
Abstract
Tissue regeneration and several diseases such as tumor and atherosclerosis depend on new vessel formation by both angiogenesis and vasculogenesis. Endothelial cells (ECs) are widely considered to be the active cellular component in these processes, followed by contractile cells such as pericytes and smooth muscle cells. The best known sources providing these cell types or their progenitors are ECs lining the vessel lumen and bone marrow. As easily evident, the vessel wall was recognized as being a passive player to a great extent except ECs of the vascular intima. Particularly, the vascular adventitia has been considered as a passive layer rather than an active part of the vessel wall. But results provided during the last few years have led to a revision of this classical view because of an apparent stem cell niche function of the vascular adventitia. This review aims to sum up findings identifying the vessel wall as an important stem cell reservoir and discusses its impact on health and disease. Antioxid. Redox Signal. 15, 981–995.
Introduction
Tissue regeneration not only requires stem or progenitor cells but also functioning blood vessels, which are capable to ensure a sufficient blood supply by connection to the preexisting blood circulation. Moreover, the creation of appropriately functioning blood vessels is also crucial for proper internal tissue structure and stroma formation. In embryogenesis, ECs are generated from hemangioblasts, which in turn are derivatives of the mesodermal stem cells and also serve as progenitors for hematopoietic cells. This de novo development of blood vessels from embryonic stem cells is called vasculogenesis and is responsible for the construction of aorta and its main branches. Several physiological and pathological processes in the adult such as endometrial cycles, tissue regeneration, ischemia, wound healing, atherosclerosis, and tumor growth require the formation of new blood vessels (17, 37, 56). Despite the existing inconsistencies, it is generally believed that at least a part of these new vessels in the adult is formed by stem and progenitor cells, which are capable of differentiating into all types of the vascular wall (VW). A problem vascular researchers face today is how the preexisting mature ECs orchestrate with EPCs coming from different inches during new vessel formation and, particularly important, to which extent the organ- and tissue-specific local progenitors are involved in these processes. This review aims to provide an overview about the VW as a niche for different stem and progenitor cell types and to discuss their potential impact on new vessel formation under physiological and pathological conditions.
Basics of the Vessel Wall Structure and VW-Resident Stem Cell Niche
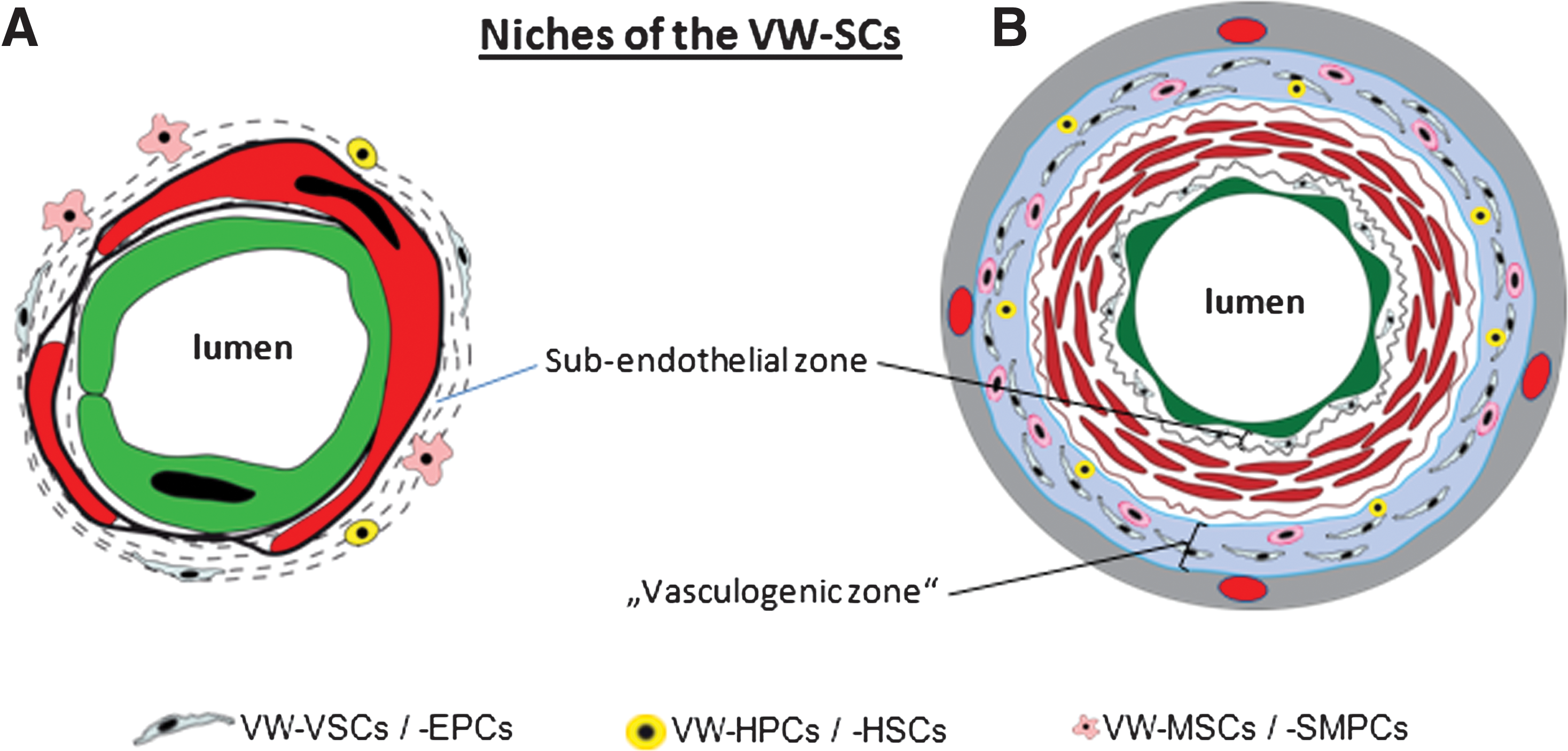
A brief overview of the basic structure of blood vessels is necessary to exactly define the location of the VW niche for stem or progenitor cells and to characterize their role in physiologic as well as pathologic processes. The smallest vessels, namely the capillaries, are built by two cellular layers, EC lining the lumen and pericytes, a noncontinuous contractile cellular component of the capillary wall. Both ECs and pericytes are in tight contact with each other and are embedded in the same basal lamina (Fig. 1A). Also, arterioles and venules belong to microvessels but, unlike the capillaries, their wall contains at least one layer of smooth muscle cells (SMCs) instead of pericytes. All microvessels are covered by a thin connective tissue layer and are integrated via this into the perivascular tissue. The wall of large vessels such as arteries and veins is principally organized by three concentric and clearly distinguishable layers: the intima, media, and adventitia (Fig. 1B). The innermost layer, called tunica intima, like in capillaries, is build by ECs. The media is constructed mainly by SMCs, and finally, the adventitia is formed by connective tissue cells. Although the wall of capillaries, small arteries, and veins is supplied with oxygen and nutrition via convection from the circulating blood and thus does not need vasa vasorum, the outer parts of the wall of larger arteries and veins is additionally supplied via vasa vasorum located within the vascular adventitia. Under normal conditions, all components of the vessel wall are mostly quiescent, but in several physiological and pathological processes with new vessel formation via angiogenesis, EC proliferation and migration have been shown (16, 32, 35, 38). Also, accumulation of α-smooth muscle actin (α-SMA)–positive cells during formation of the neointima has been described, but it remains controversial whether these cells are derived from the preexisting mature SMCs of the arterial wall or from precursor cells as recently reviewed by Orlandi and Bennett (78). Until few years ago, the adventitia was not considered to play an essential role in angiogenesis and postnatal vasculogenesis. But experimental evidence during the last few years have identified this part of the wall of fetal and adult arteries as well as veins and the wall of capillaries as a niche for various types of stem and progenitor cells (VW-resident stem cells [VW-SCs]) including EPCs, progenitors for SMCs, mesenchymal stem cells (MSCs), HSCs, the so-called mesangial cells coexpressing both endothelial and myogenic markers, neural stem cells, and germline stem cells. Further, malignant stem cells as shown for leukemic and glial malignancies are also found to reside in this niche of the vessel wall (13, 25, 27, 62, 100). These findings together suggest that the adventitial niche harbors not only progenitors or multipotent stem cells contributing to new vessel formation or renewal of vessel wall cells, but also tissue regeneration and regrowth of tumors because of the reported tight relation of the so-called cancer stem cells to the VW niche. As evident from this brief summary, a broad spectrum of stem and progenitor cell types resides in the wall of arteries and veins as well as microvessels. The fact that blood vessels are present in the smallest corner of the human body illustrates why it is necessary to understand the role of VW-SCs in health and disease.

