
Abstract
Vegetarian diets are rich in antioxidant phytochemicals. However, they may not act as antioxidants in vivo, and yet still have important signaling and regulatory functions. Some may act as pro-oxidants, modulating cellular redox tone and oxidizing redox sensitive sites. In this review, evidence for health benefits of vegetarian diets is presented from different perspectives: epidemiological, biomarker, evolutionary, and public health, as well as antioxidant. From the perspective of molecular connections between diet and health, evidence of a role for plasma ascorbic acid as a biomarker for future disease risk is presented. Basic concepts of redox-based cell signaling are presented, and effects of antioxidant phytochemicals on signaling, especially via redox tone, sulfur switches and the Antioxidant Response Element (ARE), are explored. Sufficient scientific evidence exists for public health policy to promote a plant-rich diet for health promotion. This does not need to wait for science to provide all the answers as to why and how. However, action and interplay of dietary antioxidants in the nonequilibrium systems that control redox balance, cell signaling, and cell function provide rich ground for research to advance understanding of orthomolecular nutrition and provide science-based evidence to advance public health in our aging population. Antioxid. Redox Signal. 13, 1575–1591.
Introduction
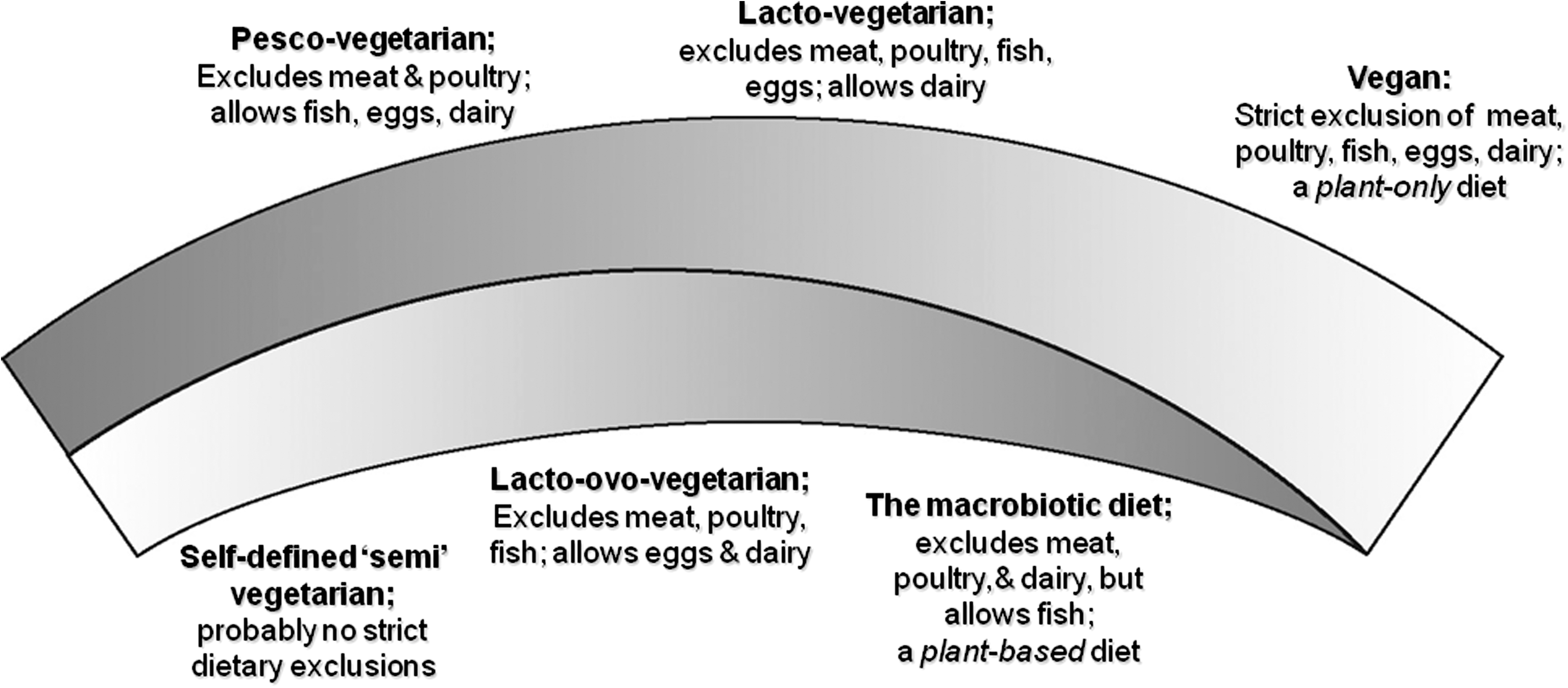
Health Benefits of Vegetarian Diet: Epidemiological, Biomarker, Public Health, Evolutionary and Antioxidant Perspectives, and a Possible Novel Role for Vitamin C
Epidemiological and biomarker perspectives
Epidemiological and biomarker findings in relation to vegetarian diets and the major public health challenges, diabetes, cardiovascular disease (CVD), and cancer, are discussed in this section.
Diabetes
Type 2 diabetes mellitus (T2DM) accounts for >90% of the 150 million cases of diabetes worldwide (2, 152). T2DM associates with central obesity, decreased high density lipoprotein cholesterol (HDL-C), increased plasma triglycerides (Tg), total cholesterol (TC), low density lipoprotein cholesterol (LDL-C), uric acid, and high sensitivity C-reactive protein (hsCRP) (2, 27, 142, 175). In combination, this increases CVD risk in T2DM 2–4 fold (65). Vegetarian diets are generally low in fat, calories, simple sugars, and cholesterol, and this helps prevent T2DM (87). The relationship between diet and incident diabetes over 17 years' follow-up was recorded in 8401 subjects nondiabetic at baseline (167). Nonvegetarians were 29% more likely to develop diabetes, and a diet that included at least weekly meat intake was associated with 74% increase in risk in comparison to a meat-free diet. After BMI adjustment, Odds Ratio (OR) for incident diabetes in omnivores was 1.34; 95% CIs 1.03, 1.75 (p < 0.05) (167). In the Adventist Health Study 2 (159), incident diabetes was examined in 60,903 subjects (159); vegan and lacto-ovo vegetarian diets associated with ∼50% lower rates compared with nonvegetarian diets (OR 0.54; 95% CIs 0.49, 0.60; p < 0.05). T2DM is characterized by hyperglycemia and insulin resistance. In 95 healthy lacto-ovo vegetarians (mean duration of diet: 10.2 years), insulin resistance and fasting plasma glucose were lower (p < 0.05) than in 107 omnivores. Mean (SD) glucose in the vegetarians was 4.47 (0.05) mmol/L vs. 4.71 (0.07) mmol/L in the omnivores; insulin was 4.96 (0.23) vs. 7.32 (0.41) mU/L; insulin resistance (HOMA-IR) was 0.99 (0.05) vs. 1.59 (0.10) (164). Higher insulin sensitivity was found also in lacto-vegetarian premenopausal women compared to omnivores (n = 49/group) (80).
Cardiovascular disease
Cardiovascular disease (CVD) causes 30% of deaths worldwide (174, 175). Various risk factors are known, but there are several emerging factors, such as high uric acid and homocysteine, low antioxidant status, and oxidative stress (9, 39, 135, 175). The vegetarian diet is likely to meet dietary guidelines for CVD risk reduction (2, 18). In the Health Food Shoppers Study and the Oxford Vegetarian Study, which each included ∼11,000 subjects, average mortality from heart disease was ∼15% lower in vegetarians, though this did not reach statistical significance: death rate ratios (DRRs) were 0.85 (95% CIs 0.71, 1.01) and 0.86 (95% CIs 0.67, 1.12), respectively (96 –98). Analysis of five prospective studies (Adventist Mortality, Health Food Shoppers, Adventist Health, Heidenberg, and Oxford Vegetarian Diets) involving totally 27,808 vegetarians in an overall study cohort of 76,172 men and women (mean follow-up 10.6 years) showed that CVD mortality was 24% lower in vegetarians than nonvegetarians (DRR 0.76, 95% CIs 0.62, 0.94; p < 0.05) (99). This is in accordance with the finding of 0.4 mmol/L lower cholesterol levels found in vegetarians, which is expected to decrease CVD mortality by ∼20% (96, 97). Less clear findings came from a German study (38) in which vegetarians had nonsignificantly lower mortality (DRR 0.71, 95% CIs 0.41, 1.18), but a significant trend (p = 0.006) of increasing heart disease risk with higher meat consumption was seen. The Epic-Oxford study (98) of 47,254 subjects with no history of CVD at recruitment showed DRR of 0.81, 95% CIs 0.57, 1.16 for IHD, (nonsignificant) in vegetarians, but their mean serum LDL-C was ∼12% lower than in meat eaters.
Vegetarians have lower blood pressure, BMI, and lipids in comparison with nonvegetarians (7, 41, 58, 90, 108, 151, 154, 156). The effect on lipids is more pronounced in a strict vegetarian raw food diet. In study of 201 adherents to a mainly raw food diet, plasma TC, HDL-C, and LDL-C decreased with increasing proportion of raw food consumed (103). The atherogenicity of oxidized LDL is greater than native LDL, and LDL of vegetarians was reported to be more resistant to oxidation that nonvegetarians (112). Other biomarkers found to be lower in vegetarians include uric acid (58, 69, 154, 156), and urinary sodium and potassium (156). Uric acid is an independent risk factor for CHD, although the mechanism is unclear (16). Low urinary sodium reflects low intake, which improves blood pressure. Plasma hsCRP is also lower (p < 0.05) in vegetarians (41, 154). This highly sensitive marker of inflammation is increasingly used in CVD risk assessment. Glycemic control relates to CVD risk, and fasting glucose is reported to be lower in vegetarians (58), though not all studies find this (41). In relation to hemostasis, overall there is a favorable impact, as shown in terms of lower concentrations of coagulation factors or increased fibrinolysis (54, 131).
