
Abstract
Hydrogen sulfide (H2S) is emerging as a physiological neuromodulator as well as a smooth muscle relaxant. We submit the first evidence that blood H2S levels are significantly lower in fasting blood obtained from type 2 diabetes patients compared with age-matched healthy subjects, and in streptozotocin-treated diabetic rats compared with control Sprague–Dawley rats. We further observed that supplementation with H2S or an endogenous precursor of H2S (
Introduction
Recent studies have revealed several potential roles for H2S in pathophysiologic functions (1, 13, 18, 20, 22, 29). H2S inhibits oxidative stress, promotes stimulation of KATP channels, relaxation and vasodilation in vascular smooth muscle cells, and relaxation of the human corpus cavernosum smooth muscle (1, 2, 18, 20, 22, 29). In vivo, H2S has been shown to inhibit leukocyte endothelial cell interactions and ischemia/reperfusion injury in liver and heart in animal studies (18). Recent studies have shown that genetic deletion of the CSE enzyme in mice markedly reduces H2S levels in the serum, heart, aorta, and other tissues, and that mice lacking CSE display pronounced hypertension and diminished endothelium-dependent vasorelaxation (29). These studies suggest that H2S is a physiological neuromodulator, vasodilator, and regulator of blood pressure (1, 2, 13, 18, 29).
This study examined the hypothesis that diabetes is associated with reduced blood levels of H2S. We determined blood levels of H2S in type 2 diabetes patients and in a rat model of diabetes. In addition, the effect of H2S and LC, an endogenous precursor of H2S, on HG-induced secretion of IL-8 and MCP-1 in cultured U937 monocytic cells was examined.
Results, Discussion, and Future Directions
The mean age of diabetes patients (52 ± 3 years) was similar to that of normal subjects (54 ± 7 years). The mean HbA1C of diabetes patients was 8.3 ± 0.5 (mean ± SEM). The mean IL-8 levels were significantly higher (p < 0.05) in diabetes patients (11 ± 1.2 pg/ml) compared with those in normal subjects (2.9 ± 0.6 pg/ml). MCP-1 levels were not significantly different between diabetes patients (491 ± 31 pg/ml) and normal subjects (432 ± 55 pg/ml) in our study population.
Diabetes patients have significantly lower blood levels of H2S compared with age-matched normal control subjects (Fig. 1a). In rat studies, a significantly lower plasma H2S level in diabetic rats was found compared with that in controls (Fig. 1b). At death, fasting blood HbA1 (16.98 ± 0.38% vs. 7.6 ± 0.2%) and MCP-1 (3,683 ± 423 vs. 2,164 ± 118 pg/ml) levels were significantly higher (p < 0.05) in diabetic rats compared with control nondiabetic rats. The effect of treatment with high glucose, without and with LC or H2S, on IL-8 and MCP-1 secretion in cultured monocytes is given in Fig. 2a and b. HG stimulated both MCP-1 and IL-8 secretion. Supplementation with exogenous H2S or LC, an endogenous precursor of H2S, inhibited the secretion of IL-8 (Fig. 2a) and MCP-1 (Fig. 2b) in HG-treated U937 monocytes. An osmolarity control for high glucose treatment (C2, consisting of 29 mM mannitol + 6 mM glucose) treatment did not show any effect on IL-8 or MCP-1 secretion (C2) in comparison with control (C1, 6 mM glucose).
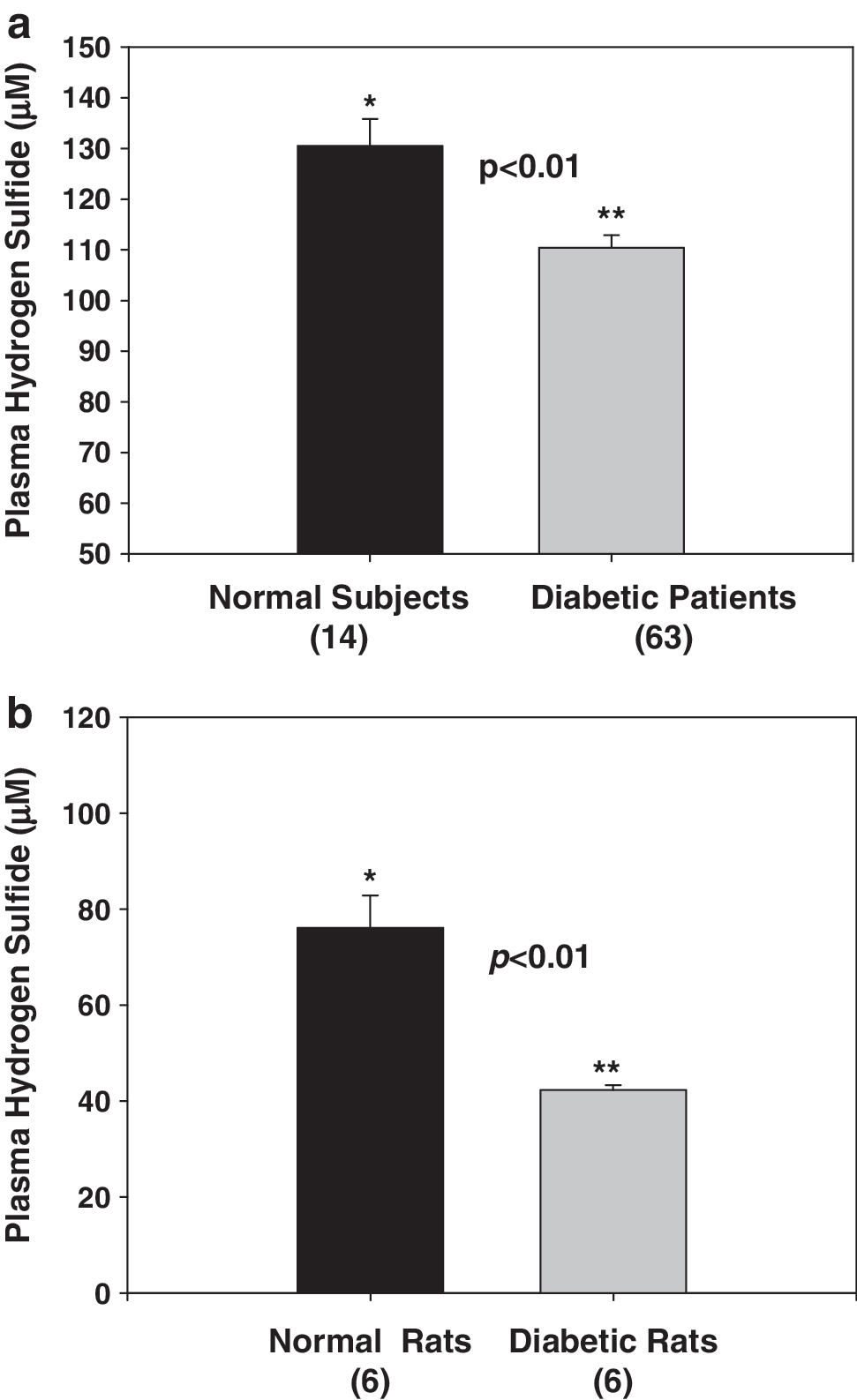