VW-SCs in microvessels
Microvessels compromise arterioles, capillaries, and venules. They share the structural similarity to be constructed by ECs and only one layer of peri-ECs, namely by pericytes for capillaries and one layer of SMCs for arterioles and venules. Pericytes cover the ECs of nonmuscular microvessels. Electron microscopic analyses reveal one of the structural characteristics of pericytes, that is, they are embedded in the same basal lamina as ECs as visualized in Figure 1A. The close relation between ECs and mature pericytes is documented by the presence of gap and tight junctions between both cell types. Mature pericytes are contractile and can regulate the diameter of and thus the blood perfusion through the microvessels. As pericytes do not cover the entire capillary wall, the pericyte-free parts of capillary wall represent the thinnest sections of microvessels where exchange of gas and nutrition mainly takes place. Although the nature of pericytes remained elusive for a long time and remains not fully understood, recent publications suggested that they might function as MSCs once they are detached from the endothelial layer. Based mainly on flow cytometric characterization of CD34+CD31−CD144− cells isolated from the wall of microvessels, it has been reported in several publications that these cells coexpress both pericyte and MSC markers such as CD140 a/b, CD146, NG2 and CD10, CD13, and CD90 (24, 108). Crisan et al. showed that the so-called “perivascular cells” natively express the MSC markers CD44, CD90, CD73, and CD105 (24). As discussed below, the term perivascular superficially implies localization outside and around the VW, but it should be considered that the cells mentioned here are integral structures of vessel wall, thus also called mural cells. Frequently, the adipose tissue-derived stem cells (ADSCs), which were found to express MSC markers, have been suggested to be generated from pericytes, but recent data questioned this concept and postulate rather the existence of VW-resident MSCs (VW-MSCs or VW-SCs) in colocalization with the mature pericytes of small vessels in adipose tissue (71, 117). Using double immunostaining with subsequent confocal microscopic analyses, the authors could show that in the native tissue, the ADSCs, which are mainly localized in vessel wall and exhibit MSC properties, do not coexpress CD34 and CD140b or CD34 and α-SMA. In a further recent publication, it was reported that CD34+ cells isolated from the vessel fraction of adipose tissue exhibit MSCs but not pericytic markers (102). Taken together, these findings suggest the presence of VW-SCs in the wall of microvessels in addition and in close vicinity to pericytes (Fig. 2A).
Based on the location of pericytes around the EC layer of small vessels and their postulated capability to differentiate to MSCs, a subendothelial niche for MSCs has been suggested (91). However, in some other publication, the location of pericytes has been described to be perivascular. Unfortunately, the term perivascular is mistakenly used in several high-ranked publications for the progenitor and stem cells, which actually are not located perivascular but within the VW. Taking into account that the vessel wall is not only constructed by ECs and the pericytes are not outside the capillary wall but rather an integral part of it as described above, it is clear that the term perivascular for VW-SCs causes confusion. Thus, we will not use this term in the following and emphatically suggest the term “VW-resident” instead of “perivascular.” A more recent publication demonstrated that not the classical pericytes serve as a reservoir for MSCs but rather a different subpopulation of cells located in the capillary wall in between the pericytes exhibit cell surface markers characteristically for MSCs. This cell bears the potential of three-lineage differentiation as shown for MSCs, whereas the mature pericytes isolated from the capillary wall showed neither the MSC markers nor the three-lineage differentiation (70). These data suggest that the origin and nature of pericytes need to be characterized. One conclusive explanation might be that they differentiate from the VW-MSCs localized in close vicinity to the mature pericytes (Fig. 2A). But also BM-derived progenitor cells can be guided to the periendothelial area of the vessel wall, where they might differentiate to pericytes.
