
Abstract
Supplementation of standardized fermented papaya preparation (FPP) to adult diabetic mice improves dermal wound healing outcomes. Peripheral blood mononuclear cells (PBMC) from type II diabetes mellitus (T2DM) patients elicit a compromised respiratory burst activity resulting in increased risk of infections for the diabetic patients.
Introduction
Innovation
This work provides the first evidence demonstrating that respiratory burst dysfunction in monocytes from diabetic patients may be corrected by fermented papaya preparation (FPP). Induction of p47phox phosphorylation as well as upregulation of Rac2 transcription emerged as two potential underlying mechanisms. Because FPP has a long track-record of safe human consumption (1, 27 –29, 44) and has proved to be beneficial in influencing wound healing outcomes (15), findings of this study lay a strong rationale for the design of phase-II clinical trial testing the effects of FPP supplementation on wound infection outcomes of type II diabetes mellitus patients suffering from chronic wounds.
Carica papaya Linn is widely recognized as a medicinal fruit (2). Fermented papaya preparation (FPP), a granular substance, is available over the counter (32). FPP possesses antioxidant properties (3, 11, 22, 35) that provide benefit against age-related complications (27). FPP is also known to protect red blood cells against oxidative damage (27, 28) and help against severe forms of thalassemia (1). Several independent observations convergently point toward the hypothesis that treatment with papaya preparations may facilitate wound healing responses (2, 17, 21, 30, 33, 34). Chronic wounds represent a major public health problem in diabetics. Our previous studies have demonstrated that wound-site macrophages of diabetics are compromised in their ability to support wound healing (23). Recently, our laboratory reported first evidence demonstrating that FPP may improve diabetic wound outcomes by specifically influencing the response of wound-site macrophages and the subsequent angiogenic response (15). FPP has a long track-record of safe human consumption (26, 28). The objectives of the current study were twofold: to determine whether FPP is able to improve inducible respiratory burst outcomes in peripheral blood mononuclear cells (PBMC) of diabetic patients, and to investigate the underlying mechanisms. The overall goal was to develop data toward future phase-II clinical studies.
Results
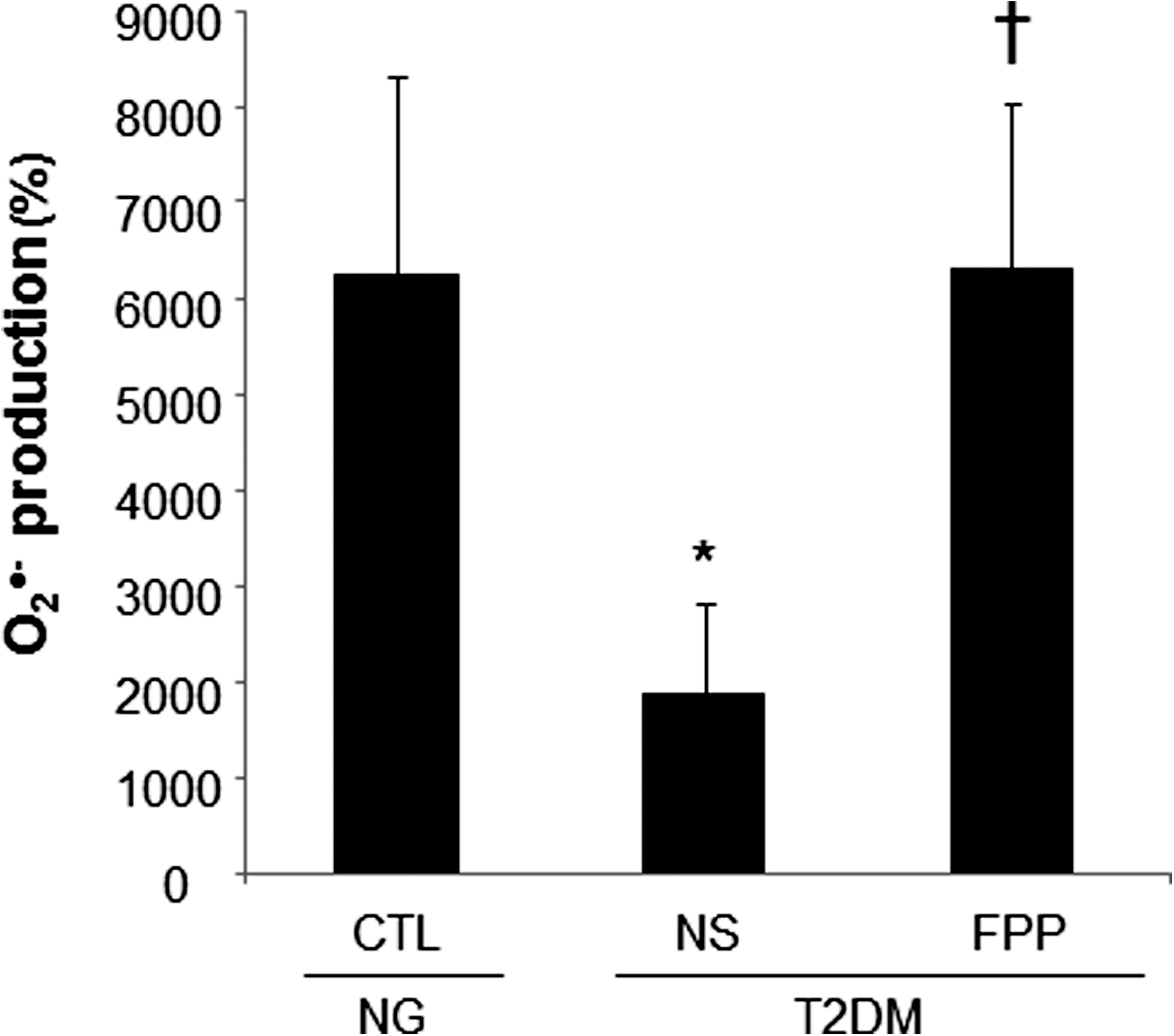
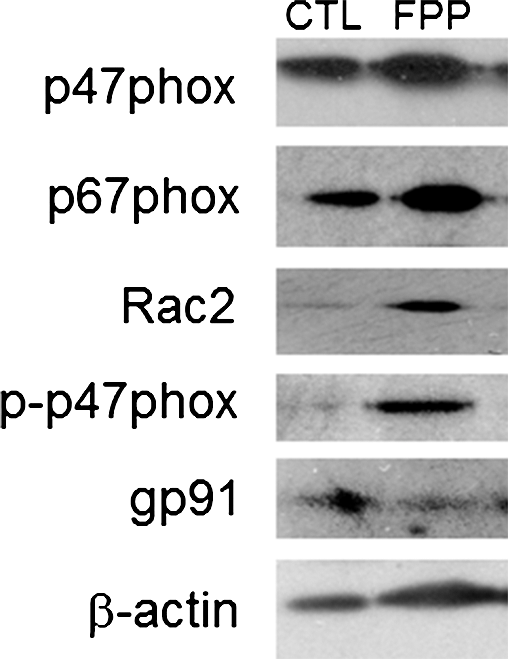
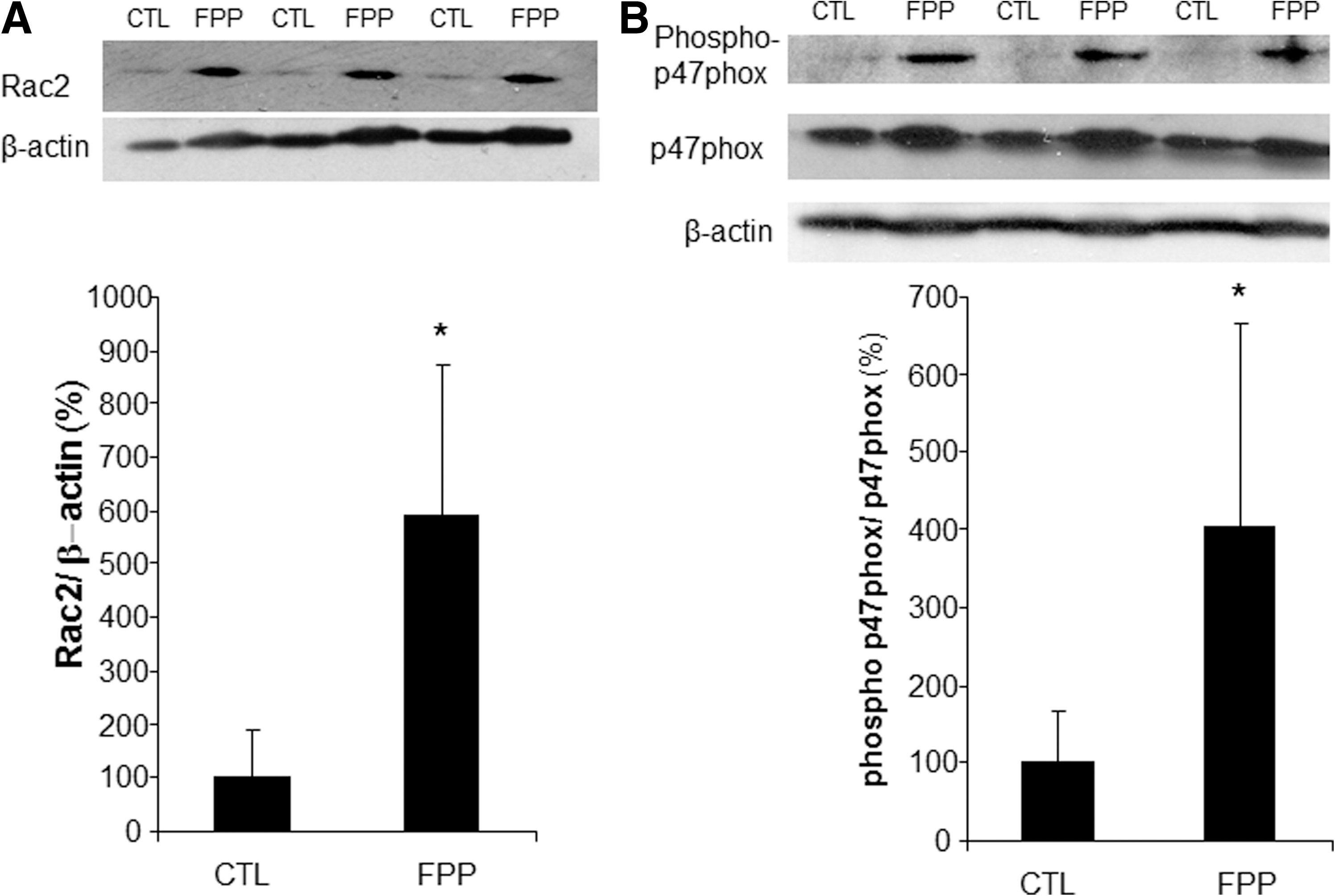