Epidemiological studies rarely produce clear-cut evidence of association between a specific factor and disease unless the impact of a single agent is very large. A complementary approach to the study of plant-based diets and health is to look at the amount of plant-foods taken and measure biomarkers of intake, and to compare these in people who subsequently stay healthy and those who do not. Fruit and vegetable consumption was found to inversely associate with CVD risk in a meta-analysis of nine studies incorporating 91,379 men and 129,701 women (46). Risk of CVD decreased by 4% and 7% for each additional daily portion or fruit and vegetable, respectively (46). In a study of dietary intakes in 350 cases of myocardial infarction (MI) and 700 matched controls, relative risk (RR) of MI was lower in those who took >3 servings per day of vegetables (132). Lower CVD mortality over an average of 9-year follow-up was seen with higher of intake of fruits, vegetables, and legumes in 10,499 participants with self-reported diabetes (126).
A vegetarian diet has various cardioprotective elements; however, a note of caution is needed. Avoiding all foods of animal origin may counterbalance some health advantages, as plant-only diets lack vitamin D, vitamin B12, heme iron, and zinc (18, 133). As an example of how this impacts health, vitamin B12 deficiency increases plasma homocysteine (Hcy), increasing CVD risk (18, 37, 118).
Cancer
Cancer, the second leading cause of death worldwide, has its origin in mutations in key genes controlling cell growth and proliferation and DNA repair (43, 81, 173). Diet is known to be an important modulator of risk, and many case control and epidemiological studies show that high intake of fruits and vegetables is protective (5, 21, 38, 49, 95, 98, 137, 144, 158, 173). In the EPIC-Oxford study, which included 52,706 participants, the risk of cancer incidence was compared between meat eaters, fish eaters, and vegetarians (98). Risk of malignant neoplasm was lower among fish eaters (IRR 0.83, 95% CIs 0.71, 0.96; p < 0.05), and borderline significantly lower in vegetarians (IRR 0.89, 95% CIs 0.80, 1.00) compared to meat eaters. Further investigation into incidence of cancers at 20 different sites in vegetarians was reported (98). Data were pooled from two prospective studies (the Oxford Vegetarian and EPIC-Oxford cohort) totaling 33,697 subjects. Relative risk (RR) of cancer was significantly lower in vegetarians (RR 0.88, 95% CIs 0.81, 0.96; p < 0.05) and fish eaters (RR 0.82, 95% CIs 0.73, 0.93; p < 0.05). Stomach and bladder cancers were significantly lower among vegetarians than nonvegetarians, but results were more variable for cancers of other sites. In collaborative analysis of five prospective studies (the Adventist Mortality, Health Food Shoppers, Adventist Health, Heideberg, and Oxford Vegetarian), DRR from different type of cancers showed no large differences but somewhat lower mortality in vegetarians (99).
Cancer is a group of complex and different diseases, and diet is equally complex and difficult to study. Nonetheless, there are elements in a vegetarian diet that can be regarded as cancer-preventive, including high content of selenium, folic acid, chlorophyll, fiber, and antioxidants (13, 29, 49, 52, 68, 173), Conversely, high intake of red meat, total calories, and salt increase risk of some cancers (49, 173). Vegetarian diets are often high in soya foods that are rich in phytoestrogens, and this is thought to lower breast cancer risk. Isoflavones intake and breast cancer risk in a cohort of 37,643 British women was investigated (163). Vegetarian women had lower average incidence of cancer (RR, 0.91; 95% CIs 0.72, 1.14), but the effect was not significant, and no evidence for a strong association between vegetarian diets in the pre- or postmenopausal period or dietary isoflavone intake and risk of breast cancer was seen (163).
