
Abstract
Women live longer than men. Moreover, females live longer than males in some, but not all, experimental animals. The differences in longevity between genders are related to free radical production. Indeed, females produce less radicals only in animal species in which they live longer than males. This is because estrogens upregulate antioxidant longevity-related genes. These considerations have led us to postulate an extended concept of antioxidant in biology: an antioxidant is any nutritional, physiological, or pharmacological manipulation that increases the expression and activity of antioxidant genes or proteins. Phytoestrogens or other selective estrogen receptor modulators lower age-related diseases and prolong life span, at least in experimental animals. This provides rational bases to study their action in humans further. Antioxid. Redox Signal. 13, 269–278.
Different Longevity Between Women and Men
An important point is that the difference in longevity between women and men has occurred in all advanced as well as primitive societies. For instance, Table 1 indicates that difference in longevity in Spain in each year listed, from 1900 through 2000, always favored women (21). Moreover, the difference between women and men increased when average life expectancy approached the value of about 80 years observed today. In fact, in 1900 (i.e., when average life span was approximately 33 years), women lived only 3.8% longer than men. However, in 1992 when average life span was around 75–80 years, the increased longevity between women and men increased to almost 10%. This is by no means a characteristic of a given country (i.e., Spain), but rather it is a general phenomenon, and Figure 1 shows that there is a significant increase in life expectancy of females vs. males in Sweden (6). This has been calculated around 8%–10% over the last 250 years (i.e., from 1750 to present times). In all cases, women always outlived men by a significant proportion.
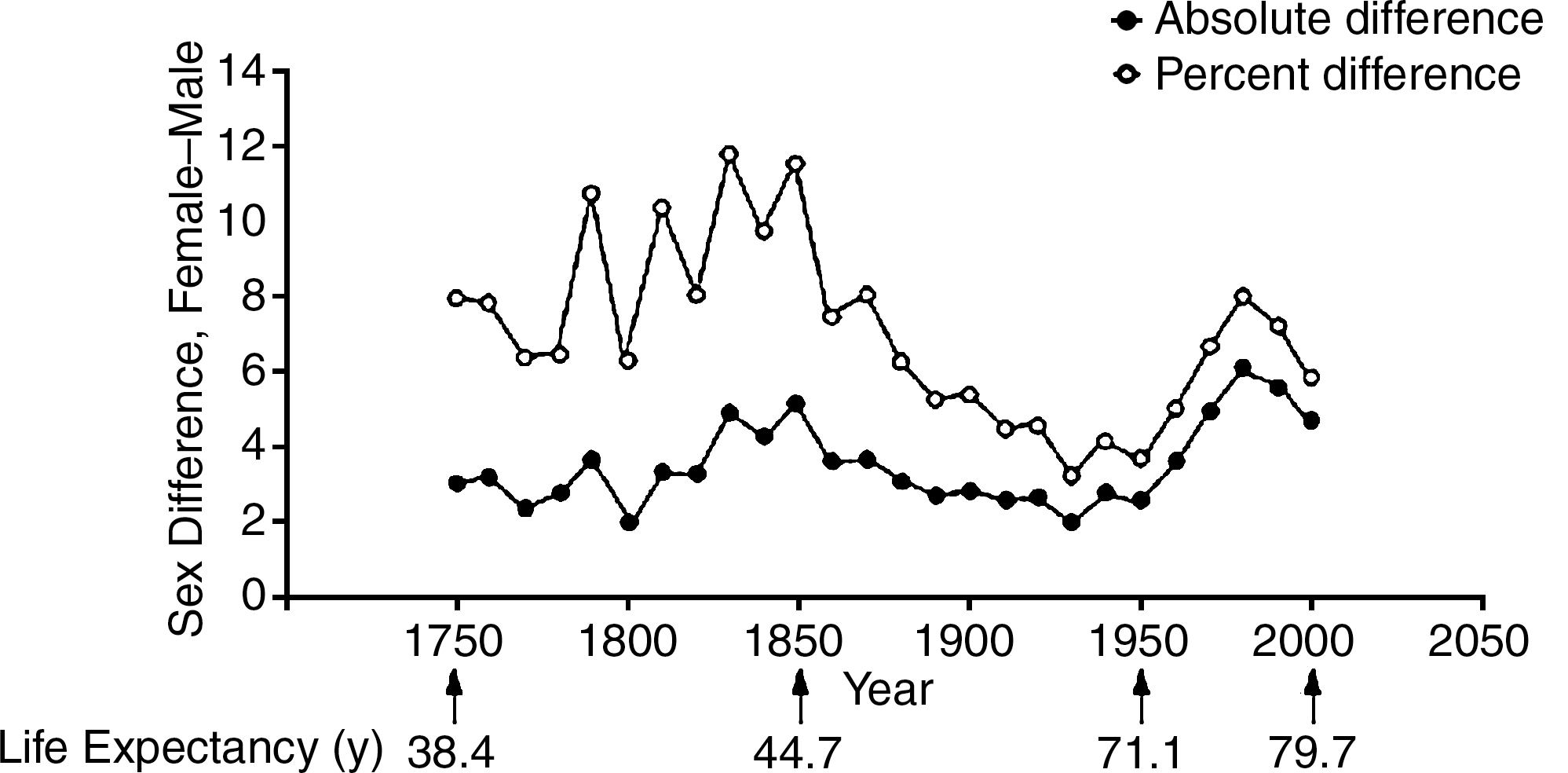
Data taken from Ref. (21).
Moreover, it is not only shown in the Caucasians from different regions, but also in other ethnics, women also live longer than men (4, 6, 12, 16, 17).
