
Abstract
Introduction
Innovation
The current study demonstrated for the first time that the autoantibody against the angiotensin II type 1 receptor (AT1-Abs) (an adverse factor existing in pre-eclamptic patients as well as in a small number of normotensive pregnant women) was transferred from mother rats to their offspring via the placenta/milk and caused insulin resistance and increased susceptibility to metabolic syndrome (MetS), a major risk factor for cardiovascular disease. These findings identified that AT1-Abs in mothers is a risk factor that should not be ignored because of the possibility of the development of MetS in their descendents.
Recently, evidence about the autoantibody against the angiotensin II type 1 receptor (AT1-Ab), first detected in nearly all women with severe pre-eclampsia and in a small number of normotensive pregnant women, might lead to the fact that IUGR has emerged (12). This autoantibody specifically recognizes the functional epitope of the second extracellular loop of the AT1 receptor (amino-acid residues 165–191, AT1R-ECII), possessing AT1 receptor agonist-like pathological effects (29). Previous studies demonstrated that AT1-Ab might cause impaired placental perfusion via varied mechanisms (30), such as enhancing NADPH-reactive oxygen species production (4), minimizing trophoblast invasion (32), and increasing placental cell apoptosis in vitro (12). Interestingly, a clinical study reported that the AT1-Ab was also detected in the fetal cord blood of antibody-positive mothers (10). The onslaughts just mentioned may contribute to pathological placental injury, limiting intrauterine fetal growth and maturation. However, there is a lack of more direct evidence demonstrating how the antibody affects offspring development. The potential path by which the maternal AT1-Ab transference to offspring occurs is unclear. The biological properties of the AT1-Ab within offspring require further elucidation. Most importantly, whether the antibodies might exert negative effects on offspring health in later years is completely unknown.
Therefore, the aims of the current study were to (i) determine the route(s) of AT1-Ab transference from mother rats to offspring; (ii) study whether the AT1-Ab present in newborn rats is biologically active; and (iii) investigate whether such offspring have increased susceptibility to MetS on maturation.