VW-SCs in medium-sized and large blood vessels
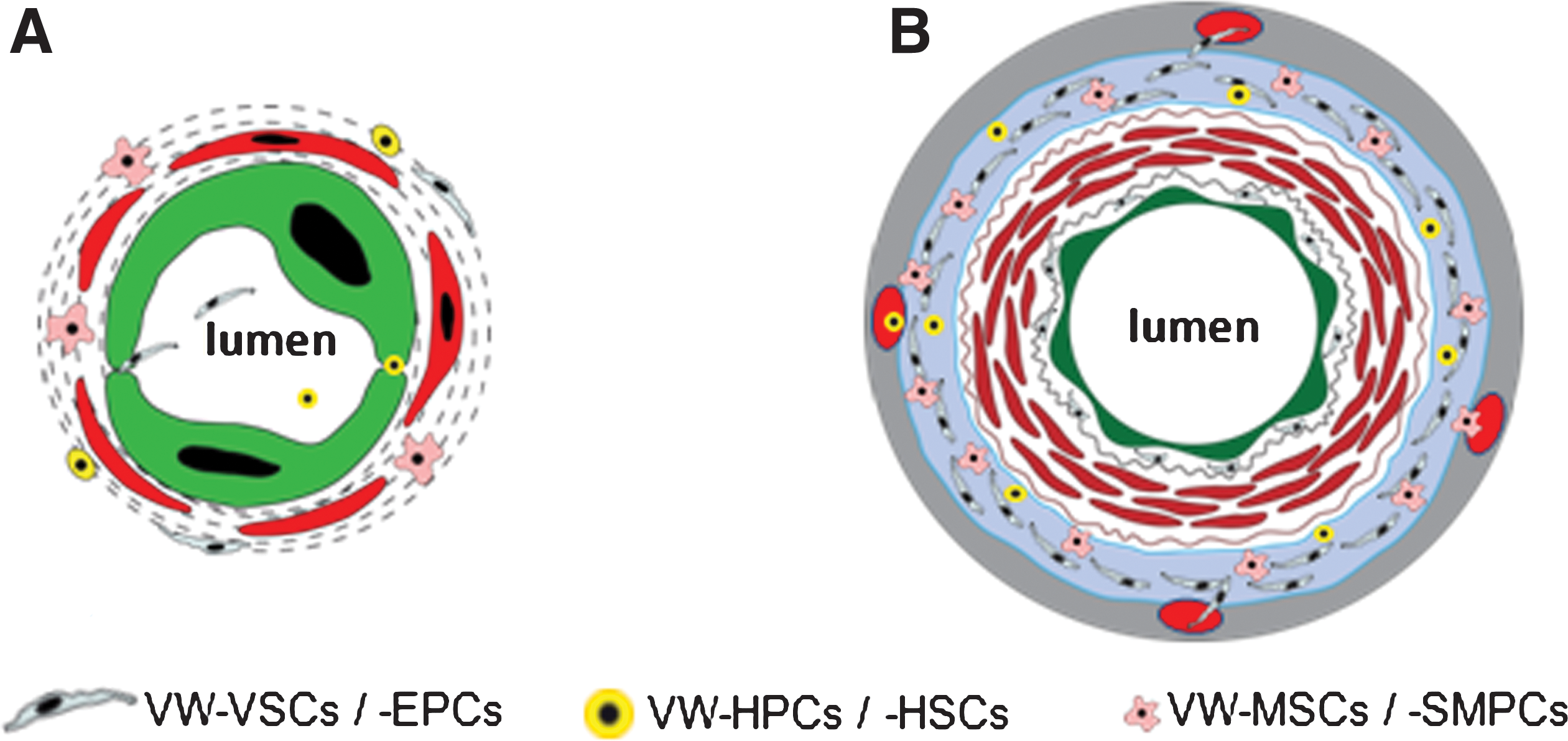
Most publications about stem and progenitor cells in the vessel wall report mainly two zones in the wall of arteries and veins harboring these cells: (i) the subendothelial zone and (ii) the vascular adventitia, particularly the so-called vasculogenic zone within the vascular adventitia close to the vascular media (2, 31, 33, 53, 64, 79, 80, 120) (Fig. 2). The subendothelial or the so-called “periendothelial” zone of microvessel wall corresponds more or less to the adventitial zone of large and medium-sized blood vessels. In contrast to the microvessels, which principally possess one layer of contractile cells (pericytes or SMCs) in their wall, the wall of medium-sized and large blood vessels comprises all three layers mentioned above irrespective of their intra- or extraorgan localization. In these vessels, CD44+CD90+CD73+CD34−VEGFR-2− MSCs and VEGFR-2+CD34+CD31− EPCs as well as CD133+CD45+ hematopoietic progenitor cells (HPCs) were mainly found in the vasculogenic zone within the vascular adventitia (33, 79, 105, 120) (Fig. 2B). Although the characterization of different subtypes of VW-SCs in their native niche remains unsatisfying and the flowcytometric determination of isolated cells is not conclusive for the in situ situation, the combination of both immunostaining on sections from different human vessels and flowcytometric analyses of cells isolated from human vessel fragments confirmed the existence of different subsets of stem and progenitor cells in the vessel wall.
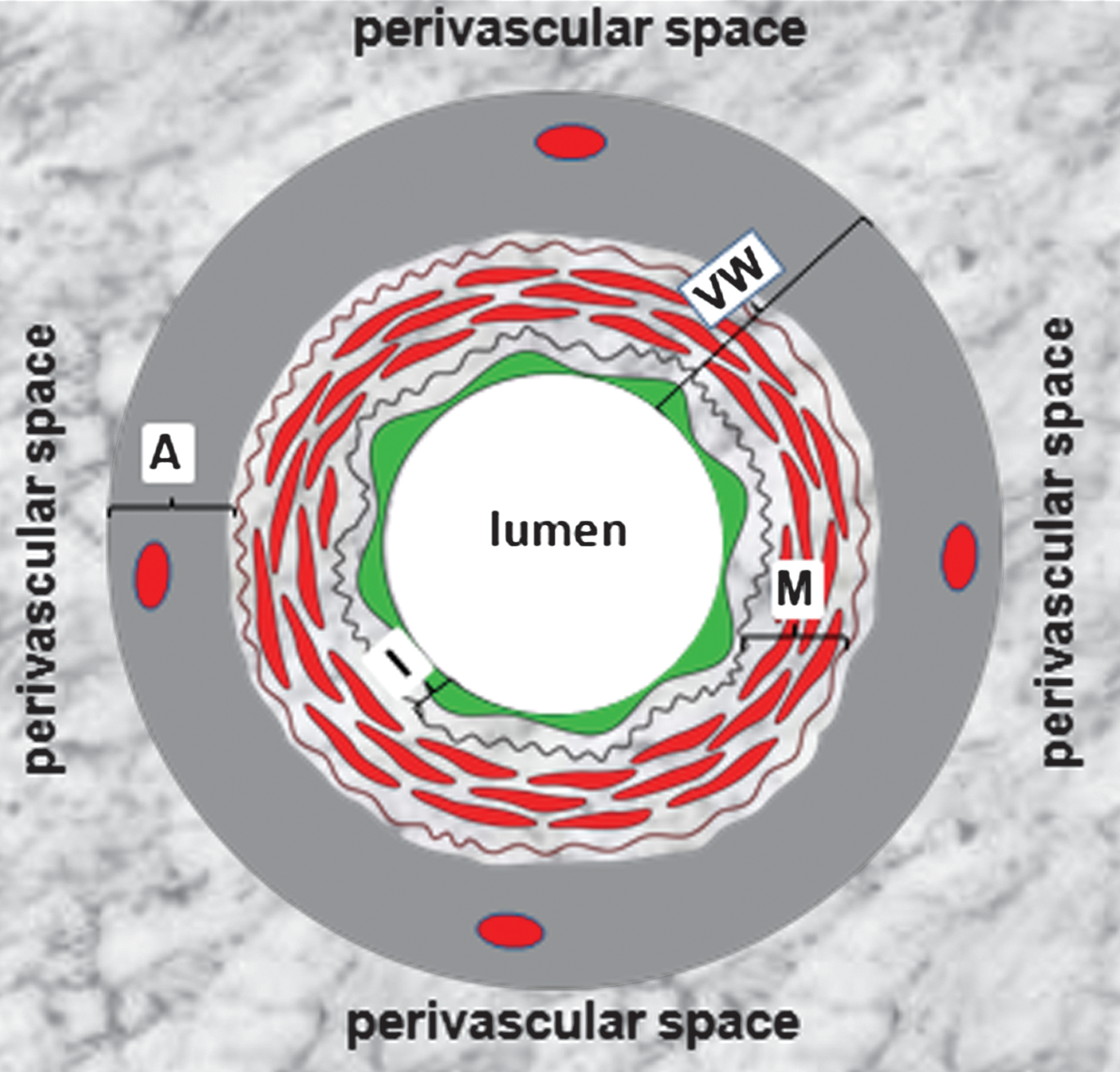
Data in the recent literature put the vascular adventitia into the focus of research of VW-derived stem and progenitor cells. These reports confirmed the existence of both multipotent stem cells such as MSCs and HSCs and unipotent lineage-committed progenitors, for example, EPCs within the vascular adventitia (2, 33, 53, 79, 105, 107, 108, 120). Basically, the vascular adventitia is constructed by fibroblasts and fibrocytes and contains a vessel network called vasa vasorum. Two fundamental functions can be attributed to this vessel wall layer: (i) it serves as an interface between the inner part of vessel wall and the perivascular tissue. The clinical relevance of this interface role of vascular adventitia is not only supported by the aforementioned findings about stem and progenitor cells but also further underlined by the recent reports showing an intensive remodeling of pulmonary artery adventitia by hypoxia and adventitial inflammatory reaction already in the initial stage of atherosclerosis (12, 26, 39). On the other hand, a recent publication demonstrated that white adipose tissue of the perivascular space, that is, outside or beyond the vascular adventitia, induces proliferative and inflammatory processes within the vascular adventitia (20). Based on such observations, an “outside-in” component of the vessel wall to atherosclerosis in addition to the conventional “inside-out” view has been postulated (89). This interaction between the vascular adventitia and the perivascular white adipose tissue again emphasizes how important it is to use the terms “VW-resident” and “perivascular” correctly to avoid confusion and to distinguish different tissue compartments. This is not only necessary because of academic correctness but also because of its relevance for potential therapeutic use of cells located in these tissue compartments. As discussed above for capillaries, and also for large vessels, the term perivascular has been used for stem and progenitor cells, which actually are localized within the vascular adventitia and thus a part of vessel wall. The correct structural relation between VW and perivascular space is shown in Figure 3. Further aspects of the adventitial interface role will be discussed later in this review. (ii) The vascular adventitia contains vasa vasorum network, which historically has been considered to provide an essential conduit supplying the outer parts of the vessel wall with oxygen and nutrition. In addition, accumulating data in the literature suggest that vasa vasorum plays a significant role in maintaining vessel integrity but also may contribute to development of vascular disease (8, 49). Further, it enables the transmigration of cells circulating in the peripheral blood, for example, BM-derived stem and progenitor cells into the vascular adventitia, as it has been demonstrated for mononuclear cells (MNCs) in pulmonary artery adventitia under hypoxic conditions (39). In brief, these data suggest that vasa vasorum might also facilitate the transmigration of BM-derived stem and progenitor cells to the vascular adventitia and thus contribute to building and maintenance of the VW-resident stem cell pool. Alternatively, the VW-SCs might have been retained there since embryonic development of blood vessels.