This study demonstrates that diabetes is associated with lower circulating levels of hydrogen sulfide. This was shown in both in type 2 diabetes patients and in rats made diabetic by using streptozotocin. The streptozotocin-treated rat is a model of type 1 diabetes, whereas the patients examined in this study were type 2 diabetes patients. Even though both types of diabetes are associated with hyperglycemia and elevated proinflammatory cytokines, further studies are needed to establish low hydrogen sulfide levels in blood from a type 2 diabetic rat model and in type 1 diabetes patients.
A recent excellent review by Olsen et al. (25) examined the blood/plasma hydrogen sulfide measurements that have been reported over the past 30 years from the perspectives of analytic methods used by various authors. Several investigators have reported blood levels of 30–100 μM hydrogen sulfide (25). These numbers have been questioned in this review (25) because no obvious odor of H2S gas in the blood or in exhaled alveolar air, even though micromolar concentrations of H2S should result in H2S concentrations in exhaled air that can be detected by the human nose.
Second, Furne et al. (10) predicted that the high level of sulfide production necessary to sustain micromolar sulfide concentrations would consume cysteine at a rate nearly 100 times greater than the daily requirement of cysteine and methionine combined. Furne et al. also showed that whole-tissue hydrogen sulfide concentrations are orders of magnitude lower than presently accepted values (10). The method used by Zhu et al (30) measured not only the basal H2S but also H2S released from acid-labile sulfur and from bound sulfane sulfur (15). Hydrogen sulfide is gaining acceptance as a signaling molecule and has been shown to elicit a variety of biologic effects; however, the precise concentrations of H2S are present in tissues and are required for biologic effects is a matter of considerable debate.
A monocyte cell-culture model has been used to explore this understanding of the various mechanisms that contribute to the development of vascular inflammation and atherosclerosis (9, 24). The monocyte infiltration into subendothelial space that occurs through the combined actions of locally produced chemotactic cytokines and adhesion molecules expressed on the injured endothelial surface is critical to the development of vascular inflammation and the development of atherosclerosis. Previous studies have shown that activation with LPS is needed to elicit proinflammatory cytokine secretion in response to high-glucose treatment of monocytes (11, 16).
We focused our study on MCP-1 and IL-8 because these cytokines do not need activation by LPS to elicit a secretion response to HG treatment in monocytes. In this study, cells were exposed to a high-glucose concentration of 35 mM. Many previous studies have reported that glucose concentrations as high as 50 mM have been found in the blood of patients with uncontrolled diabetes (5). It is true that blood glucose levels in patients are not likely to stay as high as 35 mM for 24 h. However, tissue damage in diabetes patients occurs over many years of countless hyperglycemic episodes. Thus, the glucose concentration of 35 mM used in this cell-culture study does not seem unreasonable.