Other than some heritable mutations, there are no validated biomarkers of cancer risk, but markers of genomic stability and DNA methylation, damage, and repair are potential candidates (43, 61, 62, 70) In a study of 13 lacto-ovo-vegetarians (average duration of diet: 10.8 years), 11 lacto-vegetarians (average duration of diet: 8.2 years), and 24 healthy omnivorous controls, chromosome aberrations, micronuclei and DNA damage (as strand breaks measured using the comet assay) were examined in peripheral blood lymphocytes (92). Vegetarians showed slightly lower levels of oxidative DNA damage in lymphocytes, but genomic stability was unaffected (91, 92). The effect of a vegetarian diet was investigated also in regard to DNA methylation, which is involved in gene regulation. No difference was found in 48 lacto-ovo vegetarians compared to 23 vegans in one study (62), but buccal cells from 40 young vegetarians and 40 age-matched omnivores showed decreased promoter methylation and a 3-fold increase in expression of superoxide dismutase (SOD) (157).
Summary, and the public health perspective
The balance of evidence is on the benefit side for vegetarian diets. Evidence is strongest for diabetes. Effects on CVD and cancer are complicated by potential nutritional inadequacies of an entirely plant-based diet and the lack of reliable biomarkers of cancer risk. It must be noted that the strong associations between vegetarian diet and improved biomarkers profiles and even health outcomes may be due to other facets of a healthy lifestyle. Nonetheless, public health recommendations should not wait upon results of randomized placebo-controlled intervention trials, confirmatory evidence for mechanistic action, or identification of the exact bioactive(s) in plant food (4). Noncommunicable disorders account for ∼50% of the world's global burden of disease (174), and the associated demands of our aging populations are overwhelming healthcare systems, even in affluent, well-developed countries. This highlights the need for a healthcare paradigm that puts emphasis on proactive, preventive strategies. Study is still needed to determine key agents and confirm effects at cellular and molecular levels. However, connections between plant-based diets and health have an evolutionary as well as an antioxidant perspective. Furthermore, findings in large-scale epidemiological studies (38, 78, 101) suggest a novel role for plasma ascorbic acid in connection to health assessment. This is discussed briefly, followed by evolutionary and antioxidant perspectives.
A potentially useful connection: Plasma ascorbic acid and disease risk
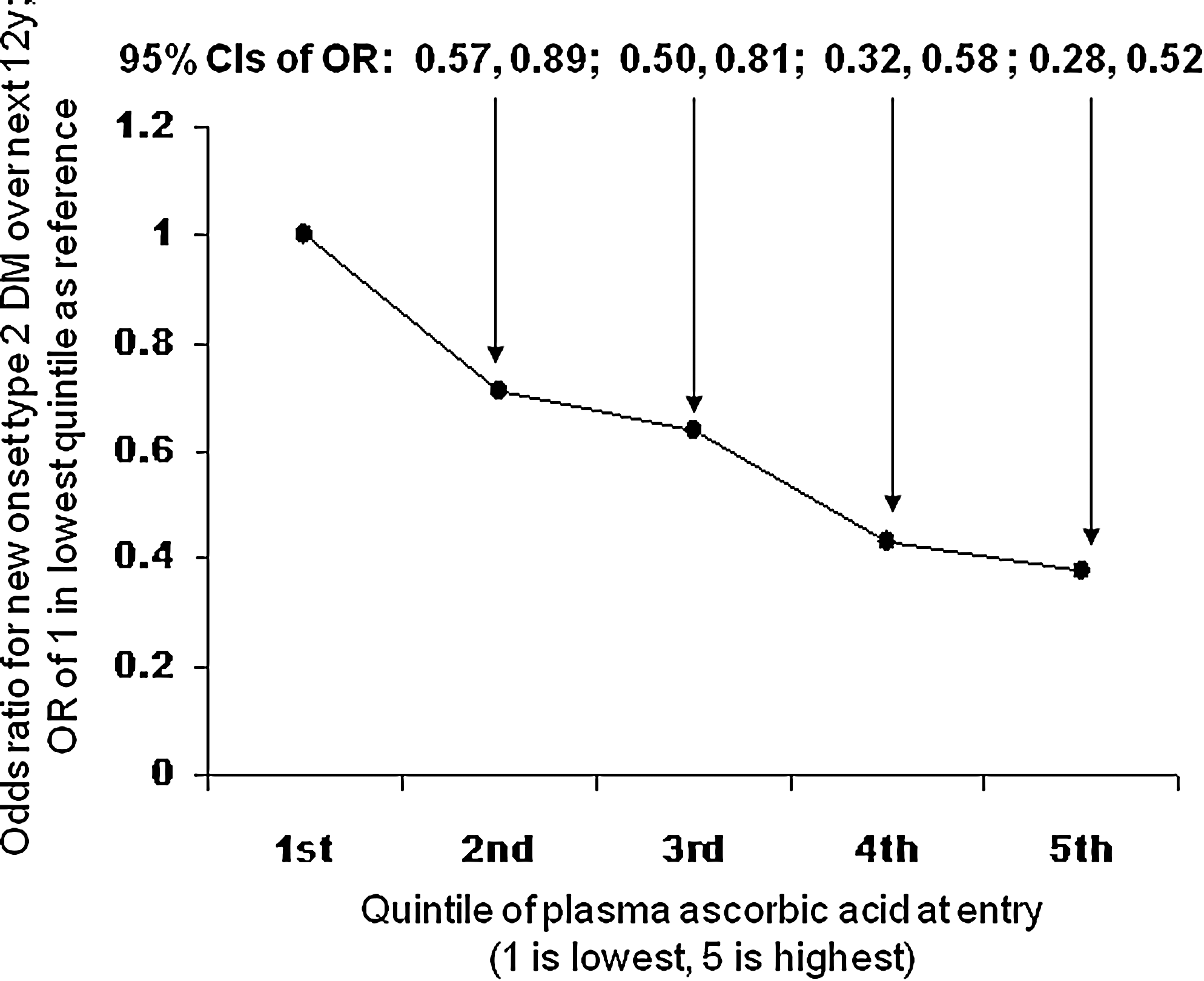
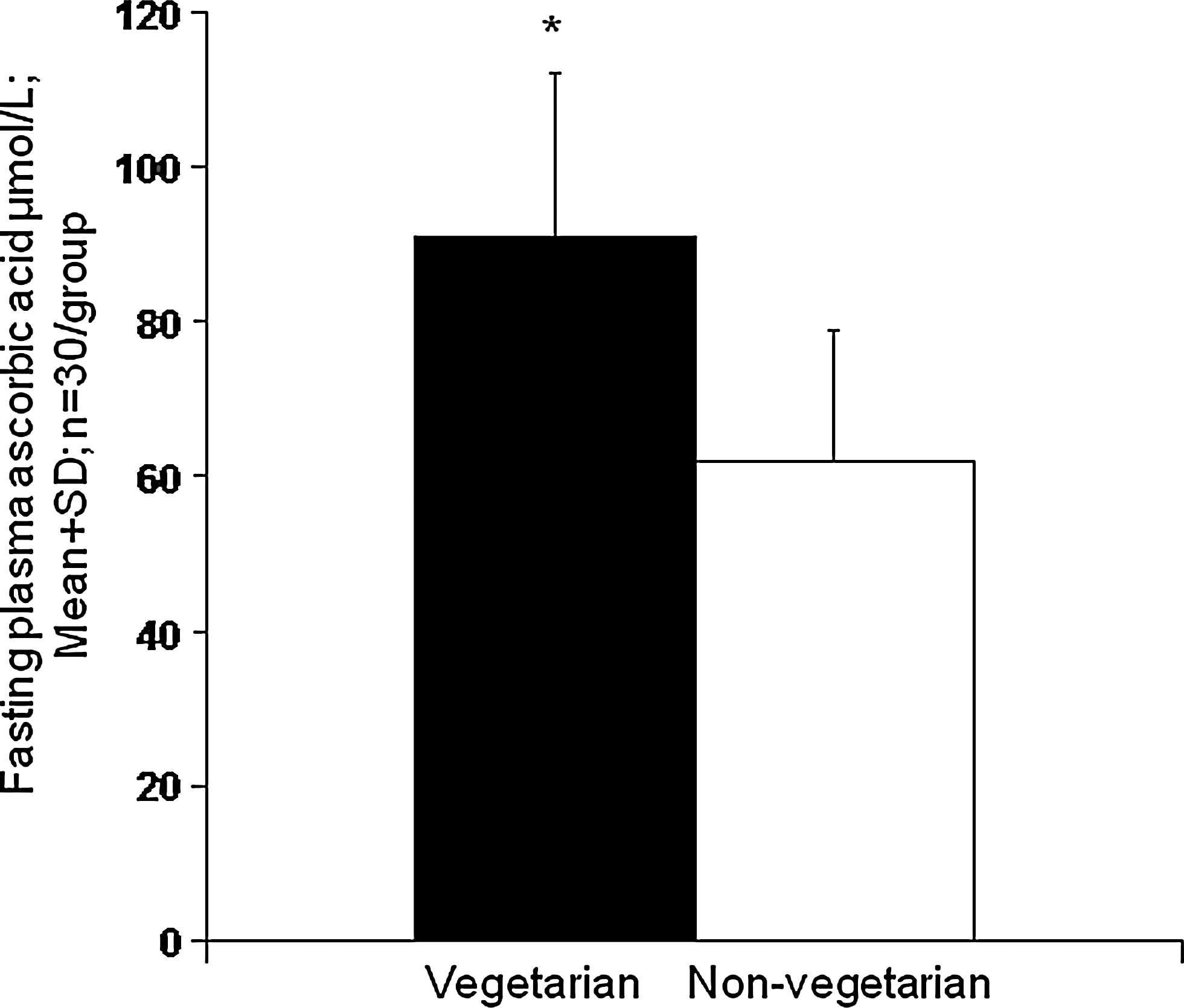
In the EPIC Norfolk study, 19,246 subjects free of diabetes at entry were followed for up to 12 years (78). Incident diabetes was higher (p < 0.5) in those with lower ascorbic acid at entry (Fig. 2). The relationship remained marked and showed a continuous linear trend (p < 0.05) after adjusting for smoking, hsCRP, waist–hip ratio, and other factors, with ∼29% decrease in risk per 20 μmol/L change in plasma ascorbic acid. Dietary input is the only source of ascorbic acid in the human body, it is recognized as vital for optimal health, and vegetarians have high levels (Fig. 3) (48, 57, 64, 154).
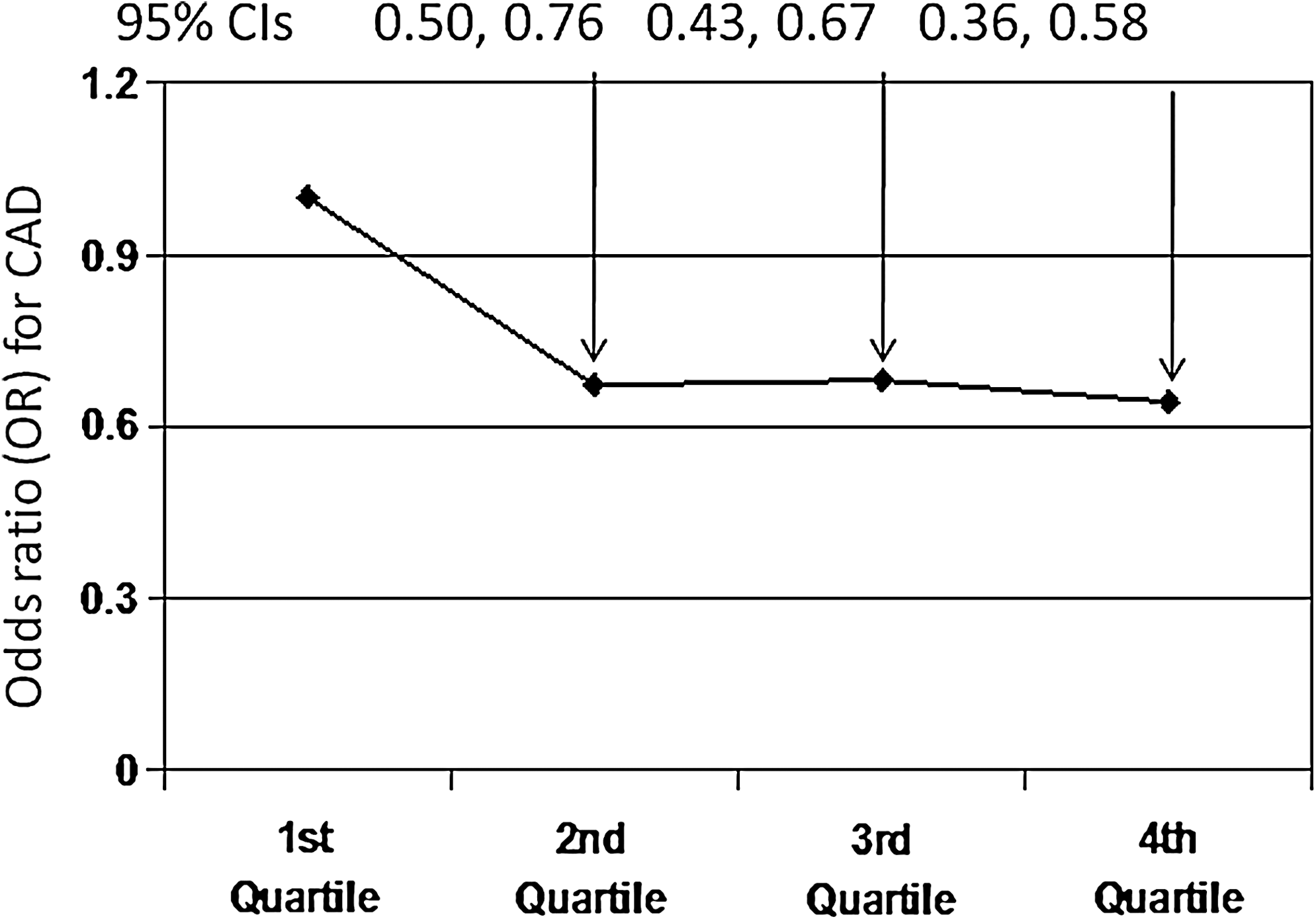
Strong inverse relationship between plasma ascorbic acid and subsequent coronary artery disease (CAD), stroke, and overall mortality was also seen (30, 101, 122). Risk of incident CAD was ∼40% lower (p < 0.05) in those in the highest quartile of ascorbic acid concentration compared to the lowest quartile (Fig. 4). Fasting plasma ascorbic acid of >55 μmol/L was proposed to confer minimal CVD risk (64). This level is likely to be attained by vegetarians, but many apparently healthy omnivores have much lower levels, and as noted, they have higher rates of subsequent ill-health and overall mortality. However, supplementation with vitamin C is not beneficial (unless there is pre-existing deficiency) (5, 28, 79), indicating that other components in plant foods are needed. Indeed, high plasma ascorbic acid may be a coincidental accompaniment to (and simple marker of ) high intake of these other components.