Developmental origin of VW-SCs
In 1988, before we were able to detect VW-SCs by modern techniques, Simionescu and Simionescu set forth the principle that the VW harbors a pluripotential pool for cell formation and local repair, for example, endothelial regeneration (99). The question is whether VW-SCs reside within the VW-resident niches only temporarily or whether they were positioned at these sites during the embryonic development of blood vessels and remained there the whole life entire. The de novo blood vessel development by vasculogenesis is provided by embryonic mesodermal progenitors, which aggregate to small clusters called blood islands in intra- and extraembryonic tissues including yolk sack (44). By action of several factors including fibroblast growth factor-2 and VEGF, the outermost cell row of these islands become endothelial precursors termed angioblasts, whereas the cells of the central part of the islands differentiate into hematopoietic precursor cells (87). Because of this close similarity between angioblasts and HPCs, the existence of a common ancestor precursor cell called hemangioblast has been postulated several decades ago. Although this speculation has faced controversial debates for a long time, recent studies provide strong evidence that indeed such hemangioblast-like cells exist, which give rise to both EPCs and HPCs (21, 51, 112). Interestingly, also within the VW-resident niche called as vasculogenic zone, CD34+CD31−VEGFR-2+EPCs and CD45+ HPCs reside in close proximity (79, 120), whereas a VW-resident hemangioblast-like cell has not been identified yet. One explanation for the origin of the VW-SCs is that they are representing a part of BM-derived stem cells and progenitors that reach the subendothelial space of microvessels by transmigration through the capillary wall (Fig. 4A) and the vascular adventitia via transmigration through the wall of vasa vasorum and finally home to the vasculogenic zone (Fig. 4B). Although transmigration of BM-derived cells to both vascular adventitia and the perivascular space has been reported (26, 39, 48) under different conditions, the aforementioned mechanism is hypothetical. Interestingly, a transmigration of BM-derived MNCs into the pulmonary artery adventitia of mice with hypoxia-induced pulmonary hypertension has been reported (39), and this process was not observed in the vascular adventitia of control mice. The extravasation of BM-derived cells including stem and progenitor cells into the adventitia of pulmonary artery under hypoxia was accompanied by a significant increase of density of vasa vasorum, suggesting that newly formed vasa supports an increased transmigration of circulating cells (26). Again in control mice without hypoxia, this phenomenon was not observed, indicating that under normal conditions the BM-derived stem and progenitor cells are not transmigrating to the vascular adventitia in considerable amounts and thus are probably not the source of VW-SCs. On the other hand, it is conceivable to postulate that VW-SCs are probably generated from embryonic stem cells, which were initially located at the periendothelial space of embryonic blood vessels including aorta and its main branches (Fig. 5). At the initial stage and during the first week of cardiovascular development, even the wall of embryonic aorta is built of only ECs (86, 87) while stem and progenitor cells are still present around the endothelium (Fig. 5A). The deposition of extracellular matrix components including laminin and subsequently accumulation of progenitor cells in the periendothelial space give rise to pericytes and SMCs around the endothelial tube (Fig. 5B, C). But a part of these stem and progenitor cells conserve their stem cell properties during further development and remodeling of the vessel wall until adulthood and probably build the vasculogenic zone within the vascular adventitia (Fig. 5D). This hypothetical concept has not been taken into consideration until recently because of main focus of stem cell research on BM-derived cells. In a very recent publication, it has been suggested that the physical association of multilineage progenitors to the vessel wall may probably be established during development as shown for lateral dorsal aorta and anterior mensenteric arteries in a zebra fish model (93). Further analyses are needed to clarify the origin of VW-SCs. This issue is of functional and clinical relevance to characterize to which extent VW-SCs interact with BM-derived progenitors during organogenesis, adult tissue repair, and development of diseases such as tumor growth and vascularization. If VW-SCs are not derivatives of BM-derived stem and progenitor cells as postulated above, then they might have a different behavior than these cells, which might be of relevance for therapeutic use of the VW-SCs.