Many diabetes patients have elevated blood levels of the proinflammatory cytokines, such as IL-8 and MCP-1 (9, 12). These proinflammatory cytokines are markers of vascular inflammation, a known risk factor for cardiovascular disease (9, 12). Darmaun et al. (7), by using an isotope method, provided evidence that an alteration in glutathione and cysteine homeostasis occurs in vivo in adolescents with poorly controlled type 1 diabetes (6, 7). The glutathione and cysteine depletion are likely to arise from increased glutathione use. Along with a host of proteins,
In conclusion, this study demonstrates for the first time that diabetes is associated with lower circulating levels of hydrogen sulfide, which may play a role in vascular inflammation. H2S or
Materials and Methods
Diabetes patients and normal volunteers
The protocol followed the guidelines approved by the Institutional Review Board for the Protection of Human Research Subjects. Blood from overnight-fasted patients was collected into tubes with EDTA. The EDTA blood was centrifuged, and the clear plasma saved for H2S, MCP-1, and IL-8 assays. All subjects were type 2 diabetes patients (n = 63) and age-matched normal volunteers (n = 14).
Animal studies
All of the procedures were carried out in accordance with the ethical standards of the institution after approval by the institutional Animal Welfare Committee. Male Sprague–Dawley rats were purchased at 49–52 days of age (200–220 g) from Harlan (Indianapolis, IN) and allowed 2 days for environmental and trainer-handling acclimation. The rats were weighed and then fasted overnight before intraperitoneal injection of 65 mg/kg streptozotocin in citrate buffer (pH, 4.5). Control rats were injected with citrate buffer alone to serve as a normal control group (group 1). The rats were tested for hyperglycemia by measuring their blood glucose concentration at 3 and 7 days after the streptozotocin injections. Blood for the blood glucose was obtained via tail incision and measured by using an Advantage Accu-chek glucometer (Boehringer Mannheim Corp., Indianapolis, IN). The rats that became hyperglycemic (blood glucose >300 mg/dl) were randomly divided into two groups: group 2, diabetic rats. The rats were maintained under standard housing conditions at 22 ± 2°C with 12:12-h light/dark cycles with a standard 8640 lab chow diet (Harlan, Indianapolis, IN). At the end of 7 weeks, the rats were fasted overnight and then killed for analysis by exposure to halothane (2-bromo-2-chloro-1,1,1-trifluoroethane). At the time of death, seven rats were in the control, and six in the diabetic group. Blood was collected via heart puncture with a 19½-gauge needle into EDTA Vacutainer tubes. EDTA blood was centrifuged; the clear plasma was saved, and buffy coat layers were discarded.
Human monocytes and treatment with high glucose (HG), LC, or H2S
U937 monocytes (two million cells/ml) were suspended in RPMI media containing 6 mM glucose. This medium is prepared by using glucose-free medium (Sigma Catalog number R1383) and was then mixed with glucose and serum and antibiotics. These cells were treated with LC or H2S with high-glucose medium (+29 mM glucose giving a final concentration of 35 mM) without and with LC or H2S. The concentrations of LC and H2S are given in the figures (10, 100, and 1,000 μM); treatments were carried out at 37°C for 24 h. Na2S was used as a source of H2S (19). Cells were pretreated with LC or hydrogen sulfide for 60 min before the treatment with high-glucose medium. Mannitol was used as an osmolarity control. All experiments were repeated 4 times.
Plasma H2S was determined by the method of Zhu et al. (30). Interleukin (IL)-8 and monocyte chemoattractant protein-1 (MCP-1) levels in the supernatant of treated cells and in the plasma of human and MCP-1 in the plasma of rats were determined by using ELISA kits from Fisher Thermo Scientific Co. (Rockford, IL) and from R & D Systems (Minneapolis, MN). We did not assay levels of IL-8 in the blood of diabetic rats because a rat-specific kit for IL-8 is not commercially available. All appropriate controls and standards as specified by the manufacturers were used; the data are expressed as cytokine secreted per two million cells or per milliliter of plasma. In the cytokine assay, control plasma samples were analyzed each time to check the variation from plate to plate on different days of analyses. The assays were repeated if the variation in control plasma value from day to day was >7%. Information on age and HbA1c was collected from patients' medical records in the clinic. Glycosylated hemoglobin in rats was determined by using Glyco-Tek Affinity column kits and reagents (catalog number 5351) purchased from Helena Laboratories (Beaumont, TX). This Glycated-hemoglobin method assesses HbA1. All chemicals were purchased from Sigma Chemicals (St. Louis, MO) unless otherwise mentioned. Data in figures is given as mean ± SEM. Data between different groups were analyzed statistically by using ANOVA on Ranks with Sigma Plot and Sigma Stat statistical software (SPSS, Chicago, IL). For cell-culture studies, Student's t test was used for data analyses. A value of p < 0.05 was considered significant.
Footnotes
Acknowledgments
The authors are supported by grants from NIDDK and the Office of Dietary Supplements of the National Institutes of Health (RO1 DK064797 and RO1 DK072433). The authors thank Ms. Georgia Morgan for excellent editing of this article. None of the authors has any financial interest in the publication of this article.