One could argue that this observed difference in longevity was due to sociological or environmental conditions rather than biological reasons. For instance, differences in the rate of smoking or in the type of work could account for some of the difference in longevity observed (19, 66). The following facts make this assumption unlikely: first, the differences are usually observed in all kinds of societies where social differences could be very marked. Second, this not only occurs in humans but also in animals, albeit not in all species. Therefore, it is reasonable to assume that there must be biological differences which explain the increased longevity of women as opposed to men.
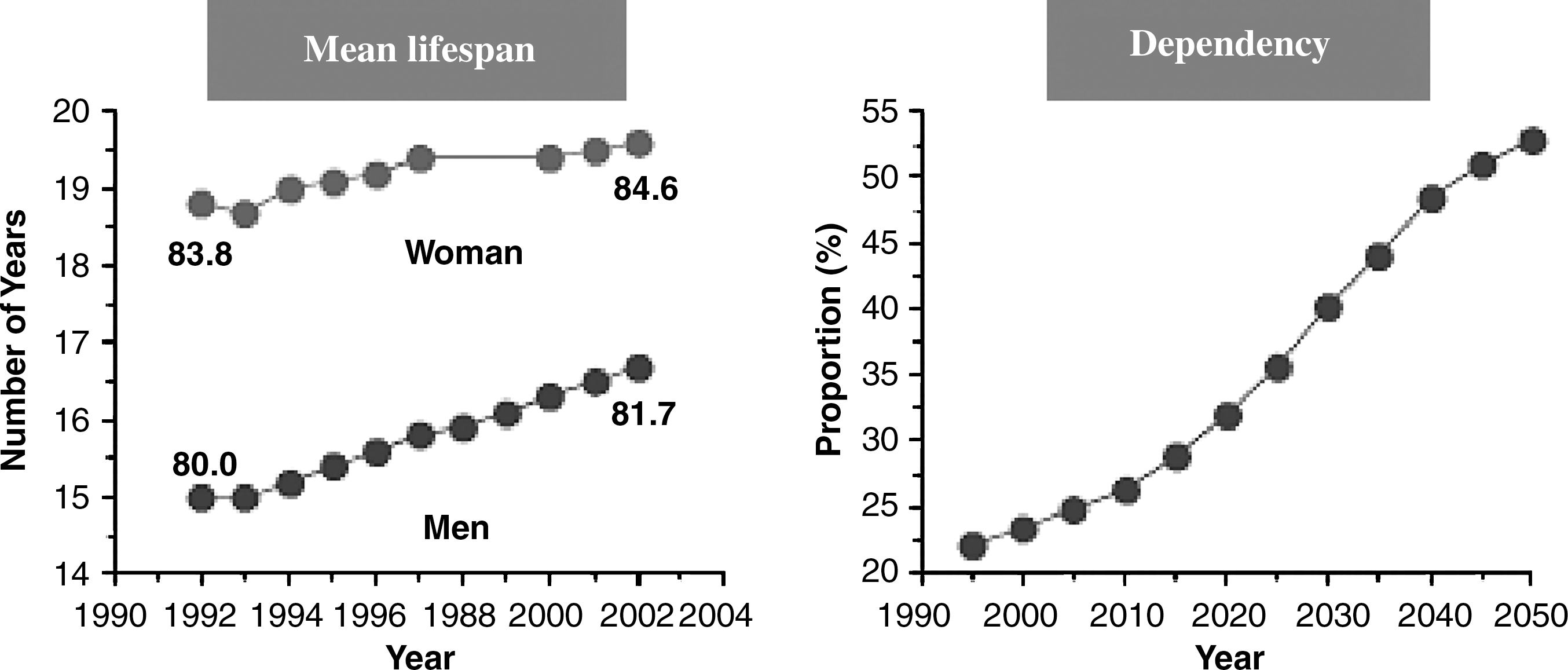
We would like to point out here another critical fact reported in Table 1: the increase in longevity in the 20th century has gone from approximately 34 years to approximately 80 years. Thus, this increase in longevity, independently of gender, is very remarkable. In fact, the average life span in Spain almost tripled in less than a century. This outstanding change had never occurred previously in recorded history and to our knowledge it is highly unlikely that it will happen again in the foreseeable future. And this leads us to consider another very important point, that of dependency. Since a vast portion of the population reaches ages well above 65 years, it is very important that we age successfully. In fact, the progress from independent healthy aging to frailty and eventually to dependency is a very important one that must be minimized by all means. Figure 2 shows that in the European Community the proportion of persons over 65 years who are dependent of care by others in 2010 is approximately 25%; however the predicted proportion of population of 65 years that is dependent on others will be about 50% in 2050. This means that of all the population of 65 years, half will be dependent on the other half for their lives. Any effort to understand aging with an aim to improving the quality of life of the aged population should be vigorously tackled. Lowering the proportion of frail people and eventually of dependent people must be a major task for gerontology and geriatrics (25)
An important concept that must be outlined here is that of the longevity curve. Figure 3 (panels A, B and C) shows that the shape of the longevity curves is very similar in different species, ranging from worms to humans. It is the X axis (i.e., the duration of the life span) that changes about a thousand fold from worms to humans. Critically important (see Panel D) is the fact that maximal life span is very difficult to change unless one acts fundamentally on the expression of longevity genes (see below). On the contrary, average life span may be modified by improving the life conditions of a society. Thus, in the days of the Roman Empire, the average life span was around 22 years; this was only very slightly improved in two millennia. Towards the end of the nineteenth century, the average life span in Western Europe was 40 years. As stated above, this more than doubled in the relatively short period of the twentieth century and the average life span went up to 85 years. The increase in average life span has been due to vast improvements in living conditions, particularly sanitation, preventive medicine, nutrition, and therapeutics. There are serious indications that this is the order of importance of the improvements in life span and that remarkable advances in clinical medicine such as the advent of antibiotics have had less significance to improve average life span than the very important improvements in public health such as sanitation, etc.