EPCs in Adult Vessel Wall
An increasing number of publications demonstrate in ex vivo and in vivo experimental models that the adult VW harbors different types of stem and progenitor cells types, which in summary are called VW-SCs (2, 33, 53, 66, 79, 105, 120). Subtypes of these cells are capable to differentiate to mature VW cells such as ECs, SMCs, and fibrocytes. EPCs are the most intensively studied vascular progenitor cell type. EPCs are defined as cells that show clonal expression, stemness characteristics, adherence to matrix molecules, and the capacity to differentiate into mature ECs (7). Several markers are considered to be characteristic of EPCs such as CD34, AC133, VEGFR-2, Tie-2, and the ligand for Ulex europaeus agglutinin-1 lectin as well as acetylated low density lipoprotein uptake (5, 7). The use of subsets of antibodies against different markers in flow cytometry and also in immunocytochemical and immunohistochemical analyses is necessary for the identification of EPCs, because to this point no single marker can discriminate circulating ECs and EPCs from the hematopoietic cells, particularly from the HSCs. The main sources for EPCs have been considered for long to be BM-EPCs and peripheral and cord blood (circulating EPCs [C-EPCs]) (4, 74, 77), but in the last few years a growing body of literature has identified the wall of adult blood vessels as an important niche for EPC, the so-called VW-resident EPCs (VW-EPCs) (1, 2, 47, 52, 53, 104, 120). In a considerable number of studies, peripheral blood cells expressing CD34, VEGFR-2, and CD133 have been recognized to represent C-EPCs, but, as mentioned above, recently published studies have questioned this concept and suggest that these cells are hematopoietic cells rather than EPCs (106, 116). Despite these discrepancies, the main body of literature supports the concept that BM-EPCs contribute to tumor vascularization as shown in the Id-mutant mouse model where tumor progression was inhibited because of the absence of mobilization of BM-derived progenitors and defective angiogenesis (40). The existing controversy regarding the contribution of BM-EPCs to new vessel formation makes apparent that new, more profound in vivo models are needed to clarify this point and to distinguish BM- and C-EPCs from HSCs and HPCs. As mentioned above, the identification of VW-EPCs proves the existence of EPC niches outside the BM. Immunostaining analyses identified a particularly high number of CD34(+)VEGFR-2(+)Tie-2(+)CD31(−)VE-cadherin(−)-negative VW-EPCs in a distinct zone within the adventitia of large and medium-sized blood vessels, the so-called vasculogenic zone (Fig. 2B) (120). The outgrowing VW-EPCs forming capillary-like tubes become positive for CD144 (2, 79, 120). Another important location for VW-SCs, which have been reported to have the potential to differentiate into EPCs, is the subendothelial zone (92, 108). Further, the EPCs isolated from adipose tissue have been also shown to be mainly present in the microvessel fraction (47) and to be located in sub- or periendothelial space close to the pericytes. Although it is unknown until now whether VW-EPCs are mobilized into the peripheral blood, it has been demonstrated that EPCs derived from peripheral organs such as liver or intestine contribute to the pool of C-EPCs (1). Since arteries and veins of these organs contain VW-EPCs, one can speculate that at least a subpopulation of EPCs coming from these organs might be mobilized from the blood vessels and may contribute to the pool of C-EPCs. Further, VW-EPCs located in the periendothelial space of vasa vasorum can be also easily mobilized into the blood circulation. The mechanisms underlying the mobilization and egress of VW-EPCs into the systemic blood circulation is as of yet unknown but of clinical relevance.
Besides the aforementioned endogenous processes, VW-EPCs have a great potential to be used in cell-based clinical therapies because fragments of adult arteries and veins can be surgically extracted from different anatomic regions and used for isolation of VW-EPCs, which can be subsequently expanded and prepared for clinical use. However, prior to the use of VW-EPCs in the clinic, several concerns should be addressed, such as ageing status, angiogenic activity, and the capacity of EPCs to integrate into existing tissue structure. Additionally, it is necessary to identify the molecular mechanisms governing the differentiation of VW-EPCs to mature ECs ex vivo. Further, another unanswered question is whether the VW harbors more ancestor cells, which differentiate into VW-EPCs and other types of lineage-committed progenitors, for example, smooth muscle. It is necessary to identify the molecular mechanisms governing the differentiation of multipotent VW-SCs into EPCs and smooth muscle progenitor cells as it was shown for HSCs by analyzing the expression of signaling lymphocyte activation molecule family receptor members (62). Finally, it remains unexplored whether and how the VW-EPCs react in different diseases such as atherosclerosis, tumor development and metastasis, local inflammatory processes within and outside the vessel wall, and finally, wound healing and tissue regeneration. Better knowledge of the molecular and cellular mechanisms that regulate the endogenous mobilization of VW-EPCs in such pathological processes is essential to design new drugs for therapeutic manipulation of VW-EPCs, for example, either suppressing or activating their mobilization, in different disorders. Also, potential interactions between VW-EPCs and other types of progenitors or stem cells resident in the vasculogenic zone of vessel wall, such as VW-MSCs and -HPCs, need to be better characterized.
HSCs in Adult Vessel Wall
The generation of HSCs in the vertebrate embryo has been a matter of controversy for several decades. Besides the “hemangioblast hypothesis,” whereby both ECs and hematopoietic cells are derived from a common maternal cell located in blood islands within the yolk sac wall, recent studies strongly suggest a hemogenic endothelium that transiently possesses the capability to give rise to HSCs during vertebrate development (11, 30, 76, 103, 121). In vivo studies on zebra fish and permanent in vitro tracing analyses enabled the observation of a stepwise transition from the intermediate hemogenic endothelium of aorta to nascent HSCs (11, 30). The close developmental relation between the emerging of ECs and HSCs is further supported by recent in vivo studies using mouse models and the blast colony-forming cells generated from embryonic stem cells (10, 30, 51, 97, 110), demonstrating indeed the existence of the “hemangioblast” as the ancestor cell for both HSCs and EPCs. Remarkably, it could be shown that brachyury (also known as T) and Flk-1–positive hemangioblasts are frequently present in the posterior region of the primitive streak before blood island development in the yolk sac (51). These data support the hypothesis that both ECs and hematopoietic cells are likely, at least partially during the embryonic development, derived from a common ancestral stem cell, which can be available in form of the hemangioblasts in the so-called blood islands and/or transiently in form of the hemogenic endothelium in the aorta. But it is to mention that these data were not confirmed by other studies. Lineage-tracing experiments, in which mouse embryonic stem cells stably expressing separate fluorescent proteins were implanted into blastocysts, predominantly resulted in multiple progenitors; however, evidence of one progenitor giving rise to both ECs and hematopoietic cells was an infrequent event (109). Besides these discrepancies, there is no doubt about the close proximity of EPCs and HSCs/HPCs in niches identified so far irrespectively of embryologic and adult period. Accordingly, results obtained from studies on VW-SCs suggest a coexistence of EPCs and HSCs within the vascular adventitia (79, 120). Interestingly, Peault and coworkers demonstrated that during embryogenesis, a population of VEGFR-2(+) angiohematopoietic mesodermal stem cells migrates from the para-aortic splanchnopleura into the ventral part of aorta, where they give rise to both ECs and hematopoietic cells (104). A further hypothesis is that HSCs are not generated from the aortic endothelium, but rather may emerge from the subendothelial mesenchyme surrounding the aortic endothelium (63). These findings might help to explain the fact that CD45(+)CD34(−) HSCs and CD34(+)Tie-2(+)VEGFR-2(+)VE-cadherin(−) EPCs reside alongside within the vasculogenic zone of the vessel wall (120) as shown in Figure 2. This symbiotic presence of both HSCs and EPCs in the VW as in adult BM or in the yolk sack during embryogenesis (104) may be due to the aforementioned subendothelial mesodermal/mensenchymal stem cells, of which a part remains in the vascular adventitia, retaining stem cell properties until adult. VW-HSCs are apparently capable to differentiate into mature blood cells as shown for CD68(+) macrophages in an in vivo experimental model by depletion of the BM (Fig. 6) (120). Further, primitive hematopoietic cells and EPCs share the expression of a great part of the cell surface markers such as c-kit, CD133, Sca-1, VE-cadherin, VEGFR-2, and endoglin (18, 65, 84). Functionally, the concerted action of both HSCs and EPCs seems to be essential for tumor vascularization. VEGF, secreted by tumor cells, recruits VEGFR-1(+) HSCs and VEGFR-2(+) EPCs to the tumor tissue and activates neoangiogenesis (59).