Fresh fruits and vegetables are the main sources of vitamin C (26, 29, 155), but lower disease risk was not seen with dietary input of fruits and vegetables (30, 78). This may be related to the limited absorption of the vitamin (12, 48), or to increased usage in inflammation or subclinical disease. This is supported by the finding of an inverse relationship between ascorbic acid and hsCRP (30,154), though relationship shown in Figure 4 was independent of hsCRP (30). Whether association is driven by vitamin C, another dietary component, or to orchestrated action within an antioxidant network remains to be confirmed (56, 105, 110, 144). Nonetheless, the strong association suggests that plasma ascorbic acid may be a useful marker of overall health status (42).
Evolutionary perspective of plant-based diets and health: An ancestral connection?
Aerobic respiration is a fundamental feature of most life on Earth. However, partial reduction of oxygen forms the more reactive species, superoxide, and from this various other reactive oxygen (and nitrogen) species (ROS/RNS) can be formed (70, 72). Superoxide and nitric oxide are not highly reactive, but their interaction produces the very reactive peroxynitrite, and other highly damaging ROS can be formed directly or indirectly from superoxide (11, 70, 128). Biological damage caused by ROS (‘oxidative stress') favored development of antioxidant defenses, and the human body has evolved to have a multilayered and integrated antioxidant system (22) (Fig. 5). This system is effective, but not entirely so, and oxidative changes to DNA, lipid, and protein are detectable in human cells and fluids (43, 44, 61, 70, 120). What accounts for this evolutionary ‘flaw’? One explanation relates to evolutionary theory and metabolic cost-benefit ratio: evolutionary ‘pressure’ is removed at a metabolic break-even point. Another explanation is that damage due to inadequate antioxidant defense was biologically less important, at least in the short term, than a system that removed ROS too effectively and prevented their use as, for example, signaling agents.

Differences in antioxidant content between modern and ancient diets are relevant in regard to the metabolic break-even point explanation. Crop growing began ∼12,000 years ago and brought dramatic and rapid change to the mainly vegetarian hunter–gatherer diet (22, 23, 31). The antioxidant content of the modern-day omnivorous diet is estimated as markedly lower than that of our Paleolithic ancestors (23, 31) (Fig. 6).
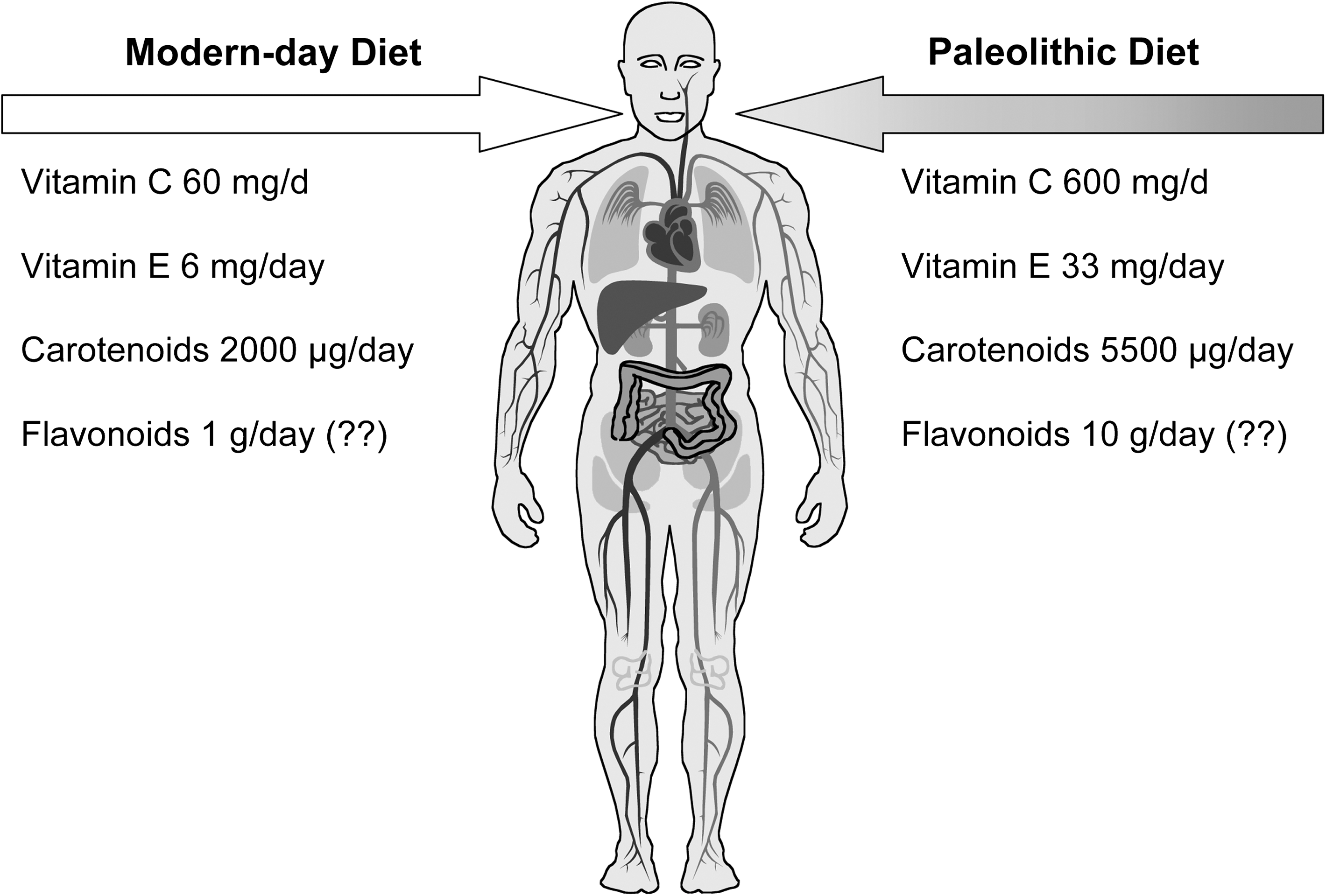
Obligate high content of antioxidants in the ancestral diet could have made up for a shortfall in endogenous antioxidants, relieving pressure for further innovation in the evolving hominid system. There is no direct evidence for this theory, but humans retain the gene (L-gulono-1,4, lactone oxidase) that catalyzes the final step in synthesis of ascorbic acid, though it is highly mutated and nontranscribed (40). Our ancestors lost the ability to synthesize ascorbic acid, and yet retained an absolute metabolic requirement for it. This innovation would have been sustained only if it brought biological advantage, such as a saving in biosynthetic costs, but this could only have been within a scenario where synthesis was not needed because of the diet (22, 23). If so, then high intake of ascorbic acid (and possibly other plant-derived antioxidants) is what suits our still largely ancestral physiology. Another explanation is that biological damage due to inadequate antioxidant defense was less important than a wholly effective system that prevented the use of ROS as bioactive agents or messengers. The two suggested scenarios are not mutually exclusive, though in one there is no disadvantage to ‘excess' antioxidant input, whereas in the other there is an important concept of redox (im)balance. With this in mind, the antioxidant perspective of a vegetarian diet, including content and effects on antioxidant status and oxidative stress, is now presented, and Part 2 explores the role of ROS in signaling and the possible effects of dietary antioxidants on redox balance and signaling pathways.
The antioxidant perspective of a vegetarian diet
There are many diverse antioxidants in plant foods, and it is not possible to identify or measure each individual one. One approach has been to measure ‘total antioxidant content’, another to use biomarkers of antioxidant status and oxidative stress markers in relation to diet.
The ‘total antioxidant content’ of various types of foods and effects on plasma antioxidant status have been published (e.g. 10, 14, 15, 17, 20, 25, 60, 74, 75, 111, 127, 143, 155, 166, 171, 172). Halvorsen et al. presented the total antioxidant (redox active) content as the Ferric Reducing/Antioxidant Power (FRAP) value of over 1000 commonly consumed foods, as well that of many herbs and spices (74, 75). Other groups have contributed to the now substantial literature on antioxidant content of foods, which can help guide food production, preparation, and selection for higher antioxidant intake. But what difference does high intake make to body systems? There is limited bioavailability of phytochemicals, which could be a physiological safeguard against possible toxic effects of what are essentially xenobiotics (71). This may help avoid antioxidant overload and a tipping of redox balance to the reductive side. There is also the issue of biotransformation and antioxidant efficiency: biotransformation might decrease or increase antioxidant activity, and in vitro ‘total antioxidant content’ of a food is not a measure of its antioxidant efficiency in vivo. Still, there is evidence that at least some antioxidants from foods are absorbed, and that dietary changes affect antioxidant status and oxidative stress. It is worth noting that post-ingestion changes in total antioxidant content of plasma (often used as a measure of antioxidant absorption) can be confounded by effects on uric acid. Red wine and fructose-rich foods increase plasma uric acid, leading to changes in plasma total antioxidant content, while increased ascorbic acid in plasma lowers uric acid by increasing renal excretion (14, 16, 48, 111, 127). To correct for changes in uric acid, it is advisable to remove it (with uricase) before measuring plasma antioxidant content or, in the case of FRAP assay measurements, simply subtracting 2x plasma uric acid concentration from the FRAP value (the stoichiometric factor of uric acid is 2.0 in the FRAP assay and does not vary with concentration) (14, 15).