However, if we want to improve maximal life span then we must understand and intervene on the fundamental aging process. For this, it is critical to have good models of aging and prominent among them is studying why women live longer than men.
Differences in Longevity Between Males and Females
As stated above, differences in longevity between genders offer interesting possibilities to understand biological mechanisms of aging. Animals with a very similar genetic background may differ in their average life span for as much as 10%. Paramount among these differences is what happens in the humans species. However, in order to find proof that this is not due to sociological changes or peculiarities between societies (i.e., whether women smoke more or less than men), the differences in longevity in animal species must be understood.
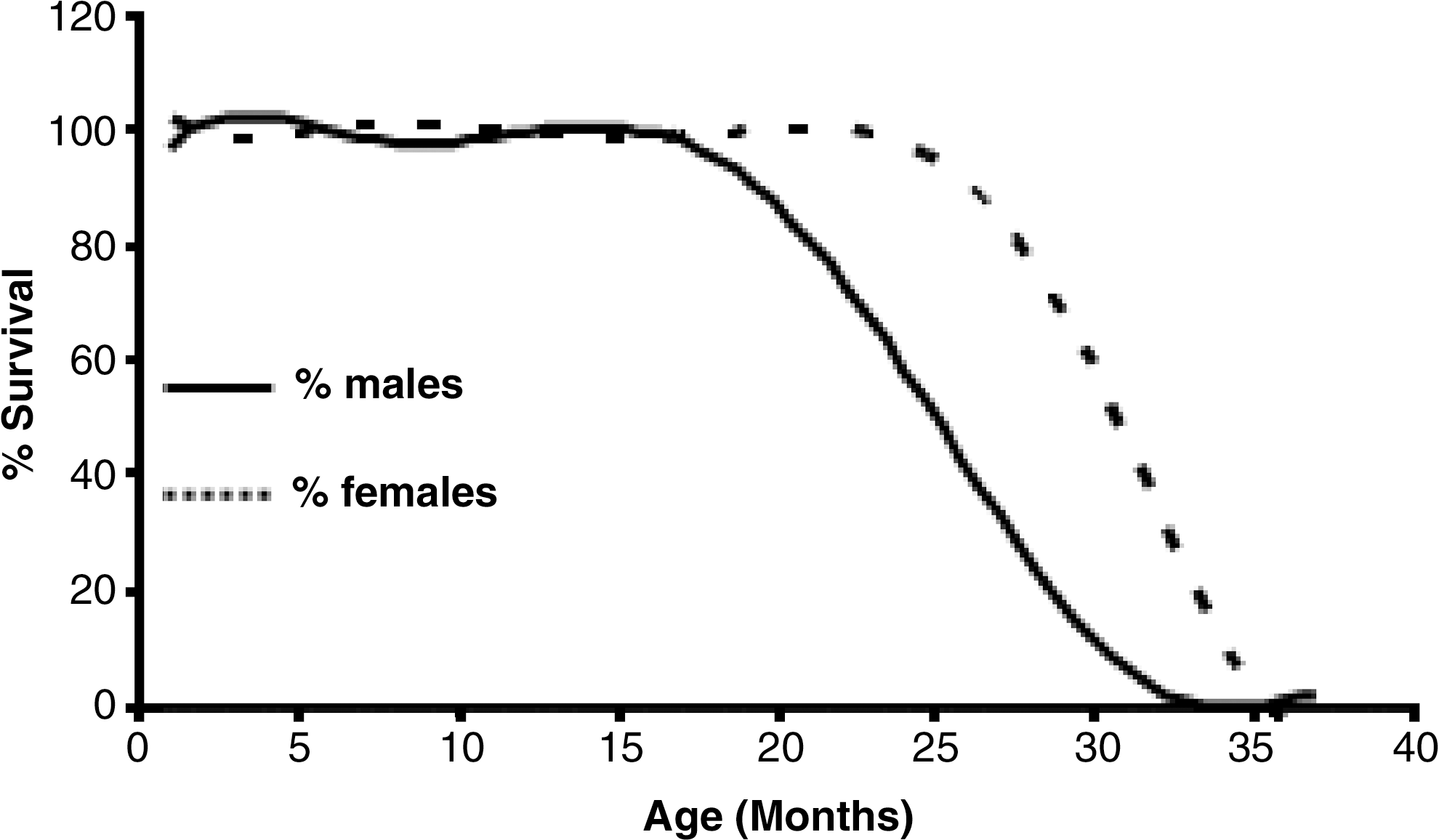
One of the most widely used laboratory animals is the Wistar rat. In this, we and others have observed that females live longer than males, the maximal difference in life span being approximately 10% (Fig. 4) (6, 10, 24, 44). We traced this difference in longevity to the presence of estrogens, as they protect females from oxidative stress. Moreover, it has been shown that ovariectomized females live less than control females, and that transplantation of young ovaries to old females induce an extension of longevity on mice (13, 46). In addition, previous work shows that estrogen replacement therapy can increase life expectancy in humans (20, 53).
Using Fisher 344 rats (the same species but a different strain in which longevity is also higher in females than in males), the group of Leeuwenburgh observed that males produce more reactive oxygen species than females (34). However, the higher longevity of females as compared with males is not a universal phenomenon. In other species of rodents such as mice, some strains show a higher longevity of males when compared with females, for instance the C57BL6 (2, 6). Moreover, a variation of this strain that was used by Leeuwenburgh's group (C57B16J mice) show no differences in longevity between sexes (57). In contrast, the Swiss albino mouse shows an increased longevity in females when compared with males (49). So not only do we have different gender specific longevity in different species, but also in different strains of the same species. This offers a unique opportunity to study comparative aging, that is, whether there are hormonal differences, different sensitivity or reactivity to hormones, or different fundamental molecular mechanisms of aging (2, 10, 57).