Results
Successful establishment of AT1-Ab-positive female rat models by active immunization
Of the 12 actively immunized female rats, 10 generated increased serum AT1-Ab levels 2 weeks after initial immunization, determined by an enzyme-linked immunosorbent assay (ELISA). As seen in Figure 1A, the target antibody level peaked at the 8th week (optical density [OD] value, 2.76±0.09 vs. 0.33±0.02, p<0.01 vs. the vehicle group at the same time point), indicating a successful establishment of the active immunization model. The AT1-Ab was not detected in the vehicle group. Additionally, AT1-Abs were of the immunoglobulin G (IgG) isotype (Fig. 1A), not IgM (OD value, 0.07±0.00 vs. 0.08±0.00, p>0.05 vs. the vehicle group, Fig. 1B) or IgA (OD value, 0.11±0.01 vs. 0.10±0.01, p>0.05 vs. the vehicle group, Fig. 1C). Immunized rats not producing AT1-Abs were not used in this study.
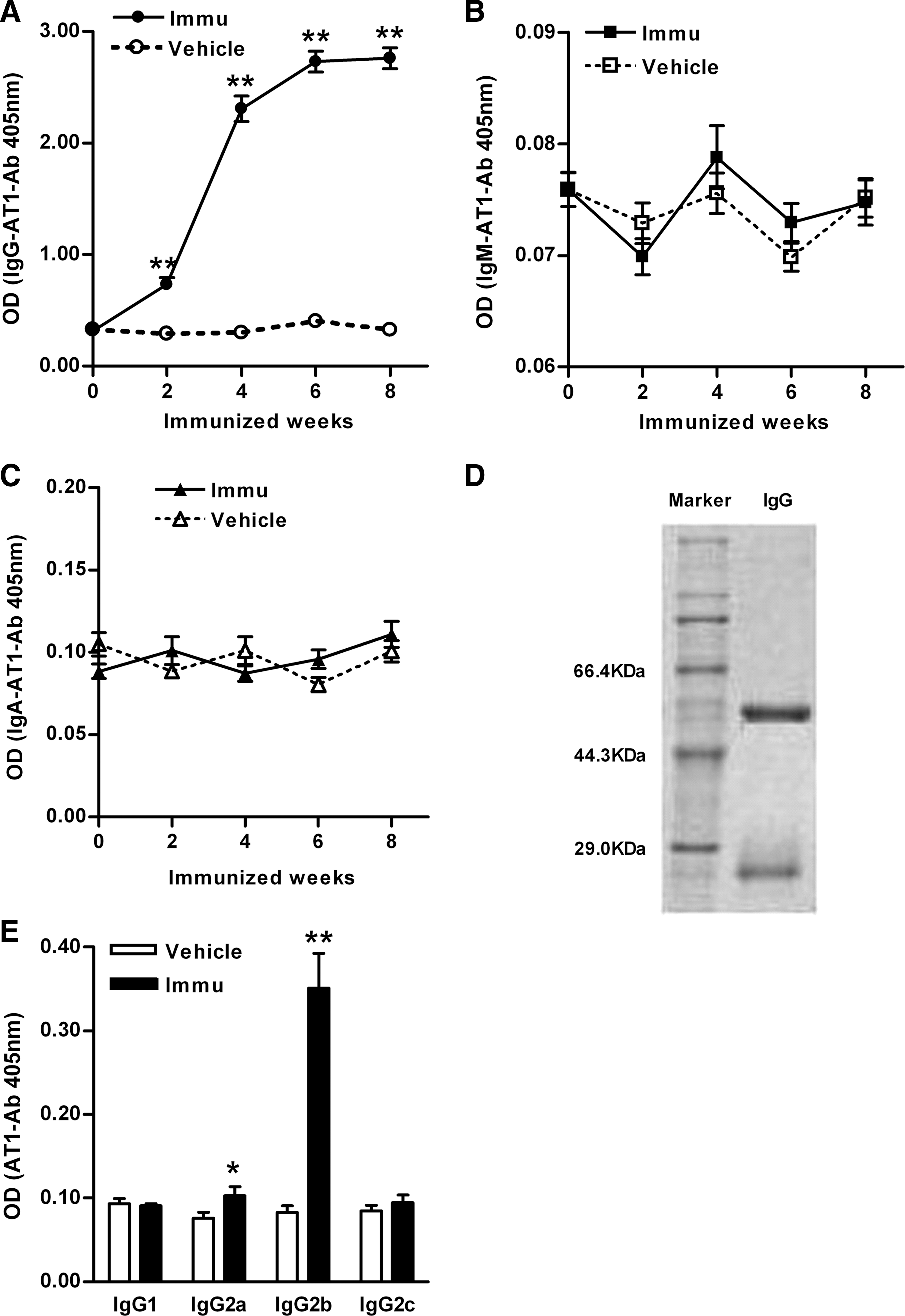
Sodium dodecylsulfonate–polyacrylate gel electrophoresis (SDS-PAGE) showed that two strong bands were seen at 55 and 25 KDa, representing the heavy and light chains of total IgGs from the immunized rats, respectively (Fig. 1D). The IgG subclass of AT1-Abs in immunized rats was determined to be mostly IgG2b (OD value, 0.35±0.04 vs. 0.08±0.01, p<0.01 vs. the vehicle group, Fig. 1E), with a minor percentage belonging to the IgG2a subclass (OD value, 0.10±0.01 vs. 0.08±0.01, p<0.05 vs. the vehicle group, Fig. 1E).
AT1-Abs transport capacity and the pathways involved
By 20 days postconception, the sera of intrauterine fetal rats of immunized mothers were AT1-Ab-positive, with a P/N value ([specimen OD−blank control OD]/[negative control OD−blank control OD]) (19) >2.1 (OD value, 2.32±0.10) according to the ELISA detection. The vehicle group sera were AT1-Ab-negative, with a P/N value <1.5 (OD value, 0.13±0.01, p<0.01 vs. the immunized group, Fig. 2A). As seen in Figure 2B, strong staining was visualized within the villus placental trophoblast cell layer and fetal vascular endothelium (short arrow) of the immunized group, indicating the AT1-Ab presence. The AT1-Ab distribution was very similar to that of total placental IgGs (long arrow). The staining pattern results suggested general placental IgG crossing in the vehicle group, but no AT1-Abs were identified.
