
Abstract
The 26S proteasome is a large proteolytic particle present in the cytosol and nucleus of eukaryotic cells. Most intracellular proteins, including those affected by oxidative damage, are degraded by the proteasome. The human thioredoxin, Txnl1, is known to associate with the 26S proteasome and thereby equips proteasomes with redox capabilities. Here, we characterize the fission yeast orthologue of Txnl1, called Txl1. Txl1 associates with the 26S proteasome via its C-terminal domain. This domain is also found in the uncharacterized protein, Txc1, which was also found to interact with 26S proteasomes. A txl1 null mutant, but not a txc1 null, displayed a synthetic growth defect with cut8, encoding a protein that tethers the proteasome to the nuclear membrane. Txc1 is present throughout the cytoplasm and nucleus, whereas Txl1 co-localizes with 26S proteasomes in both wild-type cells and in cut8 mutants, indicating that Txl1 is tightly associated with 26S proteasomes, while Txc1 might be only transiently bound to the complex. Finally, we show that Txl1 is an active thioredoxin. Accordingly, Txl1 was able to reduce and mediate the degradation of an oxidized model proteasome substrate in vitro. Thus, Txl1 and Txc1 are proteasome co-factors connected with oxidative stress. Antioxid. Redox Signal. 14, 1601–1608.
Introduction
The 26S proteasome is a rather stable particle (26), which can be dissociated into two subcomplexes, the 20S core proteasome and the 19S regulatory complex. The latter complex associates with the ends of the cylindrical core proteasome. The 20S cylinder is made up of four stacked rings. Seven different α subunits form each of the two identical outer rings of the cylinder and seven different β subunits form each of the two identical inner rings, yielding a twofold symmetric structure as: α1–7β1–7β1–7α1–7 (13).
With few exceptions, proteins must be conjugated to a chain of ubiquitin before they become substrates for the 26S proteasome. Once bound to the proteasomes, substrates are unfolded under consumption of ATP (4, 32, 38) and threaded into the hollow interior of the core proteasome, where they are exposed to the β subunits with endopeptidase activity. The substrate recognition (7, 43) and unfolding (4, 32, 38) are mediated by the 19S regulatory complex, which is also responsible for the release of ubiquitin since several ubiquitin hydrolases are associated with the 19S particle (3, 7, 15, 19, 25, 37, 42).
Recently, we and others found that the human thioredoxin-like protein Txnl1 is associated with the 26S proteasome (1, 46). Txnl1 is phylogenetically well conserved and present in most eukaryotes, including the fission yeast Schizosaccharomyces pombe. However, budding yeast seems not to encode any Txnl1 orthologue.
Here, we characterize the Txnl1 orthologue, Txl1, in fission yeast. We show that Txl1 associates with 26S proteasomes, and we isolate the previously uncharacterized protein Txc1 as a novel proteasome-associated protein. Null mutants in txl1 and txc1 do not display any growth defects, but Txl1 is an active thioredoxin that can reduce and mediate the degradation of an oxidized model proteasome substrate in vitro. Hence, although oxidative stress and intracellular protein degradation have been linked for years (23), Txl1 and Txc1 are new examples of proteins involved in both processes.
Materials and Methods
S. pombe strains and techniques
The fission yeast strains used in this study (wt, mts3-1, pad1-ProtA-ZZ, txl1::NAT, txc1::G418, trx1::G418, moe1::G418, and cut8::G418) are derivatives of the wild-type heterothallic strains 927h− and 927h+ . Standard genetic methods and media were used, and S. pombe transformations were performed using the lithium acetate procedure (31). The PCR mutagenesis was performed as described previously (2). Some deletion mutant strains were obtained from Bioneer (Alameda, CA).
Plasmids and PCR
To generate Txl1 (SPBC577.08c)- and Txc1 (SPBP35G2.02)-encoding constructs, full length cDNAs were amplified from an S. pombe cDNA preparation and inserted into pDONR221, using the Gateway cloning technology (Invitrogen, Carlsbad, CA). From the pDONR221 vector, the inserts were transferred to Gateway-compatible vectors for E. coli and S. pombe expression (30). Site-directed mutagenesis was performed using the QuikChange kit (Stratagene, Cheshire, UK).
Using Nucleobond AX (Macherey–Nagel, Bethlehem, PA), mRNA was isolated from untreated wild-type cells or cells treated with either 0.5 mM H2O2, 39°C heat shock or 1 M sorbitol for 1 h. The mRNA was treated with Turbo DNA-free (Ambion, Foster City, CA) as described by the manufacturers and then reverse transcribed with the Transcriptor First Strand cDNA Synthesis (Roche, Basel, Switzerland), before regular PCR, using gene-specific primers.