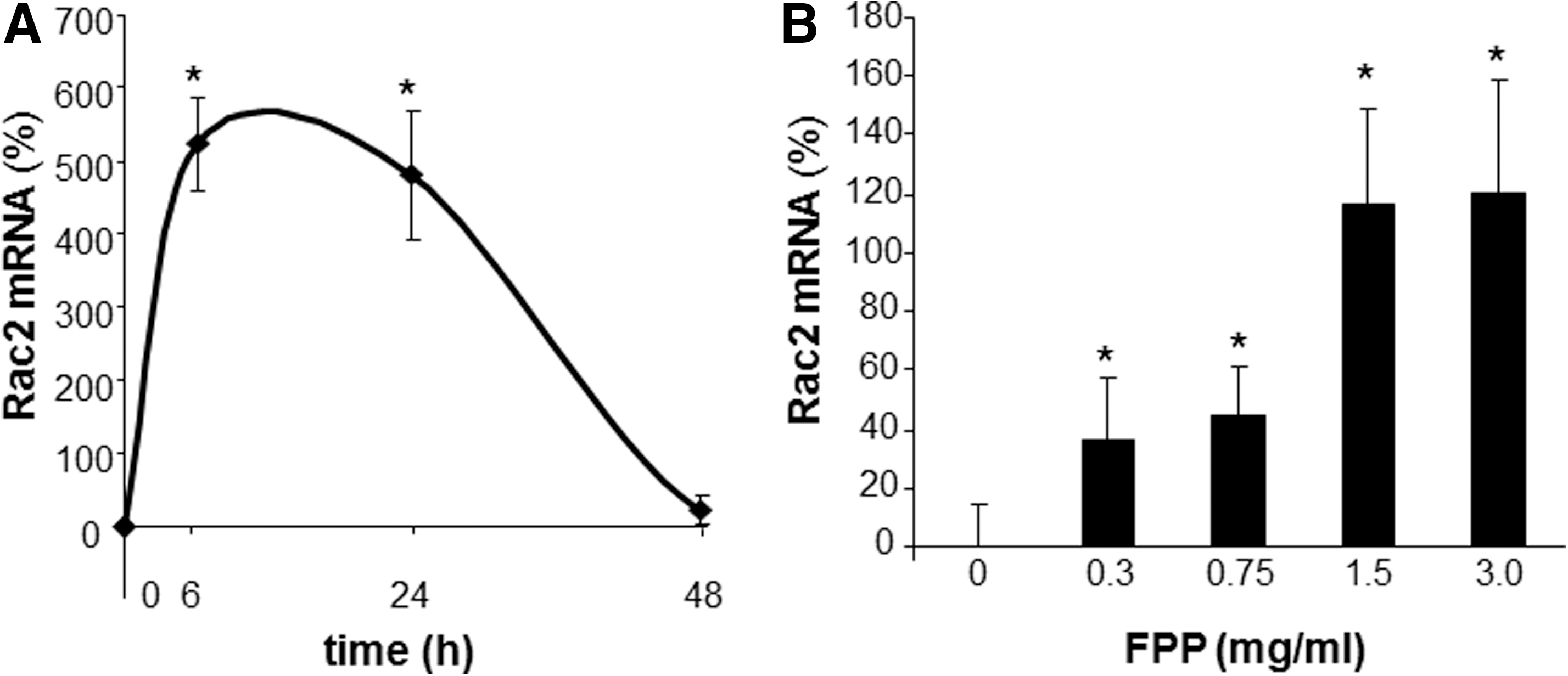
Phorbol 12-myristate 13-acetate (PMA) stimulated a potent induction of superoxide anion production from PBMCs. This response was severely blunted in PBMC isolated from T2DM patients compared to results from normoglycemic control human subjects. Intriguingly, the compromise in inducible respiratory burst activity in T2DM PBMC was corrected in response to exposure to FPP (Fig. 1). FPP induces ROS expression in cells isolated from normoglycemic subjects; the extent of induction, however, is less (20% vs. 230% in T2DM) compared to cells from T2DM (data not shown). NADPH oxidase represents the primary enzyme that is responsible for PMA-inducible respiratory burst production in PBMC. To investigate the underlying mechanisms of FPP action on respiratory burst, isolated PBMCs were cultured in presence of FPP for 24 h. Expression of the major subunits of NADPH oxidase complex was determined using Western blot (Fig. 2). Interestingly, the expression of Rac2 was significantly upregulated after FPP supplementation (Fig. 3A). FPP contains large amounts (∼90%) of carbohydrates (29). We have previously shown that matching amount of glucose supplementation did not result in ROS production as observed after FPP supplementation (15). To