As noted, ‘total antioxidant content’ is not easily related to antioxidant action/efficiency within the body, which is the extent to which an agent protects biomolecules against oxidation-induced damage. Dietary antioxidants may have different efficiencies and react differently with different oxidizing species. Therefore, diet-related effects on individual antioxidants and biomarkers reflecting antioxidant action and oxidative stress within the body are more relevant. Haldar et al. showed that plasma carotenoids and ascorbic acid are higher in healthy nonsmoking vegetarians than in omnivores (69). In a Slovakian study, plasma ascorbic acid and β-carotene concentrations were higher (p < 0.01) in old (aged 60–70 years; n = 33) vegetarians compared to 34 age-matched nonvegetarians (104). These increased (p < 0.01) in 32 men supplemented with a fruit and vegetable concentrate (140). In study of 30 long-term (5–55 years duration) Chinese vegetarians and 30 age- and sex-matched Chinese nonvegetarians, vegetarians had an average fasting plasma ascorbic acid concentration nearly twice that of the nonvegetarians (154) (Fig. 3), although lipid standardized α-tocopherol concentration was ∼11% (p < 0.01) lower, perhaps due to lack of nuts or seeds and to high phytic acid intake in the diet of the Taoists studied (154). The plasma total antioxidant capacity was not significantly different between vegetarians and nonvegetarians, but the contribution of ascorbic acid to this was markedly higher (and that of uric acid lower) than in the nonvegetarians. hsCRP was lower (p < 0.01) in the vegetarian group, and significant (p < 0.05) inverse correlations were seen overall between ascorbic acid and hsCRP, and between ascorbic acid and uric acid (154).
Measuring oxidative stress in vivo is no simple task: most biomarkers used are nonspecific, prone to artifact, or have unknown relevance to disease. Plasma malondialdehyde (MDA) is a commonly used but nonspecific biomarker of lipid peroxidation often measured by the artifact-prone TBARS (thiobarbituric acid reacting substances) method, and measurement of conjugated dienes of LDL oxidation is performed under extremely unphysiological conditions and has unknown relevance to health (24, 70). Oxidation-induced damage to DNA can be measured reliably using an enzyme-assisted version of the comet assay (43), though obtaining human nucleated cells other than white blood cells is very difficult, and the relevance of ‘global’ DNA damage to disease risk is not yet clear. More specific and sensitive tests of oxidative stress are F2 isoprostanes (70, 81, 120), and 8-oxodG (8-oxo-7,8-dihydro-2'-deoxyguanosine), which in urine represents ‘whole body’ oxidative stress (44, 61). Plasma allantoin is (in humans) the nonenzymatic oxidation product of uric acid (19), but has not been widely used in oxidative stress assessment.
In biomarker studies, lower (p < 0.05) plasma malondialdehyde (MDA) in both male and female vegetarians was found compared to nonvegetarians, and the lag time of in vitro conjugated diene formation was longer and TBARS formation was lower in LDL from vegetarians (112, 114). A study of young (20–30 years) and old (60–70 years) ovo-lacto-vegetarian and nonvegetarian women reported no differences in oxidative stress biomarkers (DNA strand breaks measured in an enzyme-assisted comet assay, protein carbonyls, and fatty acid conjugated dienes) between the young vegetarians and nonvegetarians, but older vegetarians (n = 33) had lower (p < 0.05) oxidative stress than the age-matched nonvegetarians (n = 34) (104). Interestingly, old vegetarian women did not show the higher oxidative stress levels seen in old nonvegetarians, implying that vegetarian diets prevent age-related increases in oxidative stress.
In summary, there is some biomarker evidence of effects of vegetarian diet on antioxidant status and oxidative stress, but measurement of effects on oxidative stress is far from trivial, and the reliability and clinical relevance of biomarkers of oxidative stress is controversial and unclear. Effects of vegetarian diets undoubtedly extend beyond simple antioxidant action, but new and sensitive biomarkers reflecting orthomolecular action of dietary components are needed. An emerging and rich area of study is in the role of redox active molecules in control of redox tone, cell signaling, and metabolism, and how phytochemicals contribute to or impact on this. This is explored below.
Redox Balance, Tone, Switches and Signaling; Connections to ROS and Dietary Antioxidants
Concepts of redox signaling, tone and balance
Oxidation and redox balance are basic themes in aerobic life (22, 72, 130, 165). Redox balance is a nonequilibrium thermodynamic state determined by relative and constantly changing amounts of oxidative and antioxidative (reductive) agents (94). Redox balance in turn determines the state of oxidation-sensitive sites (43, 70, 134, 165). Oxidation of some sites is damage, but changes in redox ‘tone’ of others act as biological switches for molecular function, cell signaling, adaptation to change, and cytoprotection (1, 34, 35, 55, 63, 72, 77, 86, 94, 107, 128, 150). Superoxide, hydrogen peroxide, nitric oxide, and peroxynitrite are important players in cell signaling, and oxidized fatty acids and other lipids have also been shown to have important signaling functions (107, 150, 180). Importantly, in the context of this review, phytochemicals are increasingly recognized as regulators of cell signaling (32, 63, 169, 179). Here, concepts of redox signaling and signal transduction by ROS are briefly discussed, with a view to exploring how the antioxidant-rich vegetarian diet could affect redox tone, switching, and signaling. For more detail on redox-based signaling and its regulation, readers are referred to several excellent reviews (34, 86, 94, 150).
Key sites of direct redox action are protein thiol groups of sulfur-containing amino acid residues, mainly cysteine (77, 86). Redox sensitive thiol groups, ‘sulfur switches,’ can exist in the reduced thiol (SH) form, the oxidized sulfenic acid form, or as a disulfide bridge (-S–S-) between two oxidized thiol groups within a protein (77, 88). Disulfide bridging can reveal or mask a functional site (e.g., recognition, catalytic, or binding site) within a protein. These sites are referred to as thiol/sulfide redox couples or ‘redox control nodes' (77, 94). Redox partners are required for interconversion. An electron acceptor partner (i.e., an oxidizing species) helps create the bridge; an electron donor (a reducing species) is needed to restore the SH groups and so break the bridge. Important endogenous electron donors include glutathione (GSH) and the thioredoxin (Trx), peroxiredoxin, and glutaredoxin families (8, 89, 109, 123). These employ their own thiol groups as the source of electrons, and also require redox ‘recycling’. GSH is regenerated from its oxidized GSSG form by glutathione reductase (GR), and the family of thioredoxin reductases (TrxR) restores reduced Trx (8, 88, 89, 94, 109). These enzymes need electron-donating co-substrates, and these are sourced from ‘front line’ endogenous electron donors, NADH or NADPH. Therefore, there is a chain of redox reactions involving sulfur switches that connects basic metabolic products to the control of redox control nodes, and through this to biochemical and cellular events (34, 86).
Control of each sulfur switch in the chain is determined by redox tone or balance of the microenvironment, by the redox potential of the couple, and by the presence and action of the disulfide reductases and electron donors (reductants; electron donating antioxidants). These are determined by metabolic fluxes, biochemical components, and interactions in the cell at large, but are controlled directly and dynamically by the immediate environment.
There are multiple redox control nodes, with individual and varying redox potentials (94). Such a system is highly challenging to investigate, but offers a sensitive, elegant, and dynamic mechanism of diverse pathways for control and integration of metabolic pathways. The effect of tripping a sulfur switch, by affecting orientation or tertiary structure of a functional site, is transduced via ligand binding/release, gene expression/silencing, or enzyme activation/inactivation into metabolic effect (86, 150). This redox system also offers a new approach to the study of diet and health. Biological sites may be affected by dietary antioxidants through: a) general changes in redox tone; b) protection or activation of specific redox-sensitive sites; c) effects on availability of other electron donors (6, 55, 77, 168, 179). For example, a key controlling mechanism in enzyme action is a loop of phosphorylation/ dephosphorylation, often at tyrosine residues, (34, 86, 147). Several protein tyrosine phosphatases have been reported to be regulated by redox changes, and protein kinase B (PKB) is regulated by a redox-sensitive disulfide bridge in an activation loop (179).
ROS signaling and cellular effects: An overview
ROS affect kinases and phosphatases, key agents for translating signals of cell needs into metabolic change through enzyme action, gene transcription, protein synthesis, and biochemical flux (1, 86, 165). ROS (and RNS) activate many kinases, including protein kinase C (PKC), protein kinase B (PKB), phosphatidylinositol 3-kinase (PI3-kinase), mitogen-activated protein kinase (MAPK), and protein tyrosine kinase (PTK) (1, 86, 94, 145). Kinase effects are mediated also by ROS effects on tumor necrosis factor alpha (TNF-α) which in turn activates protein kinases such as the MAPK apoptosis signal-regulating kinase 1 (ASK1). Phospholipase (PL) activity is triggered by ROS through effects on tyrosine phosphorylation, and activated PL cleaves membrane-bound phospholipids producing second messengers such as diacylglycerol (DAG) and inositol 1,4,5-triphosphate (IP3). Modulation of phosphatase activity, the other side of the coin of control of enzyme activity, also involves ROS participation, and activation of protein kinases is due in part to ROS inactivation of phosphatases (1, 35, 165, 180).