Collectively, recent data strongly suggest the existence of both the hemangioblast giving rise to ECs and hematopoietic cells as well as the transiently emerging hemogenic endothelium in aorta, which gives rise to hematopoietic cells in the embryonic period. But apparently also subendothelial mesodermal/MSCs in the wall of embryonic aorta possess the capability to generate hemogenic endothelium and thus HSCs. Based on the findings in the last few years about VW-SCs, it is conceivable to postulate the existence of VW-resident adult hemangioblast and hemogenic ECs, although they have not been identified yet. Despite these hurdles, the convincingly demonstrated close proximity of EPCs and HSCs in the wall of adult blood vessels implies functional interdependences and potential transdifferentiation processes between VW-EPCs and VW-HSCs. The characterization of these would open new therapeutic avenues in postnatal vasculogenesis, cardiovascular repair, and tissue regeneration.
Ancestors of Multipotent Stem Cells in Adult Vessel Wall
Probably one of the most complex and difficult issues in the field of stem cells is the identification and characterization of adult multilineage stem cells known under different terms such as MSCs (82), multipotent adult progenitor cells (57), and muscle-derived stem cells and/or ADSCs (81). The great majority of these cells have been characterized under culture condition after their isolation from the tissues mentioned, but no or only little information is available about their native identity, frequency, and exact anatomical location. In terms of ancestors of multipotent stem cells resident in the adult human vessel wall, we probably have an unique situation because studies during the last few years provided a great progress by identifying the exact niche within the adult vessel wall, where different types of stem and progenitor cells including ancestors of multipotent stem cells reside as also summarized above. One of the frequently identified multipotent stem cell types among the VW-SCs are VW-resident MSCs (VW-MSCs) (14, 22, 91, 107, 108, 118, 120) (Fig. 2). Generally, MSCs are thought to be multipotent non-HSCs. In the BM, the most frequently reported niche for MSCs, they represent only a small percentage of the marrow cells (15). Although none of the following markers are specific for MSCs, the most frequently used markers are CD105, CD44, CD90, and Stro-1. MSCs are negative for CD34 and CD14 (19). Under specific culture conditions, the BM-derived MSCs differentiate into fat, muscle, bone, and cartilage cells (50, 101). This so-called three-lineage differentiation potential is one of the accepted basic criteria for MSCs. BM-derived MSCs also differentiate into vascular SMCs (60). Once seeded on a synthetic vascular graft, MSCs differentiate into both SMCs and ECs in vivo (72).
Besides BM, multipotent stem cells including MSCs have been extracted from multiple organs such as dental pulp, placenta, fat, bulge region of hair follicles, and umbilical cord. In the wall of adult blood vessels, first pericyte-like cells have been thought to serve as a cellular reservoir for VW-MSCs (91). According to this hypothesis, the pericyte-like cells are present in the subendothelial space of all vessels. Although several studies have demonstrated similarities between peri-ECs of capillaries and MSCs (23, 88, 94, 96, 119), neither stringent cellular characterization, for example, in situ double immunostainings using different markers, nor exact cell-sorting analyses were performed yet to exactly determine the relationship between pericytes and VW-MSCs. It is conceivable that the periendothelial VW-MSCs in microvessels are a discrete cell type of capillary wall rather than pericytes (Fig. 2).
In medium-sized and large blood vessels, the vascular adventitia is identified to contain VW-MSCs (14, 107, 120). First, it could be shown that a small fraction of cells mobilized from the adventitia of human adult artery during ring assays express α-SMA and show a close association to endothelial tubes formed by VW-EPCs (120). More recently, a subset of progenitor cells positive for CD34 but negative for endothelial markers such as CD31 and von Willebrand factor was isolated from the adventitial layer of human adult vena saphena wall and was shown to express MSC markers as CD44, CD90, CD105, CD49a and b, CD59, and CD73 under culture conditions (14). These cells differentiated into pericytes and SMCs and cover new endothelial blood vessels. Supporting these findings, in an elegant genetic mouse model, Passman et al. demonstrated the existence of Sca1(+) vascular stem cells in the arterial adventitia capable of differentiating into SMCs (80). Of note, these Sca1(+) vascular stem cells as well as the aforementioned CD34+ ancestors to MSCs are localized precisely to the vasculogenic zone, which was previously shown to harbor EPCs and HSCs (120). In addition to VW-SCs, progenitor cells of different origins such as BM and blood circulation and MSCs from different tissues have been reported to differentiate into SMCs. This may be the reason why SMCs of the vascular tunica media and SMCs in atherosclerotic plaques were described to be heterogeneous [for details, see review by Orlandi and Bennett (78)]. Finally, similar to embryonic mesangioblasts, cells derived from the blood vessels of skeletal muscle tissue apparently have the capacity to differentiate to skeletal muscle cells and form myotubes as shown in a mouse model (29). These data strongly suggest that the blood vessel wall does not only contain vascular progenitors such EPCs, smooth muscle progenitors, and multipotent MSCs but also organ- and tissue-specific progenitors depending on the organs and tissues supplied by the vessels. So far it is unclear whether these tissue-specific progenitors are generated from the VW-MSCs under tissue-specific conditions or exist in addition to the VW-MSCs.