Differences in Longevity Between Genders Are Related to Their Mitochondrial Free Radical Production
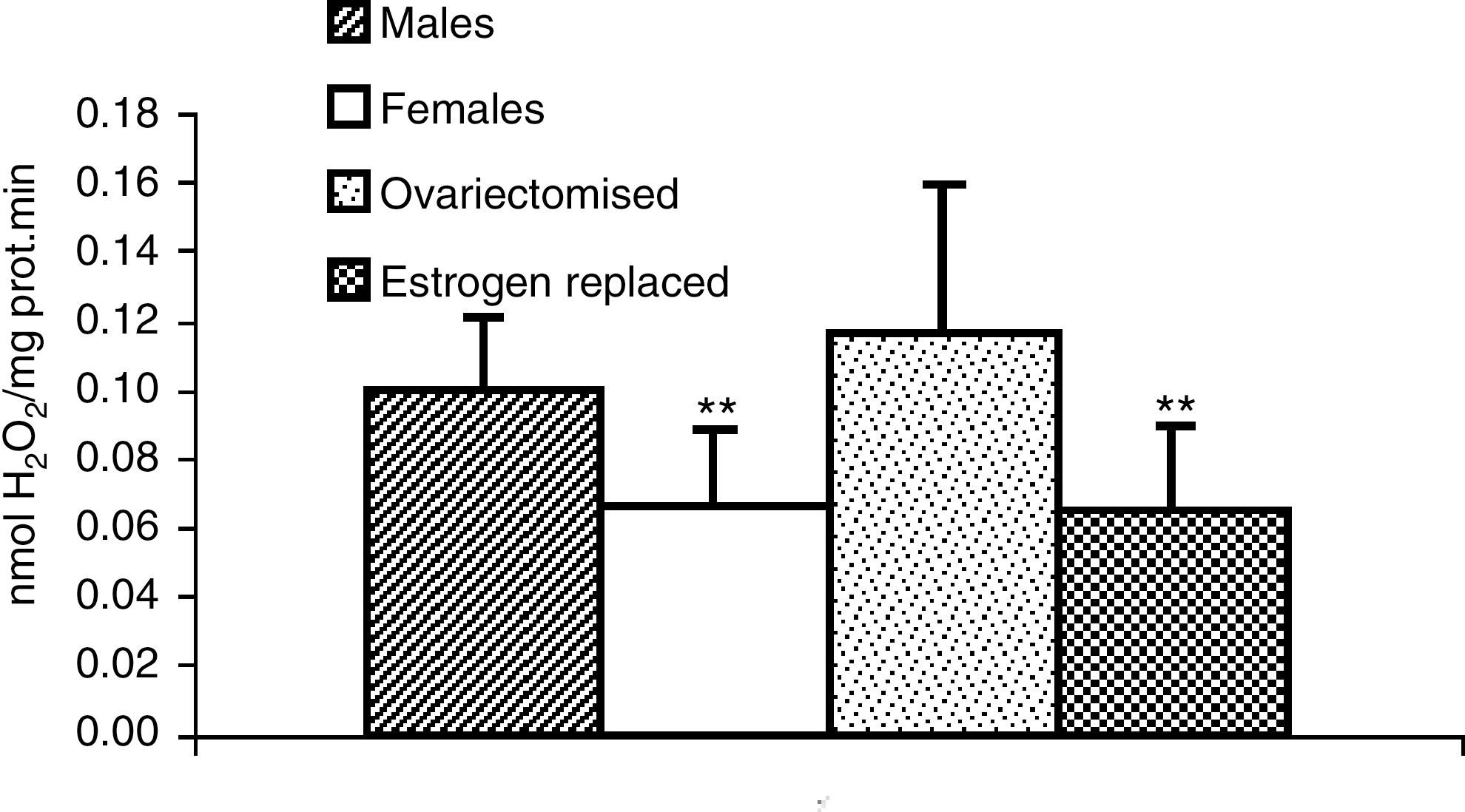
Studies in our laboratory using Wistar rats led us to observe that hepatic mitochondria from females produce approximately 50% the amount of peroxide produced by males (10). On the other hand, ovariectomized female Wistar rats produce approximately the same amount of peroxides as males (Fig. 5). When ovariectomized rats were treated with estrogens, they again produced the same amount of peroxide as normal female rats, that is, much less than males (Fig. 5). Thus, we were led to believe that the differences in radical production between males and females in Wistar rats were due to the presence of estrogens. Estrogens themselves are antioxidants because they contain a phenol structure in the A ring. However, plasma concentration of estrogens in a normal Wistar rat are so low (in a nm range) that it is highly unlikely that estrogens may behave as chemical antioxidants. Moreover, a simple calculation of the dose that is normally given to women after menopause (50 μg/day) when compared with a normal dose that is administered to persons who are treated with, for instance, vitamin E (approximately 400 IU/day) indicates that the dose of vitamin E is 8000 times more than that of estradiol. Therefore, it is highly unlikely that estradiol is thousands of times more antioxidant than vitamin E, considering estradiol as a free radical scavenger. Estrogens are antioxidants because their action as hormones, by interacting with estrogen receptors, unleash a cascade of events that lead to the antioxidant effects. We want to emphasize that we are not claiming that estrogens act in the same way in all species. The critical point is not to have higher or lower estrogen concentration in a given species or strain, but on the effect of estrogens on the cell signaling cascade that is going to be described later. Our conclusion, which will be described in more detail later in this review, is that estrogens in Wistar rats upregulate the expression of antioxidant genes, and these are the true antioxidants that prevent the release of oxidant species from mitochondria.