Superoxide and hydrogen peroxide are involved in protein kinase regulation, while nitric oxide and peroxynitrite inactivate phosphatases as well as activating kinases (1) There is some evidence for specificity of response: superoxide was reported to cause a dose-response increase in MAPK activity in cultured vascular smooth muscle cells (VSMC), while hydrogen peroxide showed no effect, and superoxide caused proliferation of VSMC while hydrogen peroxide induced apoptosis. Hydrogen peroxide is believed to be more important than superoxide as a signaling molecule. Small and uncharged, hydrogen peroxide diffuses within and between cells, is not highly reactive, and is found in millimolar quantities at some biological sites (73, 177), characteristics that favor its use as a signaling agent, though its lack of reactivity might blunt direct effectiveness. Conversely, the more reactive superoxide is negatively charged, with low mobility and short half-life, features that appear to limit use as a signal molecule, although it has the advantage of being able to act as oxidant or reductant (70). The major site of superoxide and hydrogen peroxide formation is mitochondria (150), but superoxide can be formed in cytoplasm by ‘reverse dismutation’ of hydrogen peroxide, catalyzed by copper-zinc SOD (1). This offers an elegant mechanism whereby superoxide in a relatively inactive, long-lived and easily diffusible ‘partner’ form (hydrogen peroxide) reaches distant sites before being transformed back into superoxide and used for signal transduction. The signal process may be via interaction with protein, or with nitric oxide and subsequent production of a different signaling molecule, peroxynitrite, which may activate or inhibit different pathways. This remains to be confirmed. Superoxide is produced also during the very kinase activation reactions it helps transduce, suggesting ‘feed forward’ loops in a chain of kinase activation (1, 55, 150). However, eventually there has to be a feedback or inhibitory mechanism that breaks this chain and turns the signal off. Inhibitors could be in the form of scavenging antioxidants, such as ascorbic acid or polyphenols.
The primary role of nitric oxide is blood pressure control (70), but it can inactivate phosphatases by nitration of key cysteine residues, and peroxynitrite activates JNK kinase, though how much signaling is done by nitric oxide and how much relies on peroxynitrite is not clear (1). Peroxynitrite is more reactive and able to interact with proteins, which could make it a more effective signal molecule, but removal of superoxide by interaction with nitric oxide may also offer a means to control direct signal transduction effects of superoxide. The superoxide/nitric oxide reaction is many times faster than that of superoxide/ascorbic acid (70), but higher amounts of ascorbic acid, and possibly other dietary antioxidants, may modulate availability of both ROS and limit their signaling effects.
Effects on kinases and phosphatases are not the only routes by which redox changes determine the biochemical profile and fate of the cell. For example, the redox tone of the endoplasmic reticulum (ER) determines oxidative folding in secretory proteins. This is largely determined by the activity of the flavoprotein Ero1 oxidases, which generate ROS and deplete GSH (6). The ubiquitous Ero1α form contains a sensitive sulfur switch, tripped by a change in redox state of its thioredoxin substrate, protein disulfide isomerase (PDI), and acts as a regulatory feedback mechanism in ER redox control (6, 86). Thioredoxins and thioredoxin reductase are key players in redox control of cellular events, and are themselves affected by redox tone (8, 86). Proteases, including caspases, contain critical cysteine residues and are also regulated by redox changes (86). In addition to this redox link to apoptosis, changes in redox tone affect cell-cycle progression and inter-cell communication (34, 35, 102). Gene activation and transcription are also modulated by redox tone (1, 6, 86, 94). In the following section, a particular redox sensitive gene promoter region, the Antioxidant Response Element (ARE), is reviewed and influence of dietary antioxidants examined.
The Antioxidant Response Element and dietary antioxidants
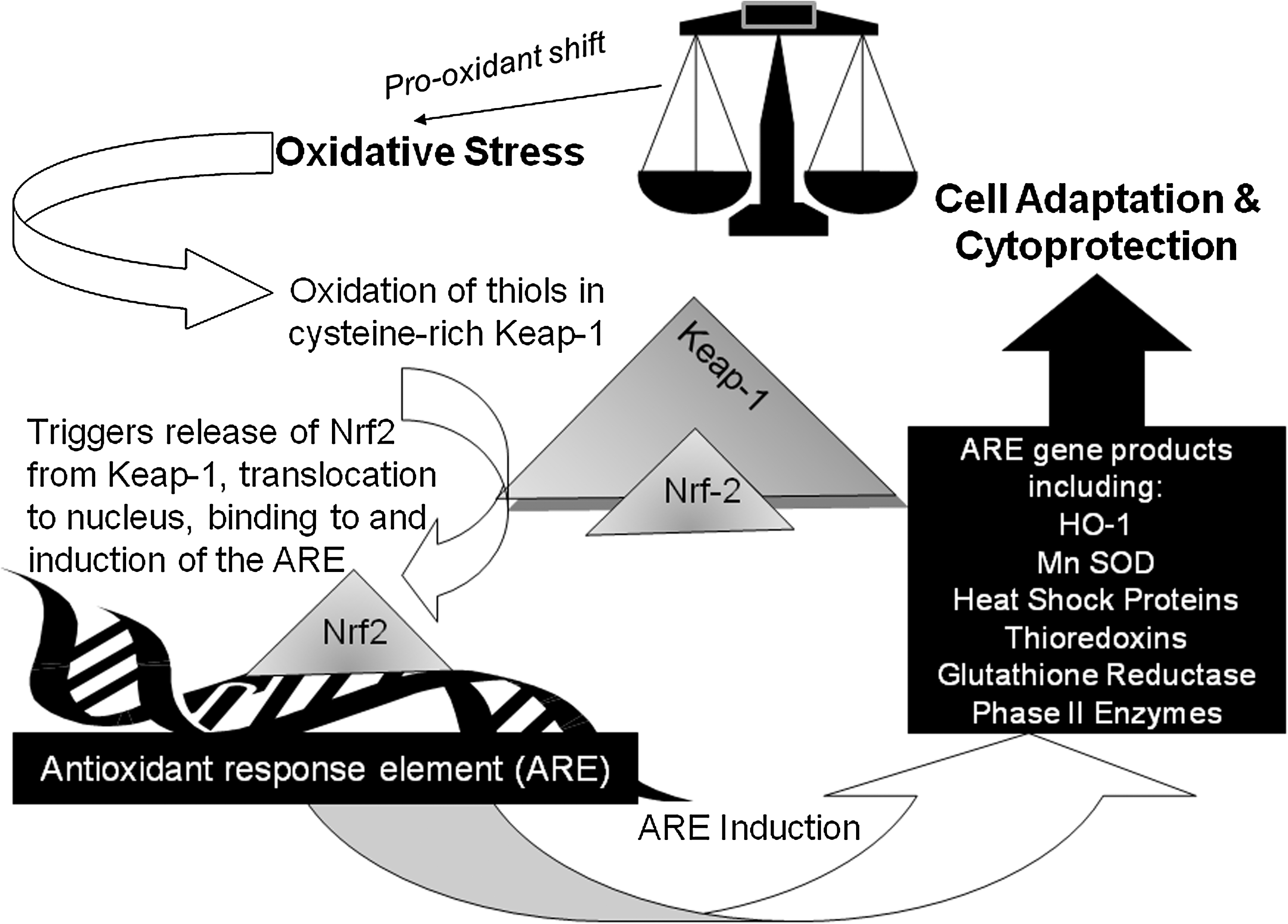
The antioxidant response element (ARE, also known as an electrophile-responsive element), is a redox-sensitive gene promoter region (63, 68, 107, 124, 153). The ARE regulates genes encoding Phase II enzymes and other products involved in cytoprotection and antioxidant defense, including heat shock proteins, antioxidant enzymes, thioredoxins, and thioredoxin reductases (107, 117, 149, 178). These pro-survival genes have been termed ‘vitagenes' (36). A pro-oxidant change in redox tone (‘oxidative stress') activates the transcription factor NF-E2-related factor 2 (Nfr2) (116, 153, 178). This is kept inactive within the cytoplasm by binding to the cysteine-rich protein, Keap-1 (Kelch-like ECH-associated protein 1). Keap-1 acts as a sensor of redox change due to reversible covalent modifications of its thiol groups. These are oxidized when the protein is exposed to oxidants and electrophiles, including the electrophilic lipid oxidation products 4-hydroxynonenal (HNE) and the J series of the cyclopentenone prostaglandins (J series CyPGS), releasing Nrf2 from thiol-modified Keap-1 (66, 86, 107, 116, 125, 153, 178, 180). Released Nrf2 translocates to the nucleus where, in a dimeric form with basic region leucine zipper (bZIP) proteins, it binds to and activates the ARE (107, 124, 153). Two critical reactive cysteine groups, Cys273 and Cys288, were identified for the redox regulation of Keap-1-binding of Nrf2, although effects of thiol changes in other cysteine residues of Keap-1 may also contribute to its regulatory function, and lipid oxidation products with two electrophilic β-carbons are reported to be more potent activators of ARE than those with only one such carbon (107, 180). Rapid modification of Keap-1 thiols has been reported also in cells exposed to nitric oxide (33). In addition to inhibiting ARE activation by binding Nrf2, Keap-1 keeps Nrf2 levels low by targeting it for ubiquination and enhancing its degradation in what is also a redox-sensitive process (84, 116). Upstream kinase action on Nrf2 is also important for stabilization of the protein (153). MAPK, PI3K, and PKC are also involved in ARE activation of certain genes and/or cell type (1, 124, 178). Signaling pathways are often controlled by action of tyrosine phosphorylation/dephosphorylation, and protein tyrosine phosphatase inactivation due to oxidation of active site cysteine (or possibly histidine or lysine) residues could lead to prolonged activation of MAPK and PI3K/Akt pathways and upregulation of ARE-activated genes (1, 124, 153).