Collectively, these studies suggest that a “glue” composed of multipotent ancestors VW-MSCs and VW-HSCs and different types and subsets of progenitor cells with a capacity to differentiate to mature VW cells, blood cells, and organ-specific cells is localized in the VW (Fig. 7), precisely in the periendothelial space of adult microvessels as well as within the adventitia of medium-sized and large blood vessels. This cellular potential is relevant not only for repair and self-renewal of vascular cells and/or vasa vasorum but also for local capacity of neovascularization in pathological processes such as tumor growth, metastases, atherosclerosis, and ischemic revascularization. Moreover, the multipotent VW-resident ancestors, particularly VW-MSCs, may have the capacity to generate not only VW cells but also other organ-specific mature cell types. This appreciation of the stem cell potential of the VW is fundamental to the clinical progression of cell-based therapies.

Clinical Relevance of VW-Resident Stem and Progenitor Cells
As VW-resident multipotent ancestors such as VW-SCs and lineage-committed progenitor cells such as VW-EPCs are apparently present in the wall of micro- and macrovessels, and as blood vessels constitute common structure of all organs and tissues, it can clinically be postulated that (i) VW-SCs can be isolated, expanded, and used for cellular therapeutics, and (ii) they may be involved in different disorders. Their endogenous activation, mobilization, and differentiation processes could be manipulated therapeutically.
VW-SCs as cellular therapeutics
To replace tissue damage by stem cell or tissue pieces created from stem cells is a great promise of current medicine, which receives progression in basic research. Although we are still far away from understanding how stem and progenitor cells orchestrate in adult tissue, numerous studies applying BM-derived cells as a whole mixture or in form of sorted cell types reported clinical benefit, particularly in cardiovascular disorders (45, 54, 61, 73). As fragments of human adult arteries, for example, internal thoracic artery, and veins, for example, saphenous vein, are readily available without any burden for patients from bypass surgery, the ancestor VW-SCs and progenitors may serve as source for individual autologous cellular therapy. Experiences with these cells until now imply that they are easily extractable from the vessel wall and expandable in vitro under culture conditions to reach a sufficient amount of cells for therapeutic application. But to this aim it is necessary to better characterize not only VW-SCs but also their interactions and interdependences to each other to prepare the best disease-specific cell mixture.
Endogenous therapeutic manipulation of VW-SCs
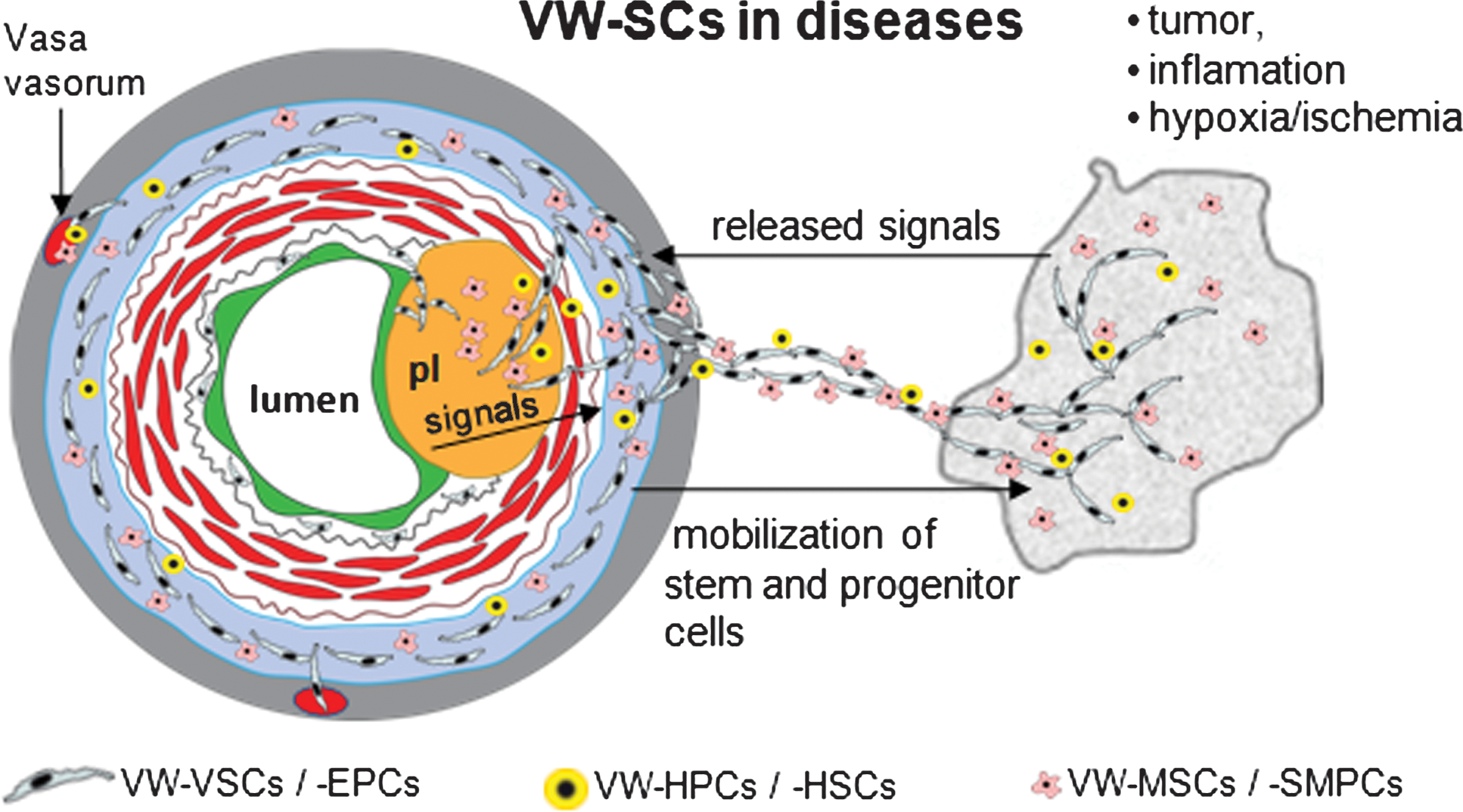
The unique localization of the VW-SCs within the VW sets forth the hypothesis that these cells can be activated and mobilized from their niche by local pathological processes such as atherosclerotic or interstitial processes, primary tumors or metastatic lesions, and ischemic events. Further, their close vicinity to the vasa vasorum of the large and medium-sized arteries and veins and to the ECs in microvessels suggest that they can be mobilized into the blood circulation in the presence of systemic and/or local stimuli.