confirm these observation in ROS production by human PBMC, glucose (matching to that present in FPP) treatment to the PBMCs was performed (data not shown). Glucose treatment alone did not further stimulate PMA-induced ROS production in PBMCs as observed after FPP treatment, suggesting that FPP is specifically effective in inducing ROS production, which is independent of the glucose present in this nutritional supplement. In addition to upregulation of Rac2 expression, FPP exposure also resulted in increased phosphorylation of p47phox (Fig. 3B). With the goal to understand how FPP induces Rac2 expression, the effect on gene transcription was investigated. Expression of Rac2 mRNA was significantly upregulated after FPP exposure, suggesting that the Rac2 transcriptional machinery was sensitive to FPP. Maximal induction of Rac2 mRNA was noted after 6 h of treatment, indicating that the regulation by FPP was rapid (Fig. 4A). Next, to determine the minimum dose of FPP required eliciting induction of Rac2 mRNA expression, a dose–response study was performed. A dose of 0.3 mg/ml was noted to be sufficient to significantly induce Rac2 transcription, whereas the response was at a peak with 1.5 mg/ml of FPP (Fig. 4B).
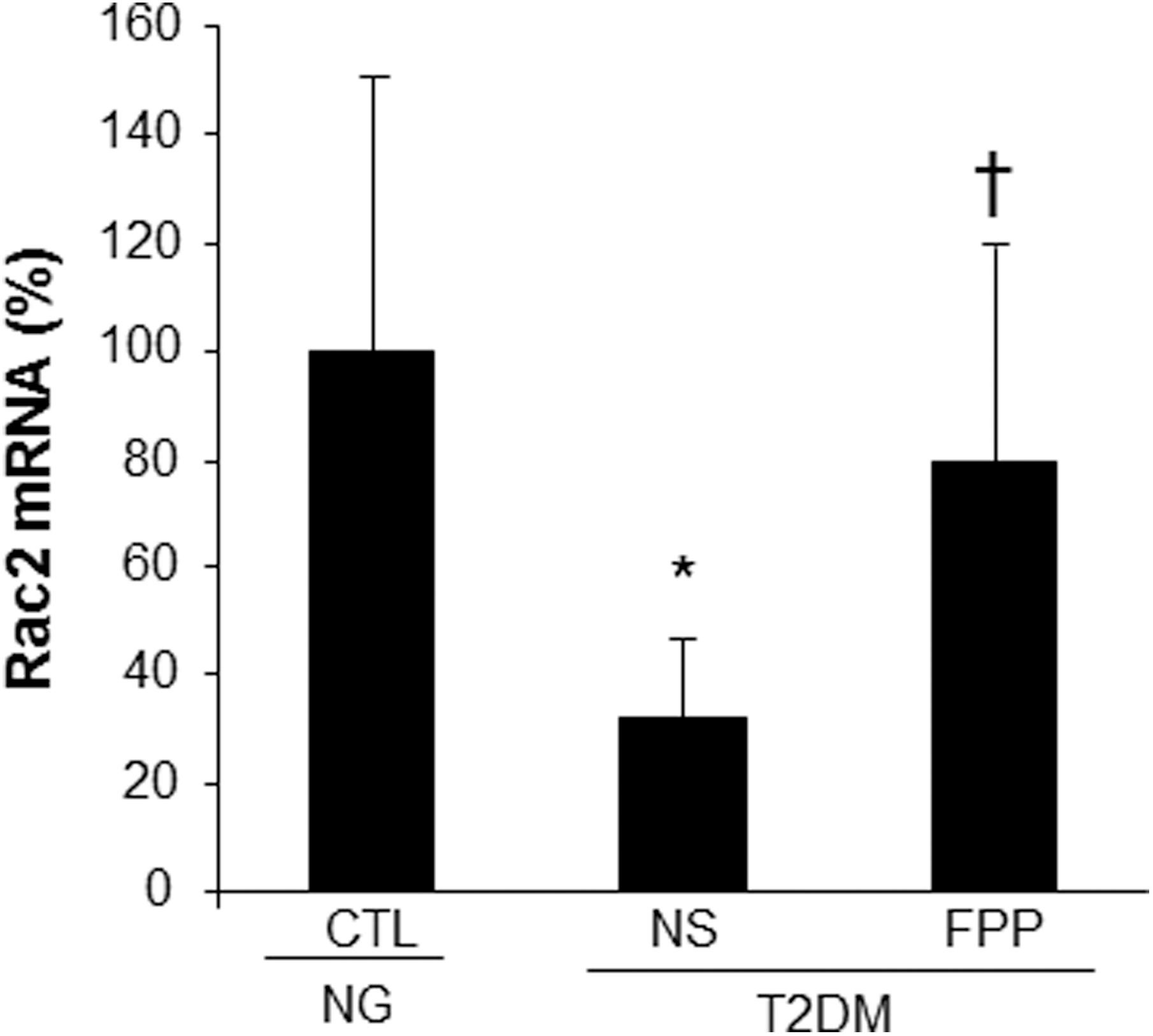