Key products of the ARE are the heme oxygenases (HO-1, HO-2, and HO-3), which catalyze degradation of hemoglobin to ferrous iron, biliverdin, and carbon monoxide (CO) (51, 76, 83, 149). Increased HO-1 is a marker of adaptive response to and is protective against oxidative stress (83, 149). Indeed, HO-1 induction has been termed a ‘therapeutic funnel’ (149) because various synthetic drugs are thought to work by their ability to induce HO-1. Bilirubin (from biliverdin) is reported to be a powerful antioxidant, and CO reportedly inhibits apoptosis and has anti-inflammatory, anti-proliferative, vasodilatory, and angiogenic effects (82, 138, 149). HO-1 releases iron from degraded hemoglobin, but upregulates expression of ferritin, a cytoprotective action, as ferritin binds and oxidizes ferrous to ferric iron (149). The effects of HO-1 are different in different cells. For example, HO-1 inhibits VSMC proliferation and promotes apoptosis, with opposite effects seen in endothelial cells, although the overall effect of increased HO-1 in these cells is protection against atherosclerosis (176). Figure 7 outlines the Nrf2-Keap-1 system of ARE induction and its cytoprotective products.
Most confirmed ARE-inducers are endogenous products or synthetic agents that induce oxidative stress (53). However, the ARE is induced by some phytochemicals, including the sulfur-containing nonphenolic compounds sulfurophane (found in cruciferous vegetables) and diallyl trisulfide (from garlic), and the polyphenolic antioxidant compounds quercetin (found in tea, onions, and apples), epigallocatechingallate (EGCG; from tea) caffeic acid phenethyl ester (from honey), curcumin (from turmeric), genistein (from soya bean), carnosol (from rosemary), capsaicin (from chili pepper), resveratrol (from grape skin and seeds), and phenolic compounds from ginger and hops (53, 136, 153). Effects on the ARE offer a molecular rationale for the health benefits of vegetarian diets that is distinct from simple antioxidant action. However, high intake of plant foods containing ARE inducers would involve high intake of antioxidants, a situation believed to relieve oxidative stress, inhibiting ARE induction. How would this removal of cytoprotection benefit the cell? Furthermore, how do antioxidant phytochemicals induce the ARE? It may be through different mechanisms than used by oxidants, such as promoting stabilization of Nrf2. However, oxidative stress is known to induce ARE. How can we reconcile this with the established antioxidant properties of ARE-inducing phytochemicals, and with the health benefits of antioxidant-rich foods?
There are very few studies of effects of phytochemicals on products of ARE induction. Lycopene at 2–4 μmol/L was reported to induce the ARE in cultured cancer cells (153), but in human studies, lycopene supplementation did not change HO-1 protein expression in lymphocytes, although less apoptosis (p < 0.01) was seen (115). Curcumin (from turmeric) increased HO-1 in astrocytes and in vascular endothelial cells (36). In an animal study, vitamin E (administered as α-tocopheryl acetate) had no effect on HO-1 induction (or on atherosclerosis), but probucol, a potent synthetic antioxidant, increased HO-1 and inhibited atherosclerosis (176). Both vitamin E and probucol contain a phenol group capable of donating one electron in a redox reaction. Probucol also contains a sulfur group that can donate two electrons. The authors concluded that sulfur groups may be critical as redox-active compounds, but noted also that probucol did not appear to directly induce ARE, and speculated that a metabolite of probucol, formed by a two-electron redox reaction, was responsible (176). The metabolite and its mechanism of action remain unknown, but a two-electron oxidation product could exert the oxidative challenge needed for Nrf2 release and subsequent ARE induction. Therefore, some of the protective effects of dietary phytochemicals could be related to cellular adaptations to direct or indirect pro-oxidant effects. Antioxidants can act as pro-oxidants (23, 71), though whether they do so in vivo is not known and difficult to investigate due to sample and biomarker limitations. Oxidation of phenolic compounds in cell culture medium produces hydrogen peroxide (73). This may be true also in the gut and bladder, and fresh urine contains significant amounts of hydrogen peroxide (70, 73, 177). In vitro studies show that high doses of antioxidants can be cytotoxic, that low doses protect DNA, and both protection and cytotoxicity are prevented by catalase, indicating hydrogen peroxide involvement (73, 170). Therefore, ARE induction and subsequent cytoprotection could result from adaptive responses to a paradoxical pro-oxidant effect of antioxidants, particularly of phytochemicals that contain sulfur or have multiple electron donating groups. This could explain why antioxidant supplementation trials have not shown benefit. If oxidative stress is decreased and the redox tone shifts to the antioxidant side, the effects of such ‘reductive stress' might be damaging. The ‘antioxidant’ response element in fact responds to pro-oxidant shift in redox tone and induces antioxidant and other means of cell protection (124, 149). There is no known protective equivalent that responds to a reductive shift in redox tone. But in the scenario suggested above this is not needed. The feed forward and feedback loops to the two sides of the same balancing system are enough. That is, we can speculate that antioxidants modulate both sides of the same scale by dual action that varies with conditions within the dynamic nonequilibrium redox system (Fig. 8).
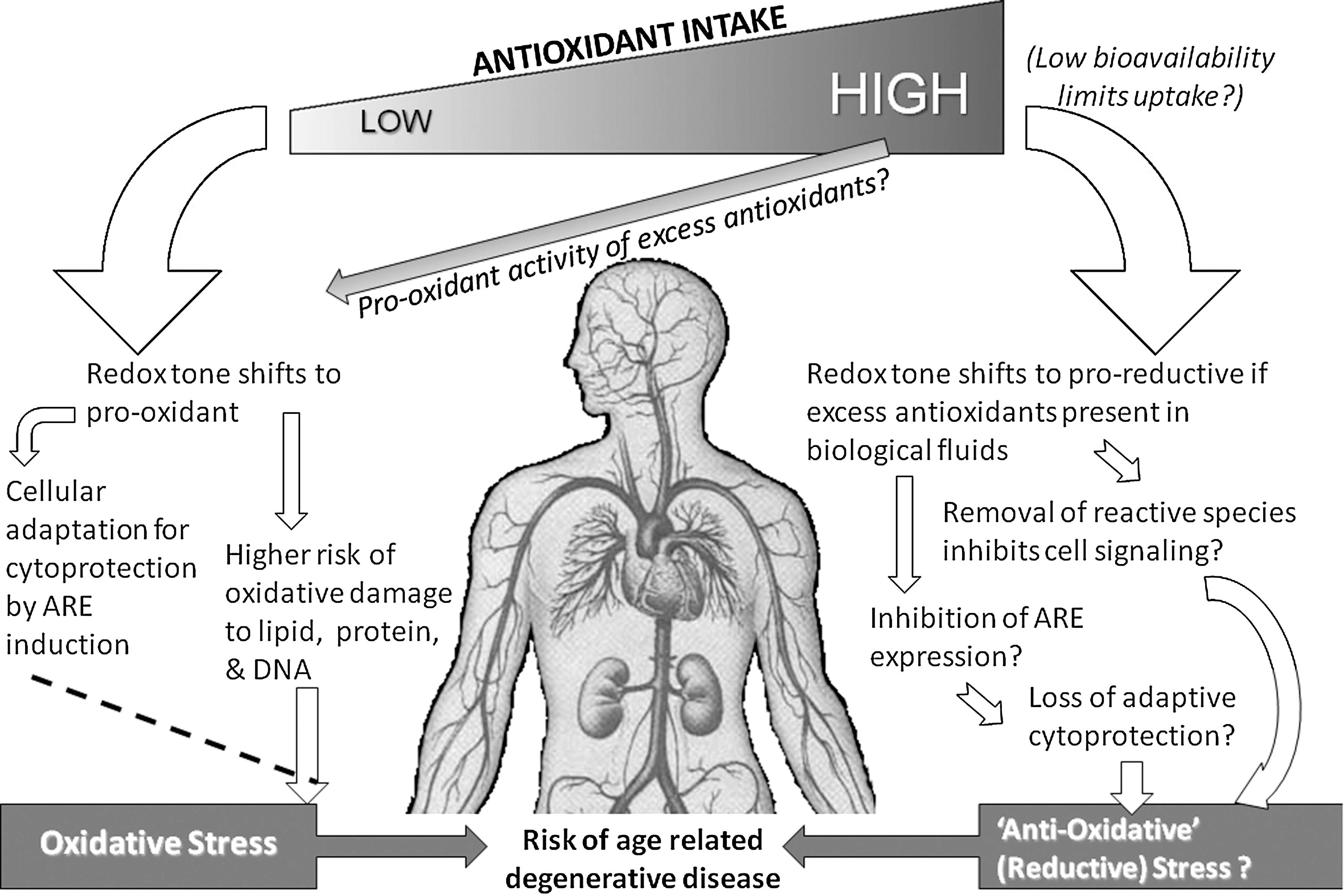
Currently, there is little in the way of direct evidence that dietary antioxidants do in fact act as pro-oxidants in vivo, thereby inducing the ARE and benefiting health through ARE-associated cytoprotective adaptations to a mild pro-oxidant shift in redox tone. Furthermore, it is difficult to investigate this concept in a physiologically relevant way. For example, in cultured primary rat hepatocytes, resveratol at 50 and 75 μmol/L was shown to increase Nrf2 and its translocation to the nucleus, and to increase ARE products (antioxidant enzymes and Phase II enzymes) (136). However, these doses are much higher than achieved in vivo. Studies of flavonoids and isoflavones in cultured cells have shown genomic induction of antioxidant defense genes (reviewed in Ref. 148). However, cell culture studies suffer from serious drawbacks, and as stated by Janssen–Heininger et al. (86), the study of redox-based regulation of signal transduction is an area full of pitfalls, as well as promises. Cultured cells are generally transformed, depleted of antioxidants, and hyperoxygenated. Further, addition of whole or extracted foods cannot emulate the doses or biotransformations that represent the in vivo situation. Virgili and co-workers attempted to address this by using human subjects as ‘bioreactors', using post-ingestion plasma from human subjects as test material (169). Using this approach, native (as it would be drunk) and ‘biotransformed’ red wine (i.e., post-ingestion plasma) had very different effects when added to endothelial cells. Effects on the ARE were not investigated, but biotransformed red wine induced nuclear translocation of the transcription factors NFκB and AP-1 and decreased expression of adhesion molecules, effects not seen with native red wine (169). This novel approach will be useful in investigation of effects of ‘native’ and ‘biotransformed’ phytochemicals on the ARE and its products.