The very interesting study by Ali et al. showed that in those strains of mice such as the Black C57BL6 they were using in which males live longer than females, it is males that produce fewer oxidants than females (2). The authors measured not only oxidant production by determining dihydroethidium oxidation in brain but they also measured the electron paramagnetic resonance (EPR) spectroscopy signals in brain mitochondria of their mice. Thus, in this strain of mice in which males live longer than females, it is males that produce fewer radicals. Far from contradicting results from a laboratory, these results nicely confirm our results. The common claim by all of us is that the gender that lives longer produces fewer radicals independently of whether it is males or females who live longer. The critical test will be to see why estrogens promote the expression of antioxidant genes in a given species and do not promote that expression in other species or strains. A third confirmation of this hypothesis came from the laboratory of Christiaan Leeuwenburgh (57). These researchers studied a particular strain derived from the C57B6J in which males and females live the same length of time. Not surprisingly, these authors did not observe changes in either oxygen consumption, complex 1 and complex 3 oxidant production, protein carbonyls, oxidized DNA, or other indicators of oxidative stress. Thus, in those strains in which females live the same as males, there is no difference in oxidant production or in oxidative stress associated with gender. The overall conclusion is that estrogens promote a lower rate of production of radicals in those species or strains in which females live longer than males. On the other hand, in those animals in which males live longer than females, it is males that produce fewer radicals and those in which longevity is the same in both genders, radical production is also similar. This overview picture is summarized in Table 2.
Testing the Free Radical Theory of Aging: Mechanism by which Females Produce Less Oxidant than Males in Wistar Rats
From the above considerations, it is reasonable to think that in those species in which females live longer than males, they will produce fewer oxidants. This is indeed the case and, as stated above, we found that mitochondria from females produce approximately half the amount of oxidants when compared with males (10).
We have also stated that the concentration of estrogens made it highly unlikely that they could be considered as chemical antioxidants. Thus, we asked ourselves what the mechanism could be by which estrogens act as antioxidants and it turned out that estrogens have to bind to an estrogen receptor. As a consequence of this, a cascade of events is started by which activation of MAP kinases particularly ERK12 (p42–44) is activated. These kinases phosphorylate IκB, the inhibitor of NFκB, which eventually dissociates from the major complex NFκB/IκB, releasing NFκB and allowing it to enter the nucleus. Then it binds to NFκB responsive elements in the DNA and hence it activates the expression of mitochondrial antioxidant enzymes [i.e., SOD2 (formerly called manganese superoxide dismutase) and glutathione peroxidase] (8).
An important consideration concerning the integrity of mitochondria as the site of free radical production in cells must be made here. What we measure is the rate of oxidants that are released from mitochondria into the cytosol. This is the result of the difference between the production and the sites I and III of the respiratory chain and the LOCAL detoxification of the superoxide anions formed. Because the superoxide dismutase (SOD 2) and glutathione peroxidase are both in mitochondria, the oxidants that are released by these organelles are the results of the primary rate of production minus the rate of detoxification.
Our results indicate that it is mainly detoxification that is affected by estrogens, at least in Wistar rats, although we also have similar results in Oncins France 1 (OF1) mice. Thus, when we discuss that the rate of mitochondrial oxidant production, it is to be understood that the primary production and the respiratory chain minus the local detoxification by superoxide dismutase and glutathione peroxidase is what determines the “total rate of production by mitochondria”.
In Wistar rats or OF1 mice, this total rate of production is dependent on the action of estrogens because it is these hormones via their binding to estrogen receptors that activate the mitochondrial detoxifying mechanisms, leading to less oxidant production in females than in males. In a similar fashion, work from other laboratories, has found that when males live longer than females, it is males that produce fewer radicals.
A critical area of research for the future is to elucidate why estrogens upregulate detoxification of oxygen radicals at the mitochondrial level in some species and not in others. But, all in all, the fact that the rate of detoxification correlates so well with females living longer than males is, in our view, a strong confirmation of the validity of the free radical theory of aging.
The Extended Concept of Antioxidants in Biology
Antioxidants were described by Halliwell and Gutterige as “any substance that when present at low concentrations compared to those of an oxidizable substrate, significantly delays or prevents oxidation of that substrate” (30). This is of course an entirely chemical definition of the term “antioxidant”. Indeed, from a chemical viewpoint, an antioxidant must delay the oxidation of a substrate when present at low concentrations compared to that of the substrate.
However, we wish to propose an extended concept of antioxidant that applies more to complex biological systems such as organs or even organisms and is not restricted to the purely biochemical definition outlined above.
An antioxidant is any physiological, nutritional, or pharmacological manipulation that activates the expression of genes which codify for antioxidant enzymes, leading to a greater cellular resistance to oxidation by reactive oxygen or nitrogen species.
In this sense, we previously published (28) that physical training is an antioxidant. Of course, we do not mean that training is an antioxidant in the chemical sense but rather in this extended sense. Physical exercise activates the expression of a number of antioxidant enzymes such as superoxide dismutase, catalase, or glutathione peroxidase (27). This increased expression of activity of the antioxidant enzymes renders the cells more resistant to oxidation and therefore exercise may be considered as an antioxidant.
In a similar fashion, and it will be discussed later on in this review, treatment with estrogens or phytoestrogens which are known to upregulate the expression of antioxidant genes (8, 10, 43) may be considered as an antioxidant, because again treatment with these substances renders the cell more protected against the deleterious effects of oxidants.
This extended concept of antioxidants is indeed of relevance in the context of this review. We must emphasize that by no means do we claim that estrogens present in females protect them against oxidation via the conventional chemical antioxidant activity (i.e., because they have a phenolic ring). Indeed, estrogens are present in similar concentrations in rodents in which females live longer than males such as Wistar or Fisher 344 rats, as well as in those in which males live longer than females such as the Black C57BL6 mouse. It is not estrogens but rather the specific action of estrogens upregulating the expression of antioxidant enzymes and thus behaving as antioxidants in the extended concept rather than in the restrictive one that may explain the differences in longevity which are observed in several species like the Wistar rat.