Binding assays
GST fusion proteins were expressed in E. coli BL21*(DE3) (Invitrogen) and bound to glutathione Sepharose 4 beads (GE Healthcare) as described by the manufacturer. The protein/bead ratio was 1 mg/ml. Binding experiments were carried out using 20 μl of beads in 1 ml of 50 % wild-type fission yeast extract in buffer A [25 mM Tris/HCl pH 7.5, 50 mM NaCl, 2 mM MgCl2, 5 mM ATP, 10% glycerol, 0.1% Triton X-100, 2 mM DTT, 1 mM PMSF, and CompleteTM protease inhibitors (Roche)]. After 2 h to overnight tumbling at 4°C, the beads were washed 4 times in buffer A and resuspended in 30 μl of SDS sample buffer. Samples of 10 μl were then separated on 12% SDS gels and subjected to immunoblot analysis.
For in vivo binding assays, 50% extracts in buffer A of S. pombe cells, expressing 6His- and Flag-tagged Txl1 or Txc1, were incubated with 20 μl anti-Flag resin (Sigma, St. Louis, MO) per 5 ml of extract for at least 4 hours at 4°C. The precipitate was then washed and analyzed by immunoblotting as above.
For isolating Txl1 mixed disulfide intermediates, txl1::NAT S. pombe cells, transformed to express Flag- and 6His-tagged Txl1C34S or untagged Txl1C34S, were lysed and incubated with anti-Flag resin in buffer A as described above, but without addition of DTT. After washing, the precipitated material was eluted by addition of buffer B (86 mM Na2HPO4, 14 mM NaH2PO4, 10 mM Tris/HCl pH 8, 8 M urea, and 2 mM N-ethylmaleimide). The eluted material was then subjected to a second round of precipitation, now using 20 μl of Ni2+-NTA agarose (Qiagen, Alameda, CA). After 4 h of tumbling at room temperature, the beads were washed 4 times with buffer B and 4 times with PBS. Finally, the pellet was resuspended in 40 μl of SDS sample buffer and resolved by reducing and nonreducing SDS-PAGE and immunoblotting.
The anti-sera, used in immunoblots, were affinity purified rabbit polyclonal anti-Mts4 (Rpn1), sheep polyclonal anti-Uch2, mouse monoclonal MCP231 anti-proteasome α subunits (Enzo Life Sciences, Plymouth, PA), goat polyclonal anti-GST (Abcam, Cambridge, UK), mouse monoclonal anti-Flag (Sigma), rabbit polyclonal anti-ubiquitin (Dako, Glostrup, Denmark), and mouse monoclonal anti-penta-His (Qiagen).
Growth assays
The strains to be assayed were grown to an OD600nm of about 1.0. The cells were then washed in water and resuspended in water to an OD600nm of exactly 0.40. Serial 5-fold dilutions of this culture were prepared before 5 μl of each dilution was spotted onto solid media. Unless otherwise indicated, the plates were incubated at 30°C until colonies formed.
Fluorescence microscopy
Immunocytochemistry was performed as described (45) using cells transformed to express GFP-tagged Txl1 or Txc1 and affinity purified polyclonal rabbit antibodies against Mts4 (Rpn1).
Assays
Concentrations of purified recombinant proteins were determined from A280nm. The thioredoxin activity of immunoprecipitated Flag-tagged Txl1 and as a control thioredoxin from E. coli (Sigma) was determined as described (18). Degradation assays with α-lactalbumin (Sigma) were performed as described (44).
Results
Txl1 and Txc1 are fission yeast orthologues of human Txnl1 and C1ORF128, respectively
Recently, we found that the human thioredoxin-like protein Txnl1 associates with the 26S proteasome via its C-terminal domain (1, 17), which according to the Pfam database is termed a proteasome-interacting thioredoxin (PITH) domain. Curiously, budding yeast does not seem to encode any orthologue of Txnl1. However, S. pombe encodes txl1+, which shows 34% identity to human Txnl1 (Fig. 1A). The active site CGPC motif is conserved (Fig. 1A).