It is unlikely that ARE induction produces measurable changes to plasma concentrations of products such as CO and biliverdin/bilirubin (though this may be worth investigating), and in human studies, white blood cells are the only easily accessible nucleated cells. In a controlled human trial with a food (passata sauce), plasma lycopene was significantly increased (from an average of 0.20 to 0.54 μmol/L; p < 0.05) after 3 weeks supplementation, but this was not associated with changes in basal or hydrogen peroxide-induced HO-1 protein levels in lymphocytes (115). However, as a quenching antioxidant, lycopene might not induce the ARE if this requires sulfur-containing or multiple electron donating antioxidants. It has to be noted also that phytochemical effects on Nrf2/ARE (53, 136, 153, 169) have been obtained largely from high-dose cell culture studies. The only dietary studies (153) were with curcumin and cafestol (a diterpene from coffee) in rodents, which increased translocation and ARE binding of Nrf2 and increased expression of some ARE products. More studies of this type are needed, and new technologies and redox proteomic methods will help (86, 88). With such tools it may be possible to focus on activity of key sites of cysteine oxidation, on kinases or phosphatases, and on activities of GR and TrxR. Reversible cysteine oxidation and the recycling action of peroxiredoxins and thioredoxins are now known to play critical roles in governing redox balance and signaling (86, 89, 109). Effects of dietary antioxidants on these will be a rewarding area of study.
Other molecular mechanisms and effects of dietary antioxidants
Non-antioxidant effects on proteins, such as FOXOs and sirtuins, membranes, enzymes, and hormones are under study (32, 100, 103, 145 –148, 168, 169, 179). FoxO proteins are a family of forkhead transcription factors of the O class that control cell cycle progression and promote cell death by activating pro-apoptotic genes (113, 139). FoxOs are targets of various stimuli, including oxidative stress. Sirtuins (SirTs) are a family of seven NAD-dependent enzymes that are referred to as ‘longevity proteins' because they increase expression of proteins that extend cell survival and silence FoxO (119, 121). Pre-B cell colony-enhancing factor (PBEF ) is the rate-limiting enzyme in NAD synthesis, and PBEF activates SirT1 by supplying NAD (121). Resveratrol is reported to activate SirT1 (113). SirT1 interacts directly with FoxO1, deacetylating it and modulating its transcriptional effects. White wine was shown to have potent effects on the longevity proteins, SirTs, FoxOs and PBEF, followed by resveratrol > tyrosol > red wine > hydroxytyrosol. However, the magnitude of effects on cardiomycoyte apoptosis and infarct size following ischemia reperfusion was different (resveratrol > hydroxytyrosol > red wine > white wine > tyrosol), suggesting that survival and anti-aging pathways may be different, and that effects of the phenolic components of wine tested do not depend on the number of hydroxyl groups (121). Catechins and flavanols did not show these effects. This could indicate that antioxidant activity is not a feature of these cellular effects seen with wines. Nonetheless, the effects on PBEF, Sirt1, and FoxO are pro-longevity effects, implying that red wine, white wine, and at least some of their phenolic components bring benefits to health via these molecular effects (121). EGCG was also reported (145) to activate FOXO, this being mediated by inhibition of the P13K/AKT and MEK/ERK pathways (which could have been through a redox-related effect), and had downstream anti-angiogenic effects. However, this was a cell culture study (with HUVEC) and high (40 μmol/L) dose of EGCG was used.
Other mechanisms of phytochemical action relate to cell-junction gap communication, binding to receptors, modulation of hormone action, and response to ligand binding. Resveratrol protected cultured rat liver epithelial cells against ROS-induced inhibition of gap-junction intercellular communication by inhibiting phosphorylation of ERK1/2 (extracellular signal-related protein kinase 1/2), and this was reportedly not driven by antioxidant action (based on in vitro antioxidant testing) (102). In cultured human cancer cells, EGCG from green tea is reported (146, 147) to inhibit receptor tyrosine kinases, thereby inhibiting MAPK- and PI3K-associated signaling pathways. Green tea polyphenols modulate cellular signaling pathways during inflammation and can themselves serve as signaling agents (100, 145 –147, 153, 160). Green tea polyphenols also inhibit cycloxygenase 2 (COX-2) and inducible nitric oxide synthase (iNOS) expression by blocking NFκB activation, and EGCG was shown to prevent death of myocytes following ischemia by blocking activation of inflammation-related compounds, including NFκB (160). EGCG is reported to affect multiple signaling pathways, including redox sensitive (e.g., NFκB, AP-1, and MAPK pathways) and nonredox receptor mediator pathways, such as epidermal growth factor receptor (EGFR)-mediated and insulin-like growth factor-1 (IGF )-mediated signal transduction pathways (100). Flavonoids can displace estradiol and bind to estrogen receptors, and activate other nuclear receptors called ‘orphan receptors' and ‘adopted orphan receptors', such as peroxisome proliferator-activated receptors (PPARS) (169).
Vitamin C is known to affect the activity of many enzymes through its antioxidant activity (48, 57, 168). However, vitamin E, regarded as the major lipophilic antioxidant (161, 162), has been shown to inhibit some enzymes and activate others in ways that are not antioxidant-related (32, 168, 179). Inhibitory effects of vitamin E have been seen on NADPH oxidase, PLA2, PKB/Akt, PKC, lipoxygenase, and COX-2, and activation effects are reported with DAG kinase and hydroxymethylglutaryl-Coenzyme A (HMG-CoA) reductase (32). Mechanisms are varied, for example, α-tocopherol can bind directly to some enzymes (e.g., PLA2), inhibit phosphorylation-dependent activation (e.g., PKC; although this could be a redox effect), and affect membrane translocation of other enzymes (e.g., PKC, NADPH oxidase) (32, 179). Diacylglycerol (DAG) kinase and HMGCoA reductase are activated specifically by α-tocopherol, and this does not appear to be related to its antioxidant function (32, 179). Interestingly, α-tocopherol diffuses laterally through the lipid bilayer of membranes and rotates around so that its phenolic head can face inwards or outwards from the membrane (32). However, it is not found randomly situated, but within complexes with particular membrane constituents, suggesting it may help form or stabilize lipid rafts, active membrane microdomains for cell signaling and trafficking (32). In short, the many and varied actions and activities of the vitamin E family cannot be laid at the door of simple antioxidant action as previously thought (161). Indeed, the role of vitamin E as a lipid-soluble antioxidant in human biology is in doubt (32). Instead, a key role for α-tocopherol appears to be in regulating genes and processes involving cell membrane function and arrangement, and affecting adhesion, fusion, phagocytosis and, crucially for inter-cell signaling, transmitter release (32, 179). Further, rather than acting as a defense against oxidation, interaction of α-tocopherol with lipid peroxides within membranes may act as an oxidation sensor that triggers more mobile ‘second messengers' in a signaling cascade (180). This changing face of vitamin E requires further study before it can be recognized fully.
Concluding Remarks
Research into the molecular effects of antioxidant phytochemicals in relation to redox tone and cell signaling is in its early stages, but is needed to confirm the mechanisms by which antioxidant-rich vegetarian diets benefit health. To date there are very few data from physiologically-relevant experimental settings, and human studies are lacking. It is clear that the idea that a diet rich in antioxidant phytochemicals is beneficial simply through increased radical scavenging is unsustainable. Some phytochemicals do have direct antioxidant action, but others that have in vitro antioxidant activity may not act as antioxidants in vivo, and yet still have important signaling and regulatory functions. The feed-forward and feed-back loops that control redox tone and the action and interplay of dietary antioxidants in cell signaling are not yet known. This is a rich area for research to advance understanding of orthomolecular nutrition and provide science-based evidence to advance public health and promote well-being in our aging population.
Footnotes
Acknowledgment
The authors are grateful to The Hong Kong Polytechnic University for supporting this work.