To test whether compromised inducible respiratory burst function in PBMC from T2DM patients was associated with lower Rac2 expression, PBMC were obtained from patients and subjected to RTPCR. Indeed, the expression of Rac2 was noted to be significantly lower in PBMC of T2DM compared to that in PBMC from normoglycemic control donors. The lower expression of Rac2 in PBMC of T2DM was significantly corrected in response to FPP treatment ex vivo (Fig. 5).
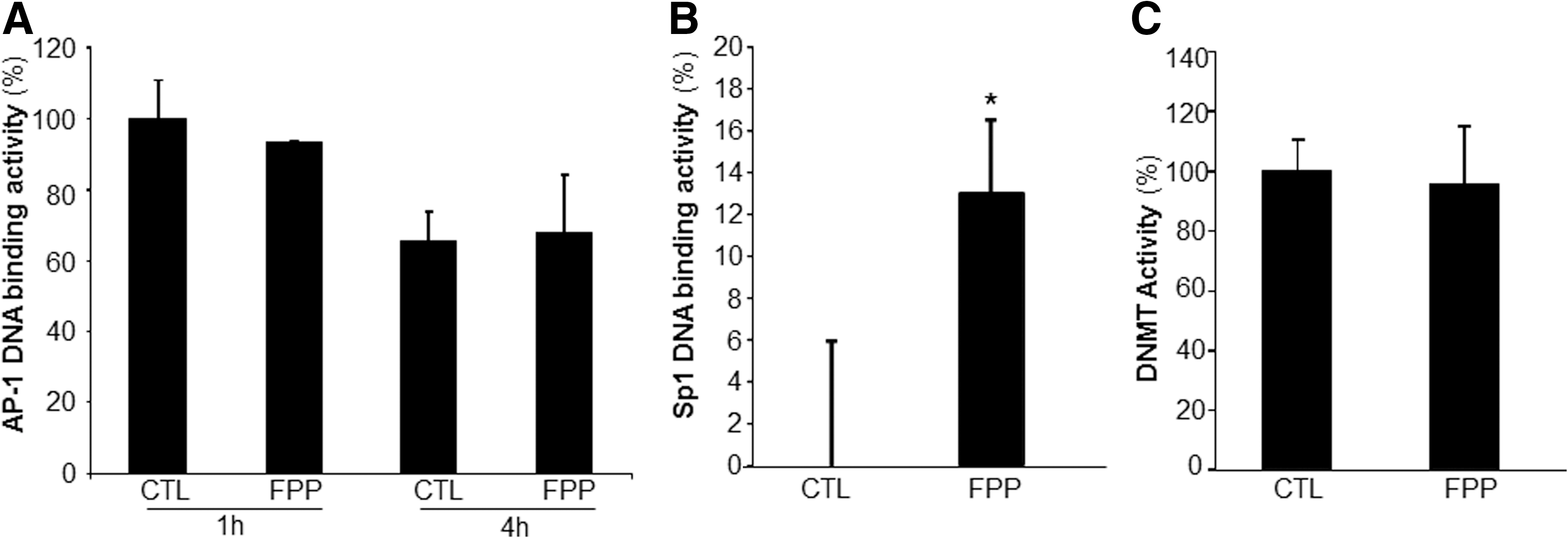
The human Rac2 gene promoter contains binding sites for the transcription factors Sp1 and AP-1. DNA binding activity of AP-1 and Sp1 were therefore tested in response to FPP treatment of PBMC. While FPP had no significant effect on the AP-1 DNA binding activity (Fig. 6A), the Sp1 DNA binding activity was significantly upregulated in PBMC after treatment of the cells with FPP (Fig. 6B). The Rac2 genomic locus exhibits distinct patterns of DNA methylation in expressing versus nonexpressing cells. A role of DNA methylation in Rac2 gene transcription has been shown (25). To determine if FPP influences Rac2 gene expression by modifying DNA methyltransferase (DNMT) activity, DNMT activity was measured from nuclear protein extracts. No effect of FPP supplementation on the DNMT activity was observed (Fig. 6C).
