ± 0.52 U/mg protein (SPC + PAG), 7.89 ± 0.73 U/mg protein (SAC + PAG), and 7.18 ± 0.9 U/mg protein in the SPRC + PAG group (p < 0.05). No significant changes in CuZn-SOD activity were observed for any of the cysteine analogues treated with PAG.
As summarized in Table 2, the antioxidant enzyme activities of GPx and CAT in left ventricular homogenates were increased in the SPC- and SPRC-treated groups but not in the SAC group. To support these findings, we determined the tissue levels of MDA, an additional biomarker of oxidative stress. In this study, MDA levels also were found to be increased in the MI vehicle group as compared with the sham group (p < 0.01). However, treatment with SPC, SAC, and SPRC reduced MDA levels.
Left ventricular CSE activity in rats after MI injury
Left ventricular CSE activity was analyzed in the tissue homogenates from all treatment groups (n = 4). SPC, SAC, and SPRC increased CSE activity by 1.1-fold, 1.4-fold, and 1.6-fold, respectively, as compared with the MI vehicle group (1.76 ± 0.12 μmol/g protein/h, 2.19 ± 0.22 μmol/g protein/h, and 2.48 ± 0.12 μmol/g protein/h vs. 1.58 ± 0.17 μmol/g protein/h; p < 0.01) (Fig. 2E). SPRC-treated