Why estrogens (or phytoestrogens) act as antioxidants in the extended concept in some strains or species and not in others is an open question which indeed deserves further research to clarify.
Understanding of the extended role of estrogens as antioxidants in one or other species may shed light on fundamental aspects of aging and help us to understand not only the differential longevity between species or strains but more fundamental facts regarding the biology of aging.
The Concept of Longevity-Associated Genes
Longevity-associated genes were defined by Leonard Guarente and David Sinclair as genes whose modulation increases the life of a given organism (61). Therefore, the critical point is that we must identify genes whose modulation (up- or downregulation) results in an increased life span.
Of course, there is little interest in genes whose modulation results in a lower life span. It is important to know that upregulation of longevity genes result in an increased longevity, but it will be very interesting that the downregulation of a gene could also increase longevity; the important point being that modulation of the genes results in more and not less, longevity.
There are longevity genes of very many species and these are not limited to one type of reaction or one given chemical species. For instance, antioxidant genes are longevity genes and this was first pointed out by Raj Sohal and William Orr in Drosophila melanogaster almost 20 years ago. Indeed, overexpression of both superoxide dismutase and catalase results in an increased longevity of Drosophila melanogaster (51).
Another very interesting type of longevity genes are sirtuins. These are deacetylases that serve to silence big areas of DNA due to their interaction with histones (22). The substrate of sirtuins is not only histone molecules but many other proteins or peptides of biological interest. In any case, sirtuins are longevity genes because their overexpression results in an increased longevity of many species ranging from yeast to C. elegans and to vertebrates such as mice (29).
Recently, we have been able to identify that p53 and p16 are longevity genes because their overexpression in a regulated manner (the so-called super p53/p16 mice) results in an increased longevity (47). This increase in longevity is independent of the tumor-protecting activity of p53 because we observe increased longevity in the whole colony of mice. This increase was maintained when deaths by cancer of the mice were not taken into account. P53 is a longevity related gene because, in the absence of DNA damage, it upregulates the expression of a new class of powerful antioxidants called sestrins (11). These sestrins are peroxiredoxins which, as noted, are upregulated by p53. Thus p53 and p16 are longevity-related genes because their overexpression enhance longevity in mice (47).
We have also recently identified a new function for the well-known molecule telomerase. Telomerase itself is not a longevity gene because when upregulated it increases the incidence of cancer in the colonies of mice studies (5). If however, telomerase is overexpressed in animals that are already protected against cancer by overexpressing p53 and p16, then the triple transgenics show a remarkable increase in longevity (64). Indeed, these animals show a 50% increase in average life span which is the longest increase in life span observed by any discrete molecular manipulation of the genome. Why do triple transgenics of telomerase p16 and p53 show such a remarkable increase in longevity? Because they promote longevity by two different mechanisms: antioxidants and also by increasing the number of cell divisions because of the overexpression of telomerase.
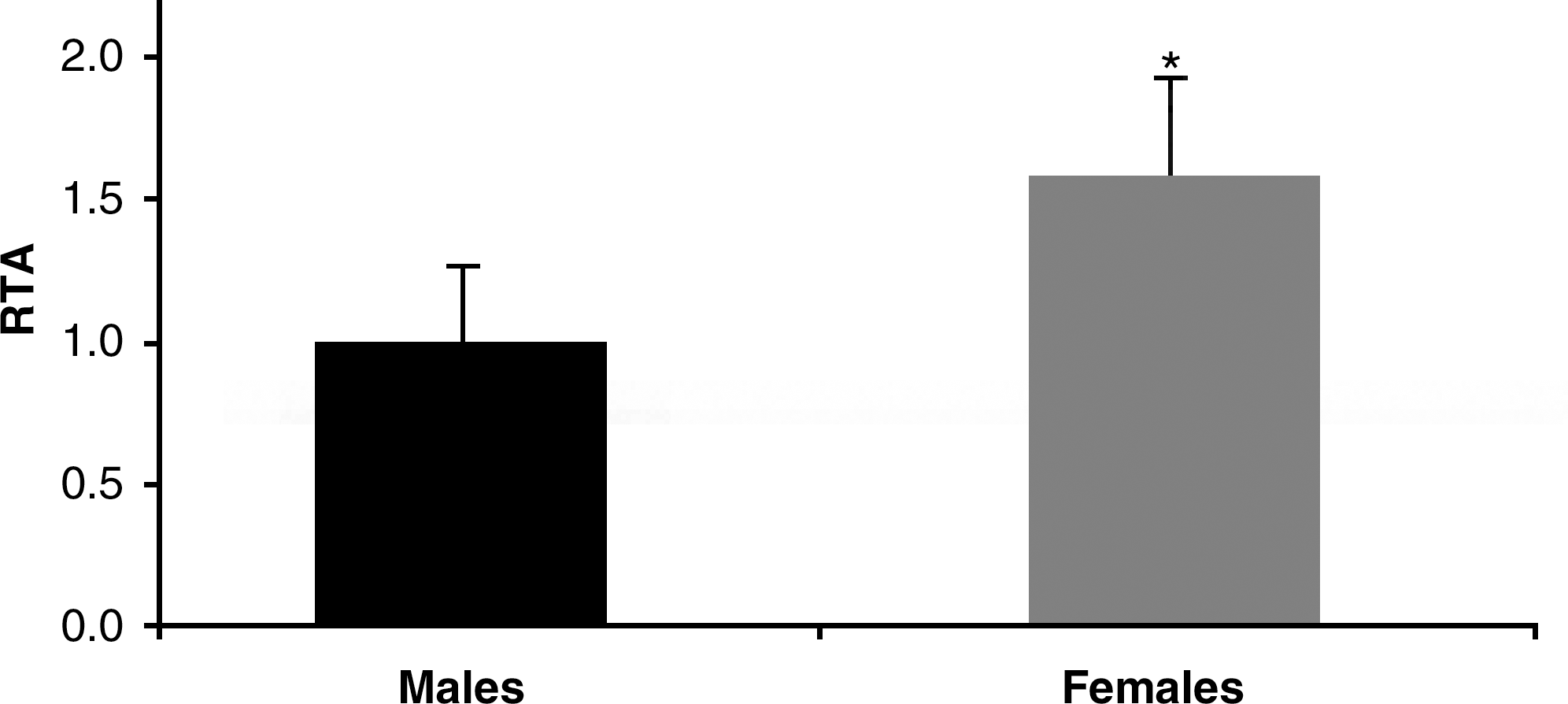
Telomerase contains an estrogen-responsive element in its promoter region (38). Thus, we reasoned that telomerase could be overexpressed in female rats when compared with males, and we found that its activity was higher in females than in males (Fig. 6). Moreover, we have found that telomerase is also regulated by glutathione (7). Since levels of glutathione in female Wistar rats are higher than in males rats, it is not unreasonable to think that telomerase may be upregulated not only in its expression because of the antioxidant-responsive element, but also in its activity because of the higher concentration of glutathione in cells from female rats when compared with males (10).