Publications of the last decade highlight the role of EPCs in tumor vascularization, either directly by integration into tumor vessels or indirectly by supporting tumor angiogenesis (3, 40, 41, 77, 83). But, apparently, only a small fraction of these cells is physically integrated into the new vessels (40, 83), and most recently, in a study using transgenic mice for endothelial-SCL-Cre-ER(T), no contribution of BM-EPCs to tumor vessel formation was observed (46). Further studies using genetic models showed that BM-derived progenitor cells are localized in the perivascular space rather than in the vessel wall (28). These inconsistencies are partially caused by different methods and techniques used in different studies but they have to be overcome to uncover the exact role of EPCs in new vessel formation. In this context, the lack of procedures to distinguish EPCs of different sources, particularly those of BM origin and that present in peripheral tissue niches, may lead to partially controversial findings. And finally, we have to consider the participation of EPCs during transient steps of vascular development. Recent findings presented by Nolan et al. underline this assumption showing that after marking the BM cells genetically, BM-EPCs were integrated into the tumor vessels during early vascularization but they were diluted by EPCs coming from the periphery at later stages of tumor development (77). Interestingly, VW-EPCs can be mobilized from the vessel wall by coincubation with tumor cells (120). Thus, a part of the aforementioned peripheral EPCs may be recruited locally from the vessel wall (Fig. 8). Recent data suggest that proangiogenic signals released by tumor cells located in close vicinity to preexisting blood vessels may reach the adventitial VW-EPCs easier and faster than C-EPCs and mature ECs because of the barrier innate to the multiple cell layers of the VW. Also, VW-EPCs associated to the wall of adventitial vasa can be activated by this manner. Thus, the co-option of preexisting blood vessels during the growth and expansion of tumor cells could first induce a mobilization of local VW-EPCs to form new vessels and subsequently activate mature ECs as well as C-EPCs. This might explain why probably the direct participation of BM-EPCs on tumor vascularization is not as high as generally expected. Further, these data also imply that tumor blood vessels are composed by a heterogeneous pattern of ECs, which may have clinical relevance for antiangiogenic therapy. New in vivo experimental models are needed to determine the direct and indirect contribution of EPCs of different niches to tumor vascularization. Further, it is conceivable to postulate that also VW-HSCs might be activated by tumor cells. Together with VW-EPCs, they create a local milieu promoting tumor vascularization and metastasis. Both EPCs and HPCs have been shown to be required for metastatic growth of tumor cells at distant sites (114). Further studies are needed to explore the local interaction between VW-EPCs and VW-HPCs/HSCs and to determine the role of it in vascularization of primary tumors and metastatic lesions.
Further, a disease with direct endothelial involvement is atherosclerosis. Transmigration of macrophages and the deposition of lipids into the subendothelial space make out the first cellular and structural events of developing plaque. The subsequent accumulation of SMCs in the intima leads to the formation of neointima, which narrows the arterial lumen with increasing size (43, 115). However, cellular treatment with EPCs contributes to the restoration of the endothelial monolayer and to the repair of endothelial damage (113), but the mechanisms behind this effect remain not entirely elucidated. Published data showed an improvement in the regeneration and repair of endothelium after systemic transfusion of the BM-derived cells containing EPCs (55, 75, 118). This is further supported by preclinical and clinical human studies demonstrating that the cardiovascular risk inversely correlates with the number and migratory capacity of EPCs (34, 69, 111). In contrast, recently published reports documented that the application of EPCs results in an acceleration of plaque size and instability (42, 118), probably due to increased plaque vascularization and by suppression of plaque stability factors (43). Similarly, in another study, the transfer of BM-derived MNCs resulted in increased atherosclerosis (98).
Until now, reports about the role of EPCs in atherosclerosis are largely limited to the role of BM-EPCs and C-EPCs, and little is known about the role of VW-SCs in this process. Emerging data suggest that CD34+CD31− VW-resident cells are capable of migrating through the vascular media toward endothelial intima and form capillary-like vessels within VW (120). A more recent publication demonstrated that these cells contain a subset of cells with MSC characteristics, which in turn can differentiate to vascular SMCs and cover the endothelial vessels (14). Finally, VW-HSCs and probably also VW-MSCs possess the properties to generate macrophages (120). As easily imaginable from these results, the VW-resident stem and progenitor cells have the potential to deliver all cell types relevant for initiation and progress of atherosclerosis (Fig. 8): (i) mature ECs from VW-EPCs may provide plaque vascularization, a process increasing the plaque growth and worsening the clinical outcome; (ii) SMCs generated from the VW-MSCs or Sca-1+ SMC progenitors may contribute to the formation of neointima, leading finally to the closure of the arterial lumen; and (iii) as shown for macrophages, the generation of local inflammatory cells from VW-resident progenitors may promote atherosclerosis because it is well known that inflammation is a major process in this disease. The crucial role of such a microenvironment for plaque growth and instability has been recently shown for mast cells accumulated in the vascular adventitia of apolipoprotein E-deficient mice (12). Further, in a recent study using chimeric rats generated by rescuing lethally irradiated animals with green fluorescent protein(+) BM, it was shown that the major fraction of cells forming the neointima are not of BM origin (90), thereby suggesting that the neointima is likely to be generated from progenitors of the VW. The essential role of the vascular adventitia in this process is further underscored by studies showing that the adventitia contributes to plaque development approximately 28 times more than the luminal side of the vessel wall (67). Indeed, there is a correlation between adventitial neovascularization and aortic plaque progression in apolipoprotein E-deficient mice (68).
Collectively, this evidence suggests that VW-resident stem and progenitor cells can be activated by tumor cells outside the vessel wall but by processes inside the vessel wall such atherosclerosis. Particularly, the composition by multipotent ancestors and different types of progenitors within the vascular adventitia provides a cellular pool with the capability to differentiate into mature cell types such as ECs, SMCs, fibrocytes, and inflammatory cells (Fig. 8). Moreover, it is conceivable that VW-EPCs, -MSCs, and -HSCs play a dual role in vascular diseases. Although EPCs are probably beneficial in the treatment of endothelial injury, dysfunction, and repair, they and the other types of stem and progenitor cells may accelerate atherosclerosis development by providing new vessels and inflammatory cells once a subendothelial plaque is formed.
Concluding Remarks
Evidences in the last few years show that the wall of adult macro- and microvessels as well as that of vasa vasorum harbors multipotent ancestors such as VW-MSCs and VW-HSCs as well as different types of lineage-committed progenitors such as VW-EPCs and Sca-1+ SMC progenitors. Considering the fact that blood vessels are a common structure of all tissues and organs, it is obvious that these cells may have a great potential in biomedicine. But the characterization of these cell types and potential subsets as well as our knowledge remains insufficient to estimate their clinical relevance and applicability. For example, it remains unexplored whether the putative “hemangioblast” or “mesangioblast” resides in the adventitial niche of adult blood vessels. Until now, VW-SCs and progenitors have not been tested in clinical trials as it has been done for BM- and C-EPCs or whole BM-derived cells, but procedures for the clinical use of VW-SCs and progenitors are being developed. Fragments of human arteries and veins are easily available, and thus, besides the big expectations in iPS cell ancestors, VW-SCs and progenitors extracted from such adult vessel fragments can be used for autologous cell therapeutics. Further, better knowledge of mechanisms how VW-SCs and progenitors can be manipulated by new drugs endogenously can also help to set up new therapeutic avenues.
Footnotes
Acknowledgment
The authors thank Jürgen Heger for his excellent technical assistance in production of graphical representations.