animals had the highest CSE activity of all three of the cysteine derivatives tested. The PAG-treated group had the lowest CSE activity of 0.78 ± 0.05 μmol/g protein/h. In the SPC + PAG (0.89 ± 0.06 μmol/g protein/h), SAC + PAG- (1.21 ± 0.09 μmol/g protein/h), and SPRC + PAG (1.34 ± 0.09 μmol/g protein/h)-treated groups, the CSE activities were significantly lower as compared with those in the SPC-, SAC-, and SPRC-treated groups alone (p < 0.05) (Fig. 2E).
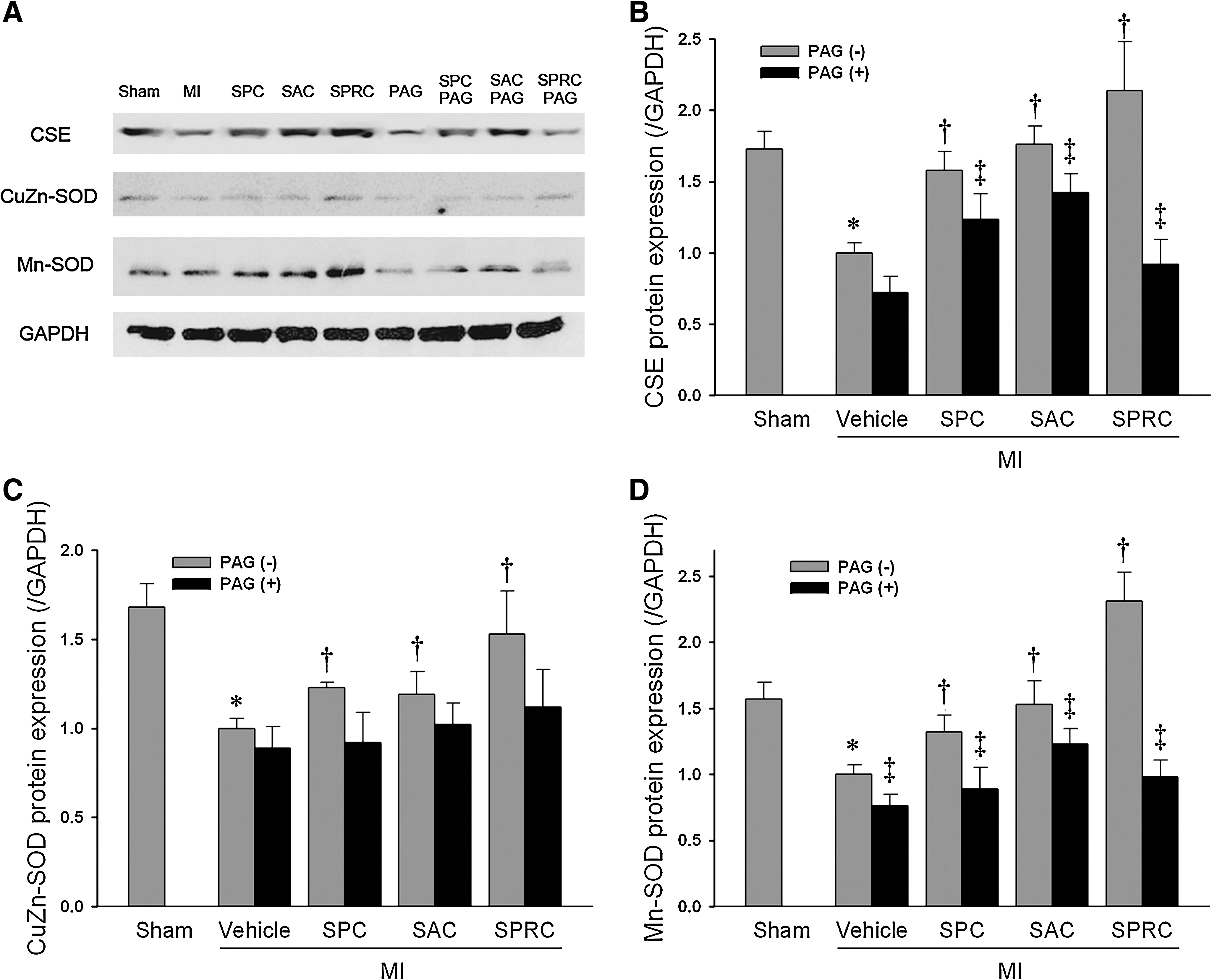
Immunohistochemical analysis of CSE and Mn-SOD contents in heart tissues of rats
As shown in Fig. 3, the CSE protein is stained red, and Mn-SOD protein is stained green. Myocardial nuclei are shown in blue after staining with DAPI. The CSE protein content of the MI vehicle group and PAG-intervened groups was lower than that in the SPC, SAC, and SPRC groups. Similar findings also were found for the tissue levels for the antioxidant defensive enzyme, Mn-SOD.
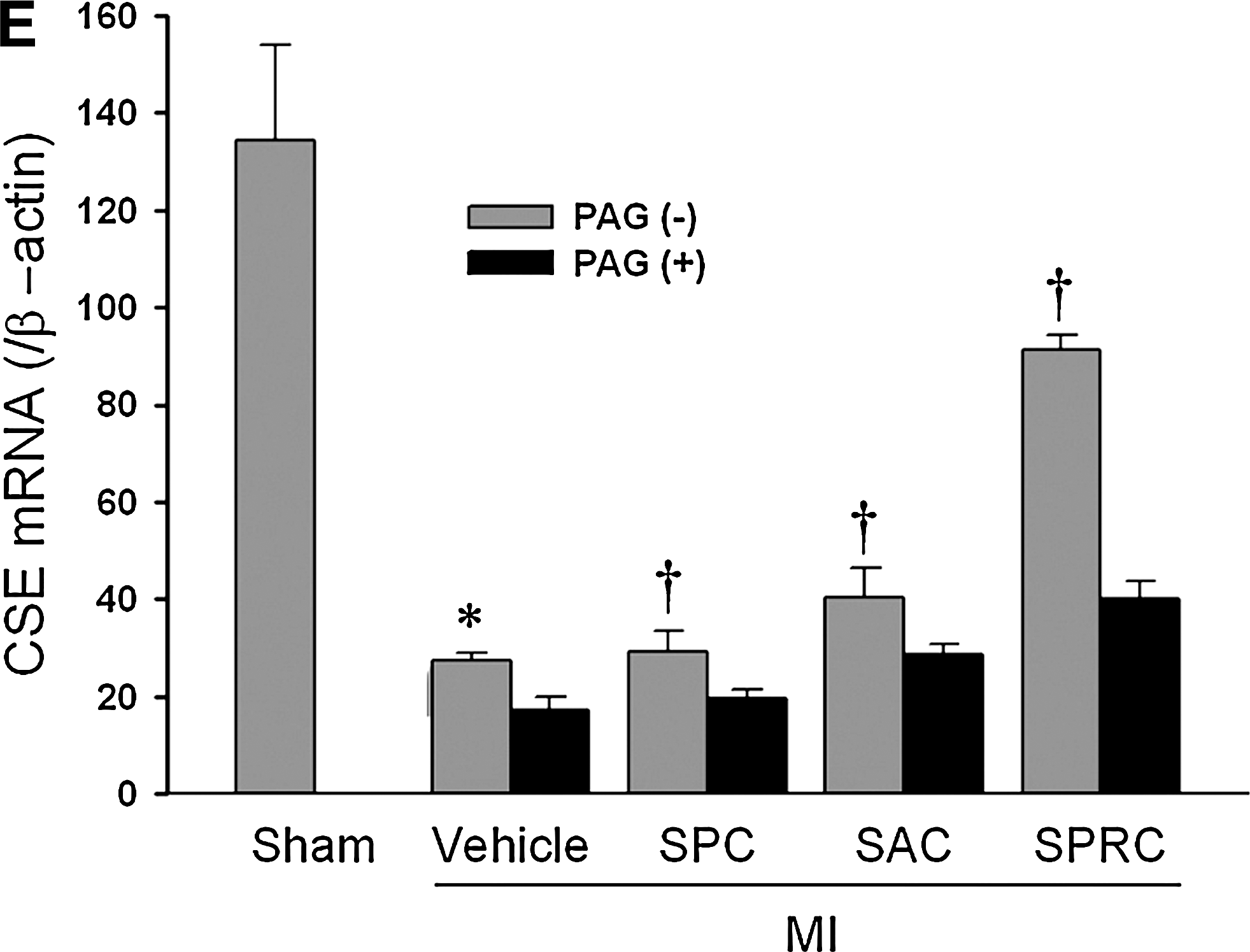
Oxidative enzyme and CSE mRNA expression