Discussion
Respiratory burst, generating superoxide (O2 •−) and derivative oxidant species, represents a major defense mechanism that fights infection (4). Immune dysfunction, including defective leukocyte respiratory burst activity, has been well documented in T2DM patients (14). Such dysfunction represents a key mechanism underlying the high infection rate in patients with DM (7, 8, 19). Findings of this study demonstrate for the first time that FPP may correct deficiencies in respiratory burst function by inducing Rac2 expression of PBMC obtained from T2DM patients (Fig. 7). Function of the NADPH oxidase complex is a key driver of respiratory burst (6). In response to appropriate stimuli, cytosolic components of this enzyme system, for example, p47phox and p67phox, associate with the membrane-bound gp91phox (NOX2) and p22phox (6). Under appropriate conditions, p47phox is rapidly phosphorylated at several serine residues and undergoes a conformational change. Phosphorylated p47phox then interacts with p22phox and gp91phox to promote the oxidase assembly and subsequent activation (20). Increased phosphorylation of p47phox is likely one of the mechanisms of action of FPP in correcting inducible respiratory burst function of T2DM PBMC.
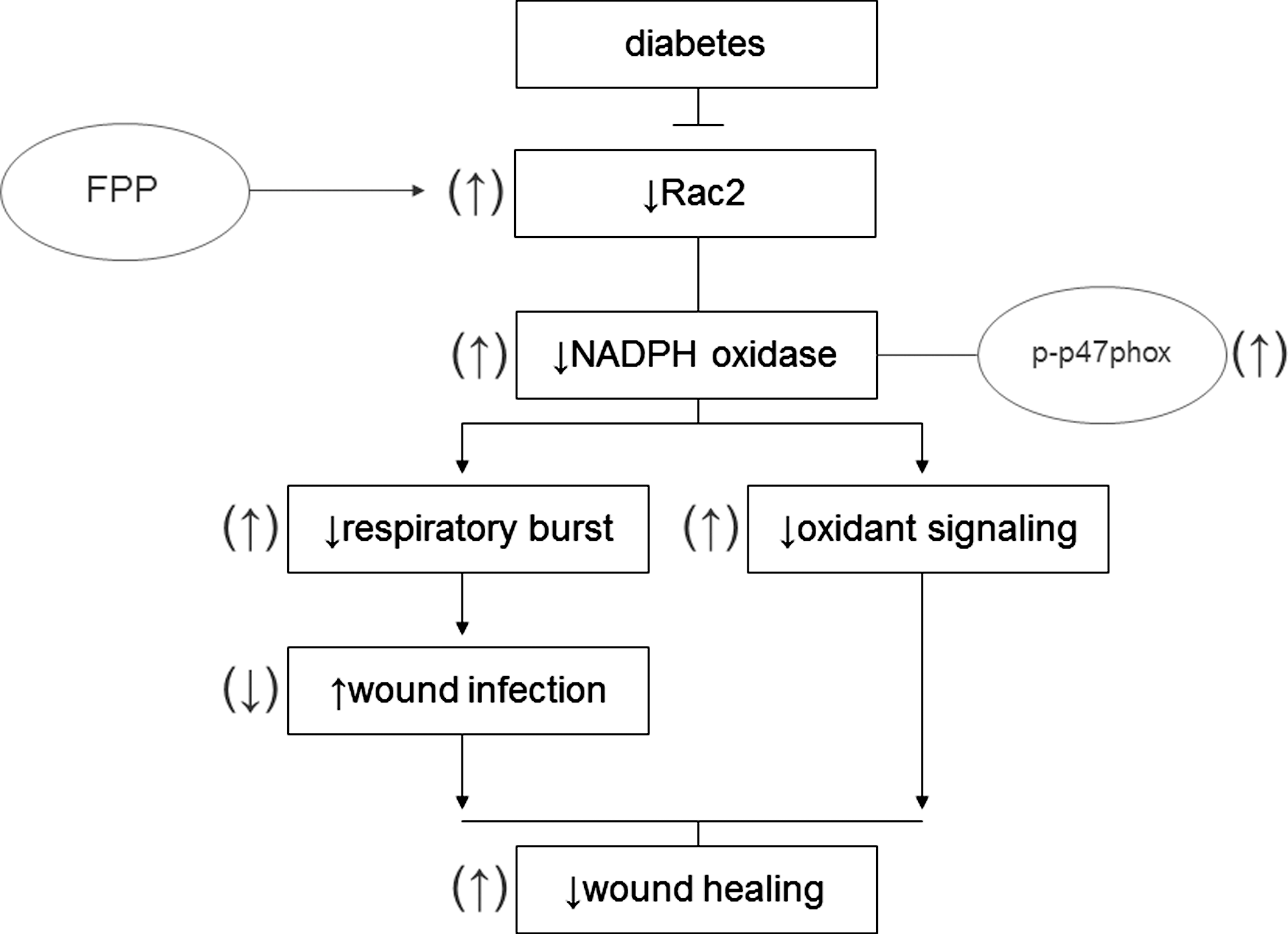
The small GTPase Rac is a member of Rho–GTPase superfamily and acts as a molecular switch to regulate various cellular functions, including NADPH oxidase activation (18). Rac has three isoforms: the ubiquitously expressed Rac1, a hematopoietic cell-specific isoform Rac2, and Rac3, which appears to be expressed in a variety of tissues (43). Rac1 is about fourfold more abundant than Rac2 in monocytes/macrophages and was suggested to be primarily involved in NADPH oxidase activation in monocytes (13). Macrophages derived from Rac2-null mice show compromised PMA-induced NADPH oxidase activation pointing toward a key role of Rac2 in enabling macrophage respiratory burst (43). Specifically, Rac2 was shown to be essential for oxidant production elicited by PMA and FcγR stimulation (43). Observations of the current study unveil the potential for FPP to bolster macrophage respiratory burst function via a Rac2-dependent mechanism.
In addition to the improper bactericidal action of the respiratory burst in T2DM patients, oxidant signaling necessary to the wound healing process is likely to be compromised in patients exhibiting improper respiratory burst in response to stimuli (41). NADPH-oxidase-produced superoxide anion is quickly converted to hydrogen peroxide, which continues down a signaling cascade creating more stable oxidants, such as hypochlorous acid, chloramines, and aldehydes, required in the wound healing process (41). FPP-induced correction of respiratory burst function may have implications in improving wound healing outcomes via correction of oxidant signaling pathways.