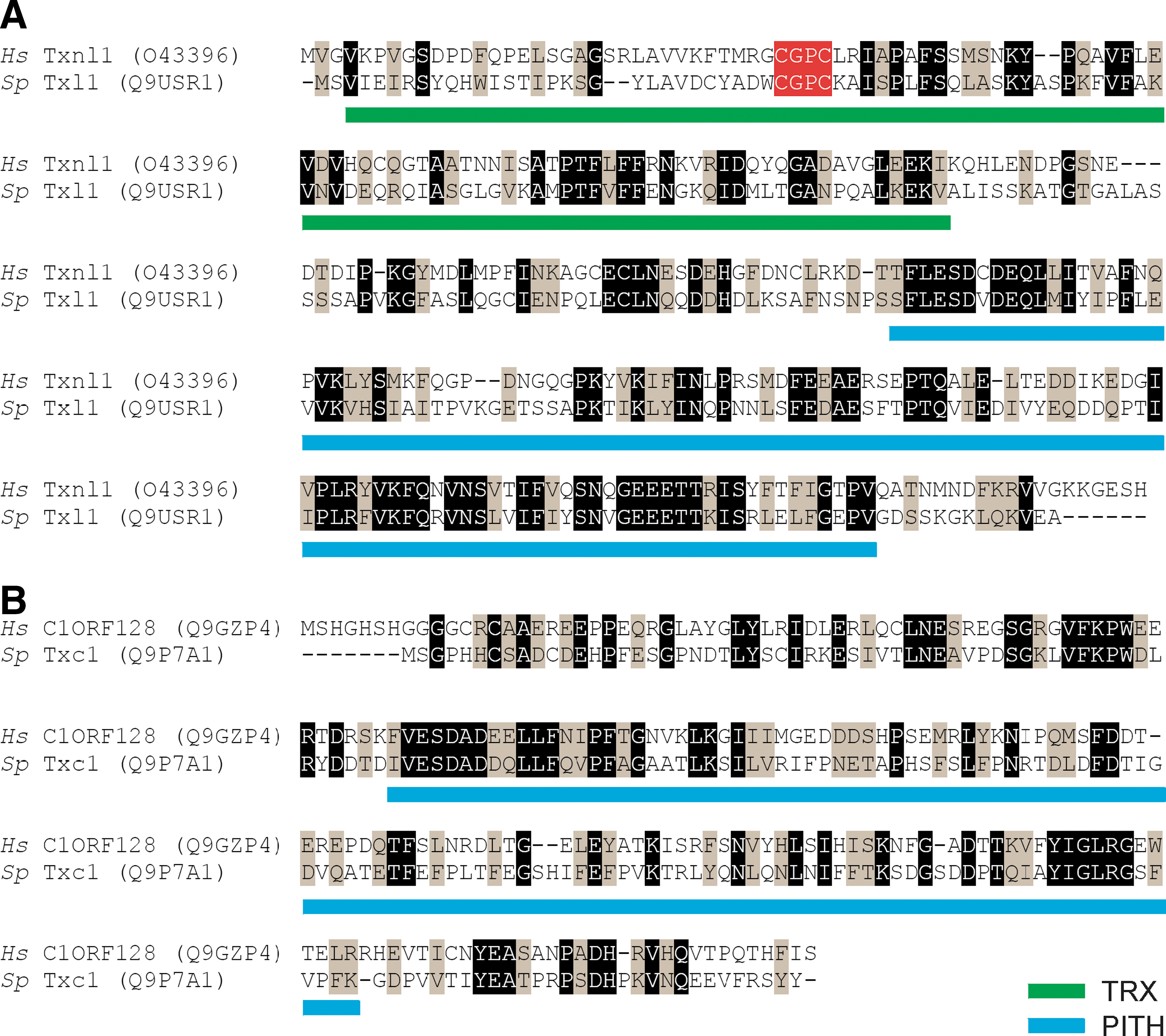
In humans, the uncharacterized gene C1ORF128 (also named ht014 or Trp26) encodes the only other known PITH domain protein. As with Txnl1, no orthologue of this gene is found in budding yeast. However, S. pombe encodes the uncharacterized gene, SPBP35G2.02, which shows 32% identity to human C1ORF128 (Fig. 1b). We therefore named SPBP35G2.02 as Txc1 for Txl1 C-terminus.