SPC, SAC, and SPRC increased CuZn-SOD expression as compared with the MI vehicle group (1.44-fold, 1.42-fold, and 1.31-fold of the MI vehicle group; p < 0.05; Fig. 4A). No significant changes were found between the cysteine analogues–treated groups with PAG and without PAG in CuZn-SOD mRNA expression.
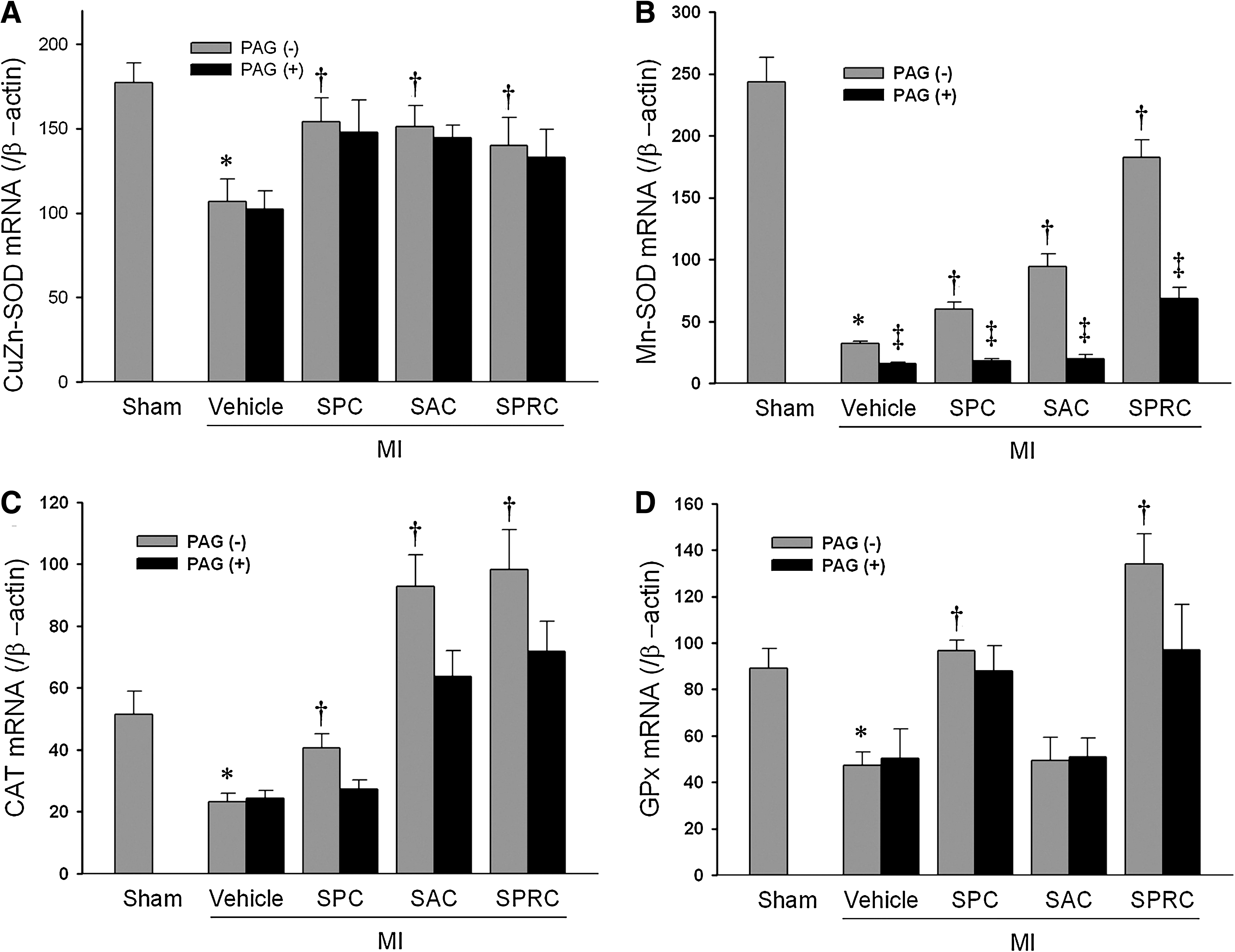
The mRNA ratios of Mn-SOD/β-actin were increased in SPC-, SAC-, and SPRC-treated groups (1.81-fold, 2.93-fold, and 5.7-fold of the MI vehicle group, respectively; p < 0.05; Fig. 4B). After PAG was added, the groups also had a lower Mn-SOD mRNA expression.
Similar changes in the expression of CSE mRNA were found in MI rats that were treated with SPC, SAC, and SPRC (1.1-fold, 1.5-fold, and 3.3-fold vs. MI vehicle rats; p < 0.01). PAG inhibited mRNA expression of CSE by 0.63-fold versus MI vehicle (p < 0.01). In the SPC + PAG-, SAC + PAG-, and SPRC + PAG-treated groups CSE mRNA expression was significantly reduced (0.66-fold, 0.7-fold, and 0.43-fold vs. SPC, SAC, and SPRC; p < 0.05) (Fig. 4A and C).
Western blotting
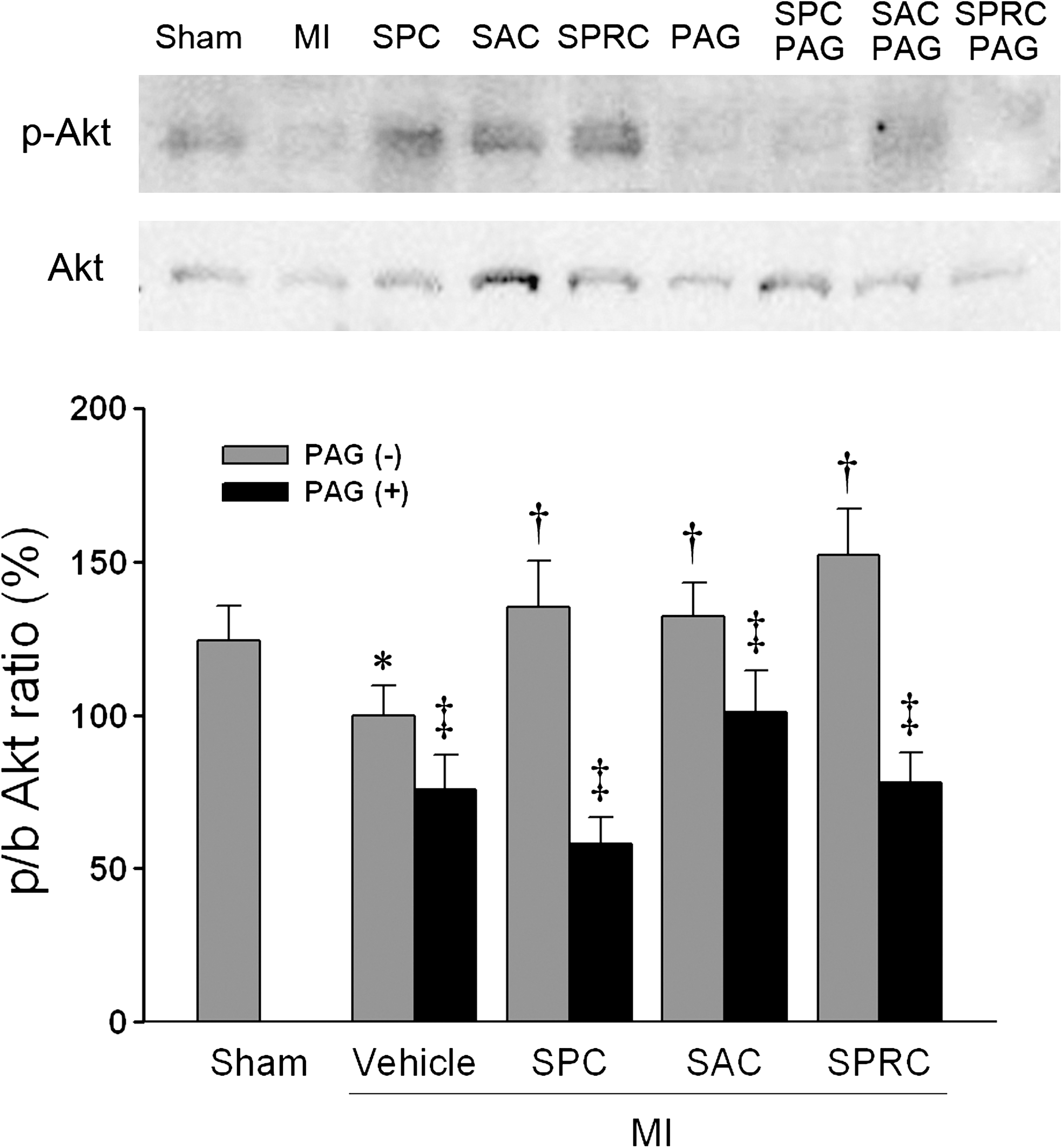
For all groups, the MI vehicle-treated rats had significantly lower CSE protein and mRNA expression as compared with the sham group. The SPC-, SAC-, and SPRC-treated groups showed a