Human DNA is methylated at the cytosine-5 position within cytosine–guanine (CpG) dinucleotides. This epigenetic modification plays an important role in determining genomic stability and transcriptional regulation (9). The Rac2 genomic locus exhibits distinct patterns of DNA methylation in expressing versus nonexpressing cells (25). Cells that lack Rac2 expression exhibit increased cytosine methylation in the sequences flanking the gene, whereas cells that express Rac2 exhibit increased cytosine methylation within the body of the Rac2 gene (25). Treatment of nonexpressing cells with the DNMT inhibitor 5-aza-2′-deoxycytidine has been shown to be sufficient to induce Rac2 gene expression (25). (−)−Epigallocatechin-3-gallate (EGCG), the main polyphenol antioxidant in green tea, exhibits demethylating activity (10). Therefore, we tested whether FPP modifies DNMT activity. No effect of FPP on DNMT activity excludes this regulatory pathway of FPP-mediated Rac2 gene transcription.
The Rac2 gene is located on chromosome 22q12 and consists of seven exons spanning 18 kb of DNA (31). The human Rac2 gene promoter lacks TATA and CCAAT boxes, utilizing multiple transcription initiation sites, including AP-1 and several putative Sp1 binding sites (25, 31). Overexpression of AP-1 has been shown to be sufficient to induce expression of a transiently transfected Rac2 promoter/luciferase plasmid, but not of the endogenous Rac2 gene (31). In the current study, FPP did not modify AP-1 transactivation in PBMC ruling out AP-1 as a potential mechanism underlying Rac2 induction by FPP. Sp1 binding sites are common in promoters that lack TATA boxes. In mice, Sp1 binding sites in the promoter region play a key role in driving Rac2 gene expression (25). Indeed, we noted that FPP increased Sp1 DNA binding activity in PBMC. This observation suggests that FPP-induced Rac2 gene transcription may be Sp1 dependent. Antioxidants such as N-acetylcysteine are known to induce Sp1-dependent transcription of the CDKN1A gene by increasing the phosphorylation of the C terminus of Sp1 (24). FPP possesses potent antioxidant functions (22). Whether the antioxidant activity of FPP is involved in the regulation of Sp1 transactivation remains unresolved.