Txl1 and Txc1 associate with 26S proteasomes
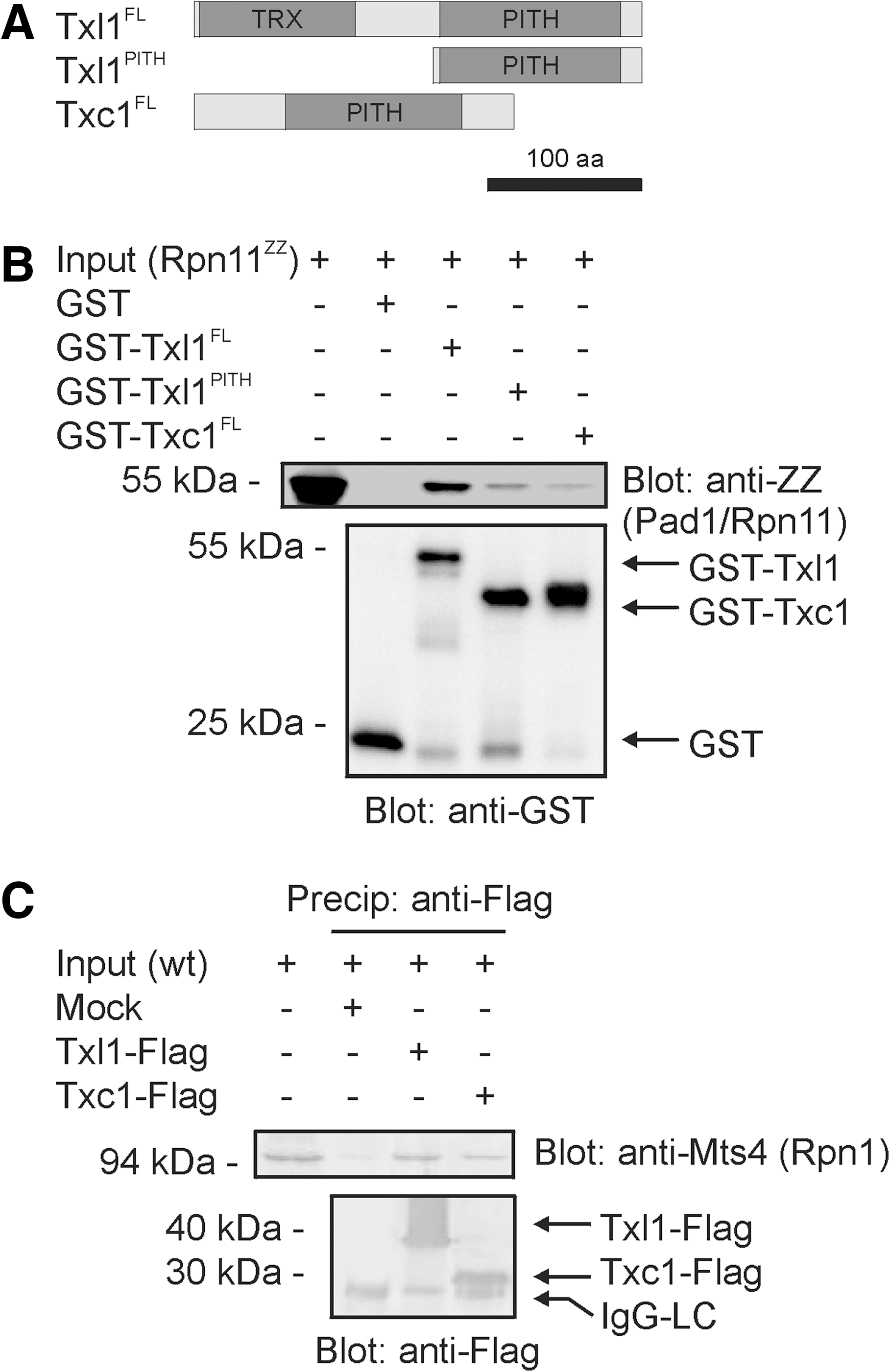
In order to determine whether Txl1 and Txc1 associate with 26S proteasomes, we expressed GST and full length (FL) Txc1, Txl1, and the Txl1PITH truncation as GST fusion proteins in E. coli (Fig. 2A). The GST-tagged proteins were purified and incubated with extracts from S. pombe cells expressing the Protein A ZZ tagged 26S proteasome subunit Pad1/Rpn11. The precipitation experiments showed (Fig. 2B) that Txl1 and Txc1 both interact with the 26S proteasome, and that the PITH domain was sufficient to mediate this interaction.
In order to verify that Txl1 is not simply associated with 26S proteasomes as a substrate, we also mixed purified 6His-tagged Txl1 with 26S proteasomes and analyzed the degradation. After 8 hours, we did not observe any measurable degradation of Txl1 (Supplementary Fig. 1; see
The human orthologue of Txl1 is associated with the 19S regulatory complex of the 26S proteasome (1, 46). In order to determine the interaction site of Txl1 and Txc1, precipitation experiments using GST, GST-Txl1, and GST-Txc1 were performed on intact 26S proteasomes and 26S proteasomes that had been dissociated into the 19S and 20S complexes by ATP-depletion and treatment with salt. When intact 26S proteasomes were used, the GST-Txl1 and GST-Txc1 fusion proteins could precipitate both 19S and 20S components (Supplementary Fig. 2; see
In order to confirm the interaction between Txl1 and the 26S proteasome in vivo, Flag-tagged Txl1 was immunoprecipitated from S. pombe cells. Immunoblotting revealed that endogenous 26S proteasomes co-precipitated with Txl1 (Fig. 2C). Immunoprecipitation of Flag-tagged Txc1 revealed that Txc1, like Txl1, also associates with 26S proteasomes in vivo (Fig. 2C). Thus, both Txl1 and Txc1 interact with the 26S proteasome and the PITH domain is therefore probably a general proteasome-interacting domain.