significant increase in CSE protein expression when compared with the MI vehicle group (1.57-fold, 1.76, and 2.14-fold vs. MI vehicle group; p < 0.05). Furthermore, SPC-, SAC-, and SPRC-treated groups were found to have a significant upregulation of CuZn-SOD and Mn-SOD protein. In contrast, PAG-only abolished the increase Mn-SOD protein expression comparing those without PAG pretreatment groups, respectively (p < 0.05; Fig. 5A, C, and D). We also determined the relative expression levels of the serine/threonine kinase Akt. SPC, SAC, and SPRC increased the phosphor-Akt/total Akt as compared with the MI vehicle group (1.41-fold, 1.36-fold, and 1.52-fold vs. the MI vehicle group; p < 0.05; Fig. 6). However, PAG inhibited the increase in phosphor-Akt/total Akt induced by three cysteine analogues.
Pathologic observations
Figures 7 and 8 present MI-induced pathologic changes in rat myocardium. The architecture of the myocardium was intact with regular myofiber arrangement in the sham-group rats. In hearts of rats treated with MI, disorganizations of cell structure and loss of adherence between cardiomyocytes were pronounced. The myocardial damage included enlarged cells with enlarged and often bizarre-shaped nuclei, occasional cytoplasmic vacuolization and partial degenerative muscle fibers. At the PAG-treated group, disarray of myocardial fibers and the degenerative muscle fibers with myocytolysis were the most prominent features. Cardiac ultrastructure was normal with intact and abundant mitochondria in hearts of the sham rats, whereas most mitochondria were seriously swelling in the hearts of rats operated on with MI. Moreover, the swelling of interstices among mitochondria and some mitochondria with ruptured outer membrane were observed as well. The SPC group showed a bubble in the nuclei, but the structure of mitochondria looked more normal than those MI vehicle group. The pathologic morphology of cardiac cells in SPRC group was almost same as that in the sham group, and the SAC-treated group were better than the MI group, especially in mitochondrial structure.

the degenerative muscle fibers. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article at

the abnormal nuclei with fragments;  (white) the bubble in the nucleus in the SPC-treated group.
(white) the bubble in the nucleus in the SPC-treated group.