Materials and Methods
Human subjects and sample collection
All human studies were approved by The Ohio State University Institutional Review Board. Subjects (adults 40–60 years) participating in the study were either healthy normoglycemic or clinically diagnosed as T2DM with good glycemic control, that is, hemoglobin A1c (HbA1c)≤7%. T2DM subjects were recruited from the Diabetes Clinic at The Ohio State University Medical Center. Subject demographics are presented in Table 1. T2DM subjects, either immunosupressive or taking peroxisome proliferator-activated receptor gamma medications, were excluded from the study. Peripheral blood (60 cc) was drawn by venipuncture. For isolation of PBMC, blood samples were collected in sodium–heparin-coated blood-chemistry tubes (BD Medical, Franklin Lakes, NJ). For studies investigating the underlying mechanisms of FPP action, PBMC were also isolated from banked source leukocytes packets obtained from the American Red Cross of Greater Columbus (Columbus, OH).
BMI, body mass index; HbA1c, hemoglobin A1c; SD, standard deviation.
Human PBMC isolation and culture
Either fresh blood or source leukocytes were diluted 1:1 using sterile Dulbecco's phosphate buffered saline (Mediatech, Inc., Manasses, VA). PBMC were isolated using Ficoll density centrifugation followed by sorting with anti-CD14 coated magnetic microbeads (Miltenyi Biotech, Auburn, CA) as previously described (36). Purified CD14+ PBMCs were then cultured under standard conditions (5% CO2, 37°C, humidified incubator). RPMI 1640 (Mediatech, Inc.) supplemented with 5% heat-inactivated fetal bovine serum (Atlanta Biologicals, Inc., Lawrenceville, GA) and 1% antibiotic–antimycotic (Invitrogen Corp., Carlsbad, CA) was used as culture media.
FPP supplementation
FPP (Immun'Age; Osato International, Gifu, Japan) was dissolved in culture media by adding the desired FPP quantity in culture media, followed by sterile filtration using a 0.22-μm vacuum-driven filtration system (Millipore, Billerica, MA). The dose (3 mg/ml) of FPP was chosen based on prior studies (35).
Superoxide anion measurement
Induced superoxide anion generation was measured using a LumiMax® superoxide anion detection kit (Stratagene, La Jolla, CA). Briefly, freshly isolated PBMC were treated with PMA (1 μg/ml). Superoxide was measured at 430 nm using a luminometer (model Lumat LB9507; Berthold Technologies, Bad Wildbad, Germany) (36).
Western blot
Primary antibodies against Rac2 (Millipore, Temecula, CA), Nox2/gp91phox, phospho-p47phox (Abcam, Cambridge, MA), p47phox, and p67phox (Cell Signaling Technology, Beverly, MA) were used to detect the corresponding antigens. β-Actin was considered as housekeeping protein for normalization of sample loading.
Isolation of RNA, reverse transcription, and quantitative real-time polymerase chain reaction
Total RNA was extracted from PBMCs using a mirVana RNA isolation kit (Ambion, Austin, TX) according to the manufacturer's suggested protocol. mRNAs were quantified by real-time polymerase chain reaction assay using SYBR green-I (Applied Biosystems, Carlsbad, CA) as described previously (36, 38
–40). β-Actin was used as the housekeeping gene. ΔΔCt comparative analysis was used to normalize gene expression data against housekeeping gene. The primer-set used for individual genes is listed below: Rac2 5′ GCC TGG CAC TGG CCA AGG AG 3′ 5′ CTA GGT GGG AGC GCT GGG GT 3′ β-Actin 5′ GTA CCA CTG GCA TCC TGA TGG ACT 3′ 5′ CCG CTC ATT GCC AAT GGT GAT 3′
DNA binding activity of Sp1/Sp3
A TransAM Sp1/Sp3 family transcription assay ELISA-based kit (Active Motif, Carlsbad, CA) was used as described (37).
DNMT activity
DNMT activity was measured from nuclear protein extracts using DNMT activation/inhibition assay (Active Motif) according to the manufacturer's protocol.
Statistics
Data are reported as mean±standard deviation of at least three experiments. Comparisons among multiple groups were made using analysis of variance. p<0.05 was considered statistically significant.
Footnotes
Acknowledgment
Supported by DK 076566 to S.R. and in part by a funding from Osato Research Institute, Japan.
Author Disclosure Statement
The authors declare that FPP and partial research funding was provided by Osato Research Institute of Japan.