Gene expression
Previous reports on Txl1 in S. pombe and other organisms indicate that its expression may be regulated at the transcriptional level in response to cell stress (24, 29, 41). To test this, we performed reverse-transcriptase PCR on wild-type cells exposed to either H2O2, heat shock, or sorbitol. Expression of txl1
+ and txc1
+ was not significantly affected compared to the control, whereas trx1
+ expression was visibly induced by both H2O2 and heat shock (Supplementary Fig. 3; see
Phenotypes of trx1, txl1, and txc1 null mutants
Next, we constructed null mutants of txl1
+ and txc1
+. Both txl1Δ and txc1Δ mutants were viable and did not display any obvious phenotype, neither during normal growth conditions nor with physical or chemical insults such as high temperature, H2O2, DTT, caffeine, canavanine, cadmium, or sorbitol (not shown). Also, a txl1Δtxc1Δ double mutant appeared unaffected compared to wild-type cells (not shown). When protein degradation is strongly compromised, ubiquitin–protein conjugates are stabilized. We did not observe increased levels of ubiquitin-protein conjugates in txl1Δ or txc1Δ mutants, while stabilization was evident in the proteasome mutant mts3-1 strain (Supplementary Fig. 4; see
In S. pombe, Txl1 and Trx1 constitute the only thioredoxins in the cytosol. In order to determine whether Txl1 and Trx1 have an overlapping function in the maintenance of a reduced cytosolic environment, we generated trx1Δ and trx1Δtxl1Δ mutants and compared their growth under normal conditions or in the presence of H2O2. The trx1Δ mutant was highly sensitive to H2O2. However, the txl1Δ mutant was not sensitive to H2O2, and at low H2O2 concentrations the sensitivity of the trx1Δ mutant was only slightly increased upon loss of Txl1 (Fig. 3A). At higher H2O2 concentrations, the trx1Δtxl1Δ mutant was not more sensitive to H2O2 than the trx1Δ strain, indicating that Txl1 only targets a subset of oxidized cellular protein. Accordingly, trx1Δ cells exhibit cysteine auxotrophy, whereas txl1Δ and txc1Δ cells do not (Fig. 3B), indicating that Txl1 does not target 3’-phosphoadenosine-5’-phosphosulfate reductase involved in de novo cysteine synthesis (36).

Txl1Δ, but not txc1Δ, displays a synthetic growth defect with cut8Δ
High-throughput epistatic miniarray profiling (34) recently revealed that txl1Δ displays a synthetic growth defect in combination with a cut8Δ mutant (46). Since Cut8 has been reported to regulate the degradation of certain proteins by tethering the 26S proteasome to the nuclear membrane (39, 40), and since Txl1 physically interacts with the 26S proteasome, we decided to characterize the genetic interaction in more detail.