The concept of the longevity genes is thus very important because, if we identify these genes, then we may be able to find physiological or nutritional manipulation which may result in an activation of these genes and probably may help us to increase life span and more importantly, the quality of life in the old animals and persons. Indeed the overexpression of longevity genes not only increases life span, but also health fitness as shown, for example, in better motor coordination.
As stated above, females, in those species in which they live longer than males, do so because estrogens cause an upregulation of longevity-related genes. Of those we have identified, superoxide dismutase, glutathione peroxide, and telomerase, all are longevity-related genes that are upregulated in those strains and species in which females live longer than males (10).
Upregulating Longevity-Related Genes by Physiological, Nutritional, and Pharmacological Interventions: The Practical Problem
We have seen previously that in those species in which females live longer than males, estrogens are very useful because they upregulate the expression and activity of antioxidant, longevity-related genes (65).
This is certainly no surprise because estrogens act via estrogen receptors and promote the expression of life-prolonging genes. How this action takes place and why it does not in some species or strains in which males live longer than females is something that has not been experimentally tackled.
In any case, since in humans as well as in Wistar rats or Swiss OF1 mice, females live longer than males, it is reasonable to expect that estrogen therapy may be useful to upregulate the expression of longevity-related genes.
A major practical problem that presents itself when pondering the possibility of treating persons with estrogens is two-fold. On the one hand, it is obvious that it cannot be used in men because of the feminizing effects of estradiol (50). On the other hand, it may not be advisable to use it in women at perimenopause or in full-blown menopause because it may increase the risk of cancer (55), particularly in tissues which are sensitive to estrogens such as breast (32) or uterine cancer (1).
Epidemiological evidence indicates that treatment of women after menopause with estradiol may increase the risk of cancer (23, 39, 45), as well as that it of cardiovascular disease (37, 55). Moreover, there is large controversy about its beneficial effects at the neurological level (26, 31, 48, 56, 62, 68). Thus work on estrogen-like molecules as alternative treatment to classical estradiol should be pursued.
Two possibilities present themselves: one is pharmacological intervention (especially with synthetic estrogen receptor modulators) and the other is treatment with phytoestrogens (i.e., estrogenic molecules that come from plants and that have special effects on animals). Moreover, we can also aim at nutritional modifications, for example, increasing the amount of soya which is fed to the animals or persons. This offers the possibility of increasing the intake of phytoestrogens with foods that may be considered functional foods rather than with phytoestrogens or selective estrogen receptor modulators (SERMs) that have to be considered as drugs.
The suggested pharmacological and nutritional interventions with SERMs, phytoestrogens or changes in food are detailed in the two next sections of the present review.
Pharmacological Intervention: SERMs May Lower Age-Related Diseases
A selective estrogen receptor modulator (SERM) is a molecule that binds with high affinity of estrogen receptor but that has tissue-specific effects distinct from estrogens (40). SERMs have both agonist and antagonist estrogenic properties, depending on the target tissue. They are used in the treatment of menopausal symptoms as well as in treatments associated with cancer, especially breast cancer (33, 41, 58, 63). Their favorable vascular effects have been thoroughly studied. For instance, raloxifene lowers serum levels of cholesterol and homocysteine. It also improves endothelial function and reduces vascular muscle tone (40)
There is evidence of the neuroprotective effects of SERMs in the protection against neurodegenerative diseases (18). SERMs may act because they may have direct genomic, estrogen receptor-mediated effects, but also via a nongenomic pathway involving the activation of a number of cell signallers such as mitogen-activated protein kinases (MAP kinases). The possibility that they might act because of a direct, chemical, antioxidant action is remote (18).
The importance of the role of SERMs in cell signaling is illustrated by the fact that there is an acquired resistance to these compounds after chronic exposure. In fact, acquired resistance to tamoxifen is a serious therapeutic problem that can be explained by the cellular reaction to repeated stimuli with SERMs (35)
Phytoestrogens may be considered as SERMs because they bind to estrogen receptors and their action depends on the target tissue (3). But since they are components of foodstuffs, their action will be considered in more detail in the following section of this review.
The vast majority of studies regarding actions of SERMs are related to their action of treatment or prevention of specific diseases such as cardiovascular, breast cancer, or neurological conditions. Little, if any, efforts have been centered on the possibility that SERMs might increase longevity of male animals. There is certainly much information on the possible toxicity (15) of these compounds but not on the actual question of whether they might prolong life span.
Endocrine disruptors are another group of compounds that have weak estrogenic activity (42). Prominent among these is bisphenol A (36). They are most likely to cause harmful (rather than beneficial) effects (36). The issue is important. More that three billion kilograms of bisphenol A are produced per year and are used to manufacture plastics of all kinds. These are in close contact with foodstuffs and a proportion of these are ingested by the population. The very likely harmful effects of these endocrine disruptors on longevity are now being studied because their effects on longevity deserve close attention.