Discussion
Garlic (Allium sativum) is believed to be useful for disease prevention. The beneficial effects of garlic have been ascribed to its potent antioxidant action (16). SAC is one of the major organosulfur compounds found in aged garlic extracts, and the allyl moiety appears to be crucial for any biologic activity. Similarly, SPC has been identified in extracts prepared from Allium species such as garlic and onion (10, 19). Several studies have indicated that SAC and SPC can inhibit TG and cholesterol biosynthesis in cultured rat hepatocytes (8, 11). SPRC, a chemical species containing a propargyl structure, also was reported to have anticholesterolemic activity (13). We recently showed that SPRC and SAC have cardioprotective effects in MI by reducing the mortality rate as well as by reducing the infarct size in rats, and we proposed a role for the H2S-associated pathway in mediating this protection (21). In this article, the infarct size/total area of the myocardium also was significantly reduced in rats subjected to SPC treatment, as compared with the MI vehicle rats by 26.1 ± 1.6% vs. 36.2 ± 1.3%, respectively (p < 0.05).
To expand on our previous studies, we evaluated the biologic effects of three cysteine analogues in a rodent model of MI. We also found that SPC, SAC, and SPRC displayed cardioprotective effects by reducing infarct size and by reducing LDH and CK leakage from cells (markers of cell-membrane integrity). H2S is a strong reducing agent and may readily react with labile molecules, particularly those derived from reactive oxygen and nitrogen species (7, 23). Interestingly, in this study, all three compounds were found to preserve tissue GSH levels and to reduce plasma MDA levels as compared with the MI vehicle group. This observation correlated with a committed upregulation of the enzyme CSE and with the induction of the cardioprotective enzymes SOD and GPx in myocardial tissues. Banerjee and colleagues (3) previously reported that rats fed with raw garlic homogenates showed a significant preservation of myocardial superoxide dismutase (SOD) activity and an improvement in myocardial morphology after MI induction with isoproterenol. In our study, we found that SPC preserved the activity of cytosolic copper/zinc SOD (CuZn-SOD); the predominant SOD in mammals. SPRC improved manganese SOD (Mn-SOD) activities, and SAC was found to increase both Cu-Zn SOD and Mn-SOD activities in MI tissues. In addition, each compound was found to maintain tissue GSH levels after the induction in MI. These findings correlate with a reduction in the levels of MI injury. With the use of immunofluorescence assays, we can see the positive correlation of expression of CSE and Mn-SOD (Fig. 3). It suggested the production of endogenous H2S might increase the protein expression of the antioxidant enzyme Mn-SOD. SPRC involved the CSE/H2S pathway may be a means of activating Mn-SOD.
In a separate set of experiments, we also evaluated whether the cysteine analogues used in the current study activated the Akt signaling pathway. Akt is a serine/threonine kinase, and a number of studies have reported that the activity of this protein can be regulated through redox stress (20). The Akt protein, once activated, can phosphorylate a wide range of intracellular substrates that regulate growth, metabolism, and survival (5). The Akt pathway is activated not only by various growth factors and cytokines but also by G protein–coupled receptor agonists, such as bradykinin, through transactivation of receptor tyrosine kinase or activation of nonreceptor tyrosine kinase, such as Src, in a ROS-dependent manner (17). In our study, we found increased expression of phospho-Akt in these cysteine-containing compounds–treated groups. But the SPRC-treated group had the more significant Akt-signaling pathway. PAG may abolish this redox-sensitive signaling in all PAG-intervention groups.
In conclusion, we report that the novel cysteine analogues SPC, SAC, and SPRC enhance cellular antioxidant defenses in rats with MI. SPRC might have a higher selection for CSE. We hypothesize that SPRC perhaps acts as an H2S donor and that the H2S released might participate in activating signaling cascades associated with the prevention of oxidative stress in MI. Collectively, the ability of the CSE/H2S pathway to alter the oxidative condition suggests that the modulation of CSE expression and H2S production may provide a novel therapeutic avenue for the treatment of ischemic cardiac diseases.
Contribution of the Authors
Q. Wang is the main person who conducted the study; X.L. Wang and H.R. Liu assisted in the experiments. P. Rose was involved in scientific discussion of the study, and Y.Z. Zhu is the PI for this project.
Footnotes
Acknowledgments
The current study was supported mainly by research grants from the National Natural Science Foundation of China (grant 30772565), National 973 project (grants 2007CB512006 and 2010CB12600), Shanghai-Unilever Research & Development Fund (08540750400), National Natural Science Funds for Distinguished Young Scholar (30888002), and Fudan postgraduate innovation fund (EYF129001).
Author Disclosure Statement
No competing financial interests exist.