We first generated cut8Δ, txl1Δcut8Δ and txc1Δcut8Δ mutants and compared their growth. Growth of the txl1Δcut8Δ double mutant was reduced compared to wild-type cells and either of the single mutants (Fig. 4A). However, no synthetic effect was observed in the txc1Δcut8Δ double mutant (Fig. 4B), indicating that Txc1 is functionally distinct from Txl1.
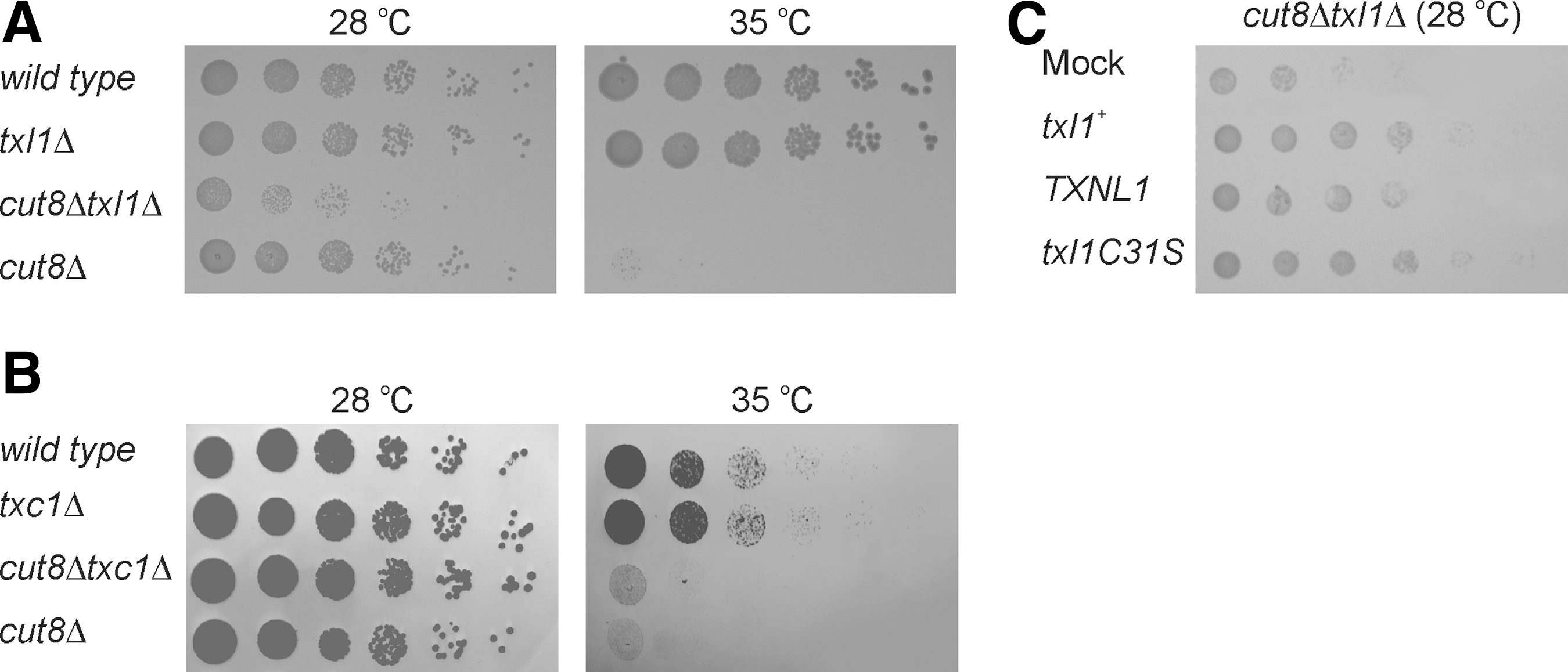
Overexpression of wild-type Txl1 or a point mutant in the active site Txl1C31S rescued the synthetic growth defect of the txl1Δcut8Δ double mutant (Fig. 4C), revealing that Txl1 enzymatic activity is not required for maintaining its Cut8 relevant function. Expression of human Txnl1 could also complement the growth defect of the double mutant (Fig. 4C), whereas Txc1 overexpression did not complement (not shown).
Moe1 is a component of the eIF3 complex, and proteasomes have been reported to mislocalize in moe1Δ mutants (49). We therefore also generated a txl1Δmoe1Δ double mutant. However, we did not observe any synthetic effects with this strain (not shown).
Txl1 co-localizes with 26S proteasomes
In order to further characterize the connection between Txl1, Txc1, and 26S proteasomes, we analyzed their subcellular localization using GFP-fusion proteins and fluorescence microscopy. Previous studies have shown that proteasomes are assembled in the cytosol in mammals (9), but in the nucleus in yeast (27). Fission yeast 26S proteasomes localize to the nuclear periphery in wild-type cells (37, 45) but are distributed throughout the cell in cut8 mutants (39, 40). We found that Txl1 co-localized with 26S proteasomes along the nuclear periphery in wild-type cells (Figs. 5A and 5B and Supplementary Fig. 5, see

We also analyzed the localization of 26S proteasomes in a txl1Δ strain. However, we did not observe any difference in this strain compared to the wild type (not shown).
Txl1 is an active thioredoxin
Since the main difference between Txl1 and Txc1 resides with the thioredoxin domain of Txl1, we decided to analyze Txl1 thioredoxin activity.
With insulin as a substrate (18), we found that Txl1, isolated from S. pombe cells (Fig. 6A),displayed thioredoxin activity, which was lost upon incubation with the alkylating agent, N-ethylmaleimide (NEM) (Fig. 6B). However, in comparison with thioredoxin from E. coli, Txl1 was significantly less active (Fig. 6B).