Nutritional Intervention: Phytoestrogens Upregulate Longevity-Related Genes
Considering the action of phytoestrogens to promote the expression of longevity-associated genes, one has to bear in mind the fact that there are two major types of estrogen receptors, one is estrogen-receptor alpha (ER alpha) and the other is the estrogen-receptor beta (ER beta) (52). The distribution of estrogen receptors in the organism is not homogeneous (60).
Previous studies in our laboratory showed that treatment of cells with genistein at concentrations normally found in the plasma of people who eat an Eastern-type diet (with a high intake of soya)(59) activates a pathway that is started by binding of genistein to estrogen-receptor beta [94% of the binding of genistein is to estrogen-receptor and only the rest, i.e., 6% to alpha (54)]. This eventually leads to the phosphorylation of the MAP kinases ERK 1 and 2, then to phosphorylation of the IKB molecules of the NFkB complex, liberating the p50 subunit of NFkB alpha. P50 enters the nucleus, binds to estrogen-responsive elements, and activates the expression of longevity-related genes, many of which are antioxidants (9). It is noteworthy that the concentrations of genistein that are effective in activating the pathway just mentioned are those found in plasma of people who have considerable amounts of soya in their diets (14). We are aware that only tiny fractions of the phytoestrogens that are ingested are absorbed (59). However, this would be a major setback if we were trying to explain antioxidant effects of the phytoestrogens based on their chemical structure and therefore so much dependent on concentration. But what we propose is that even minute concentrations (in the range of nanomolar) of this and probably other phytoestrogens may unleash an important response in the cell by binding those specific estrogen receptors and activating the expression of genes. It is in this kind of overall picture that one should see the role of phytoestrogens to promote beneficial effects through activation of genes.
Moreover, in collaboration with the team led by Professor Giovanni Mann at King's College in London, we tested whether modifications in food intake of soya might lead to effects similar to what we had observed in cells in vitro. And indeed it does: treating rats with a soya-based diet activates the expression of a number of longevity-related genes, as well as other interesting genes, such as eNOS, which are of physiological importance. Indeed, rats that took a soya-rich diet, showed lower blood pressure than controls (43). Thus, not only do we observe in vitro effects with phytoestrogens at concentrations present in plasma, but we do observe favorable effects of phytoestrogens when they are taken up with food. A methodological note of interest is that the normal laboratory chow which is fed to animals contains high amounts of soya (much more than our Western diet). Thus, the soya-containing diet is the normal laboratory diet and a soya-free diet which resembles a Western diet are to be specifically prepared for us to serve as control of the soya-containing diet. In this sense, ordinary food for animals in most laboratories is an “Eastern-type” food, based on soya, and one has to prepare food especially low in soya to resemble the normal Western diet (i.e., not based on soya).
In any case, the experiments just reported underpin the importance of diet modifications to supply phytoestrogens to animals and therefore activate the expression of longevity-related genes with the aim of increasing longevity of the male animals to resemble that observed in females. To our knowledge, no attempt has been made to compare a soya-rich and soya-poor diet on longevity of a controlled population of animals and this experiment is now being carried out in our laboratory.
Concluding Remarks
Females live longer than males in many species, including humans (21, 67). This also happens in the Wistar rat (10), but is not true for all the animal kingdom. The C57BL6 mouse does not show differences in longevity between genders and it may be that in some species males even live longer than females (2, 57).
The different longevity between genders provides a tool for research in gerontology because, with a very similar genetic background (same species, same strain), we find differences in longevity. For instance, Wistar rats show an increased production of radicals by mitochondria in males when compared with females. Concomitantly, females have higher glutathione levels and less mitochondrial DNA damage (65). In contrast, C57BL6 mice, in which females and males live the same length of time, do not show differences in mitochondrial radical production nor in oxidative damage to mitochondrial components (57).
We have traced the increased longevity of females to the protective effect of estrogens that do not act as chemical antioxidants but rather stimulate the expression of antioxidant enzymes. Estrogens bind to estrogen receptors, activate MAP kinases, and eventually activate phosphorylation of IκB (the regulatory subunit of NFκB). The NFκB enters the nucleus and activates expression of the antioxidant enzymes, such as glutathione peroxidase or Mn-superoxide dismutase (8). Of course, the difference in longevity cannot be due to estrogens per se (C57BL6 contain similar plasma concentrations as Wistar rats) but to specific actions of estrogens that occur in Wistar rats (and presumably humans) and do not occur in C57BL6 mice (2, 57).
On the practical side, since administration of estrogens to promote male longevity cannot be used because of their feminizing effects, we have used phytoestrogens, such as genistein. This flavonoid, which is present in relatively high levels in soya, at concentrations found in plasma of persons who eat an Eastern-type, soya-rich diet (59), promotes the expression of the same antioxidant, longevity- related genes as estradiol itself (9). Because genistein almost exclusively binds to estrogen receptor β, it does not have the feminizing effects of estradiol and may be used in males.
We propose that phytoestrogens and SERMs may be interesting compounds to try and increase longevity in male rats to values similar to females. Because humans share the same different gender difference in longevity as Wistar rats or Swiss mice, results of research to increase male longevity and fitness to resemble those of females are likely to have clinical importance.
Footnotes
Acknowledgments
This work was supported by grants BFU2007-65803/BFI from the Spanish Ministry of Education and Science (MEC); ISCIII2006-RED13-027 from the “Red Temática de investigación cooperativa en envejecimiento y fragilidad (RETICEF), and EU Funded COSTB35.
Author Disclosure Statement
The authors do not have any commercial associations that might create a conflict of interest in connection with this manuscript.