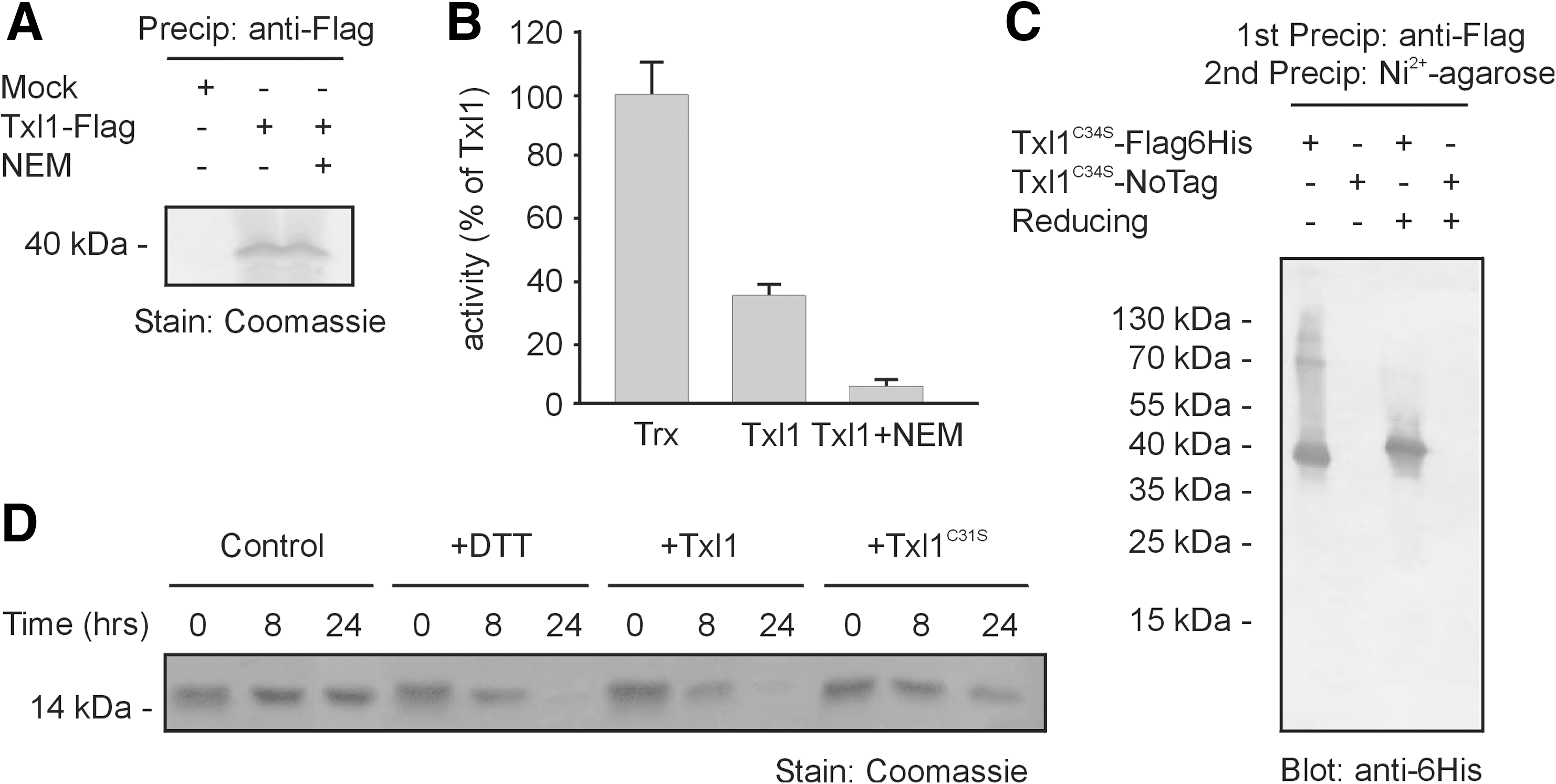
Having established that Txl1 is an active thioredoxin in vitro, we next expressed a Flag- and 6His-tagged Txl1C34S mutant in txl1Δ cells. Since Cys34 is the second cysteine in the active site of Txl1, changing this residue to a serine should still allow Txl1 to engage in disulfide exchange reactions. However, the mixed disulfide intermediates should be stabilized (1) so that substrates become covalently bound to the Txl1 enzyme. Precipitation of Txl1C34S, using a tandem affinity purification approach with first anti-Flag antibodies and then Ni2+-NTA agarose under denaturing conditions, revealed the presence of some slower migrating bands that were not observed upon treatment with reducing agent (Fig. 6C), suggesting that Txl1 is an active thioredoxin also in vivo.
Since several proteasome subunits, including Mts4/Rpn1 (50) and some 20S subunits (6), have been found to be glutathionylated, these might be deglutathionylated by Txl1. In addition, proteasome-associated deubiquitylating enzymes, such as Uch2/Uch37, are cysteine proteases and may rely on Txl1 for activity. However, we were unable to trap any Mts4/Rpn1, Uch2/Uch37, and 20S α subunits using the Txl1C34S mutant (Supplementary Fig. 6; see
Txl1 enhances degradation of a model proteasome substrate in vitro
A possible function of a proteasome associated thioredoxin, such as Txl1, includes reduction of oxidized proteasome substrates. Previous studies have shown that for efficient degradation, the proteasome requires that the substrate is sufficiently unfolded, and if disulfide linked, also that the disulfide bonds are reduced (44). We therefore investigated this hypothesis by analyzing the degradation of the tightly folded protein, α-lactalbumin, which contains several disulfide bonds. Mixing α-lactalbumin with purified 26S proteasomes did not result in any significant degradation. However, upon addition of the reducing agent DTT or wild-type Txl1, but not a Txl1C31S active site mutant, the substrate was degraded (Fig. 6D). Although α-lactalbumin is not ubiquitylated and an artificial proteasome substrate, the results indicate that at least under some conditions, proteasomes may rely on thioredoxins to catalyze the reduction of oxidized substrates prior to degradation.
Discussion
Txl1 and Txc1 are largely uncharacterized proteins that are conserved between fission yeast and higher eukaryotes, but are not present in S. cerevisiae. We show that both proteins contain PITH domains and physically associate with the 26S proteasome. We therefore conclude that the PITH domain is probably a general proteasome-interacting domain. In the case of Txl1, we also observed a co-localization with the 26S proteasome, indicating that most or all of Txl1 is associated with 26S proteasomes in vivo. In contrast, Txc1 did not co-localize with 26S proteasomes, and we therefore speculate that Txc1 only transiently associates with 26S proteasomes in vivo, and that a significant fraction of Txc1 is not associated with 26S proteasomes in wild-type cells.
The structures of the human and plant Txl1 homologues Txnl1 and At3g04780.1-des15, respectively, have been solved (12, 35). The structure of the PITH domain is essentially a jelly roll β-sandwich, with a two-stranded β-sheet sealing off one end forming a cavity. A conserved cysteine residue is exposed in this cavity and may play a role in proteasome binding (12). Interestingly, the structure of the PITH domain closely resembles budding yeast Doc1/Apc10, a subunit of the anaphase-promoting complex or cyclosome (APC/C) (35). The APC/C is a multi-subunit E3 ubiquitin-protein ligase that regulates cell cycle progression (33). However, no specific function of the Doc1/Apc10 subunit has yet been described, and the relevance of the structural similarity to PITH domains therefore remains unknown.
Previous studies on Txl1 in higher eukaryotes have linked Txl1 function to endocytosis (8) and protection against glucose deprivation-induced cytotoxicity (21). In S. pombe, Txl1 is not involved in response to glucose deprivation (22), and we suggest that the observed effects on endocytosis and glucose-deprivation protection are indirect.
In C. elegans, Txl1 expression is induced by the unfolded protein response (41), and in mammals Txl1 expression is increased in response to proteasome inhibition (1, 29). In S. pombe, Txl1 expression has been reported to be slightly upregulated by oxidative stress via the stress activated transcription factor Pap1 (24). We did not observe any stress responsive expression of txl1 + and txc1 +. However, in agreement with published microarray data (5), we did observe a significant induction of trx1 + in response to oxidative and heat stress.
Both during aging and in general, proteasomes are prone to oxidation (10, 14, 20, 47, 48), and oxidation as well as glutathionylation of the 26S proteasome affects the proteolytic activity (6, 20). In this respect, it is not surprising that under oxidative stress conditions, the proteasome would need an enzymatic system to maintain the particle in a reduced form. Accordingly, 26S proteasomes have been reported to remodel scrambled RNase A (28), indicating that proteasomes are associated with thioredoxin activity, that we suggest can be ascribed to bound Txl1. We probed our substrate capture precipitates for the presence of Mts4/Rpn1 and 20S α subunits that have been reported to be glutathionylated (6, 50). However, none of these components were isolated as Txl1 targets.
We show that Txl1 is an active thioredoxin, but unlike the other cytoplasmic thioredoxin, Trx1, null mutants in txl1 are not overly sensitive to oxidative stress and are not auxotroph for cysteine. These data argue that the thioredoxin activity of Txl1 plays a more specialized role and probably targets a more narrow selection of substrates than Trx1.
A null mutant in txl1 (46), but not in txc1, displays a synthetic growth defect with mutants in cut8. However, a Txl1 active site point mutant can complement the genetic interaction. Since Cut8 tethers proteasomes to the nuclear rim, proteasomes become mislocalized in cut8 mutants (39, 40). However, we did not observe any synthetic growth defects in a txl1Δmoe1Δ strain and proteasomes localize normally in a txl1 null mutant. We therefore suggest that the observed genetic interaction with cut8 is not directly connected with the function of Cut8 in proteasome localization.
In conclusion, we show that Txl1 and Txc1 interact with the 26S proteasome. Txl1 equips the particle with protein disulfide reduction capabilities that for some substrates may be required for efficient degradation. However, despite our efforts, the exact molecular mechanism for Txl1 and Txc1 function remains elusive.
Footnotes
Acknowledgments
We thank Dr J.H. Roe for sharing valuable reagents and Dr C.R. Wilkinson and Dr L. Ellgaard for helpful discussions. This work has been supported financially by grants from the Lundbeck Foundation, the Novo Nordisk Foundation, and the Danish Natural Science Research Council to RHP and from the Medical Research Council to CG.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.