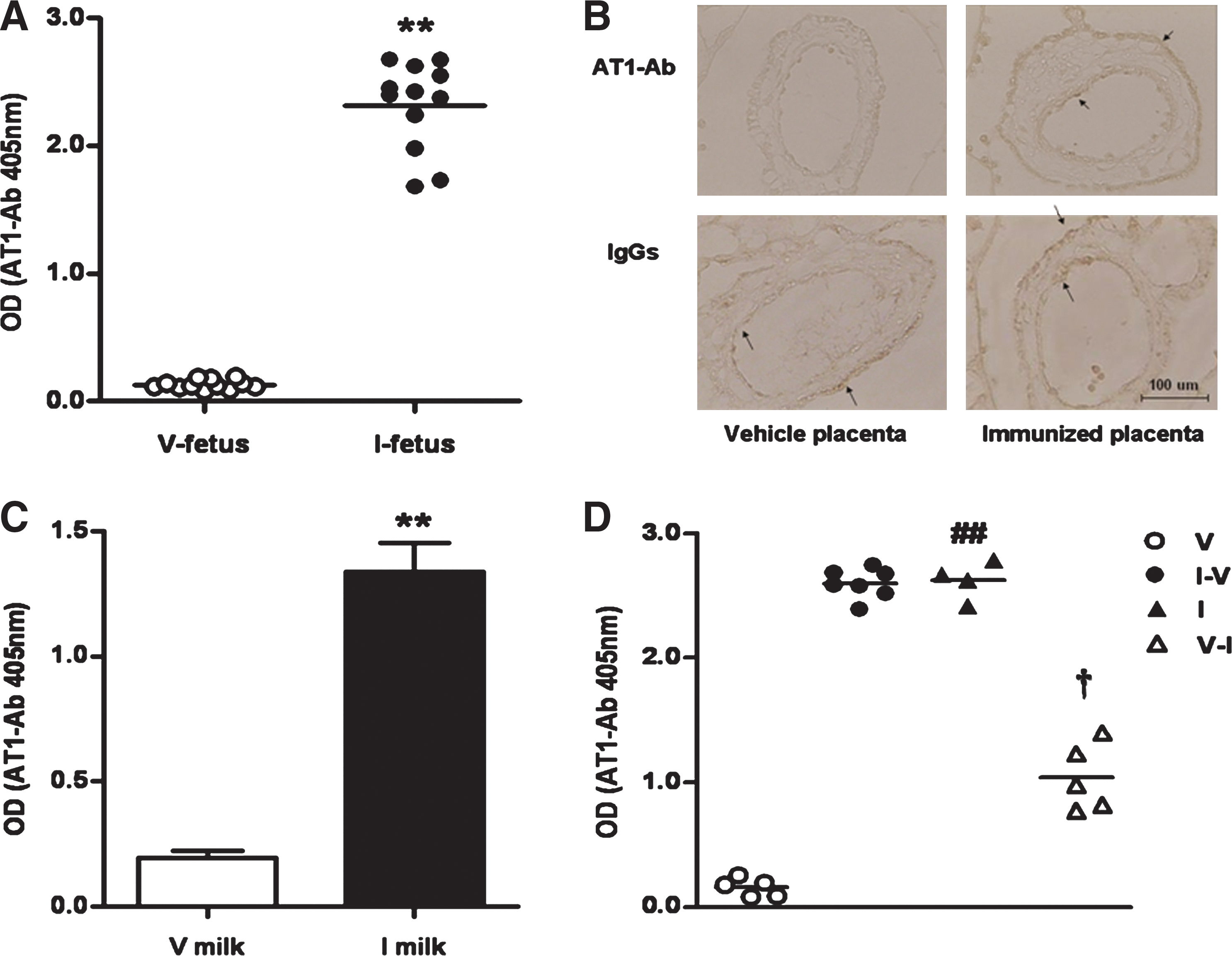
Figure 2C demonstrated that milk obtained from the neonatal gastric contents (7) in the immunized group contained significantly higher AT1-Ab titers than the vehicle group after 1 week of nursing (OD value, 1.34±0.11 vs. 0.19±0.03, p<0.01). To obtain more evidence that mothers' milk is another route via which AT1-Abs are passed to newborns, cross-feeding was initiated. Three pairs of mother rats met the inclusion criteria for this experiment phase (one pair was excluded due to refusing breastfeeding). After 7 days, AT1-Abs were manifested in the sera of the female newborn vehicle rats breastfed by the immunized mothers (OD value, 2.60±0.05 vs. 0.16±0.03, p<0.01 vs. male newborn vehicle rats, Fig. 2D). In contrast, the AT1-Ab levels in the sera of the immunized female newborn rats fell sharply after they had been fed by the vehicle mother rat milk (OD value, 1.04±0.12 vs. 2.62±0.08, p<0.01 vs. male newborn rats in the immunized group). The experiment was repeated with mother rats exchanging male infants, with similar results (Supplementary Fig. S1; Supplementary Data are available online at
Biological activity of AT1-Abs in the immunized group's newborns
Elevated titers of AT1-Abs were still present in the sera of 22-day-old (normal weaning age) immunized group rats, but not in the vehicle group offspring rats (2.22±0.10 vs. 0.13±0.01, p<0.01, Fig. 3A). Antibody titers gradually declined and were undetectable after the 7th week.
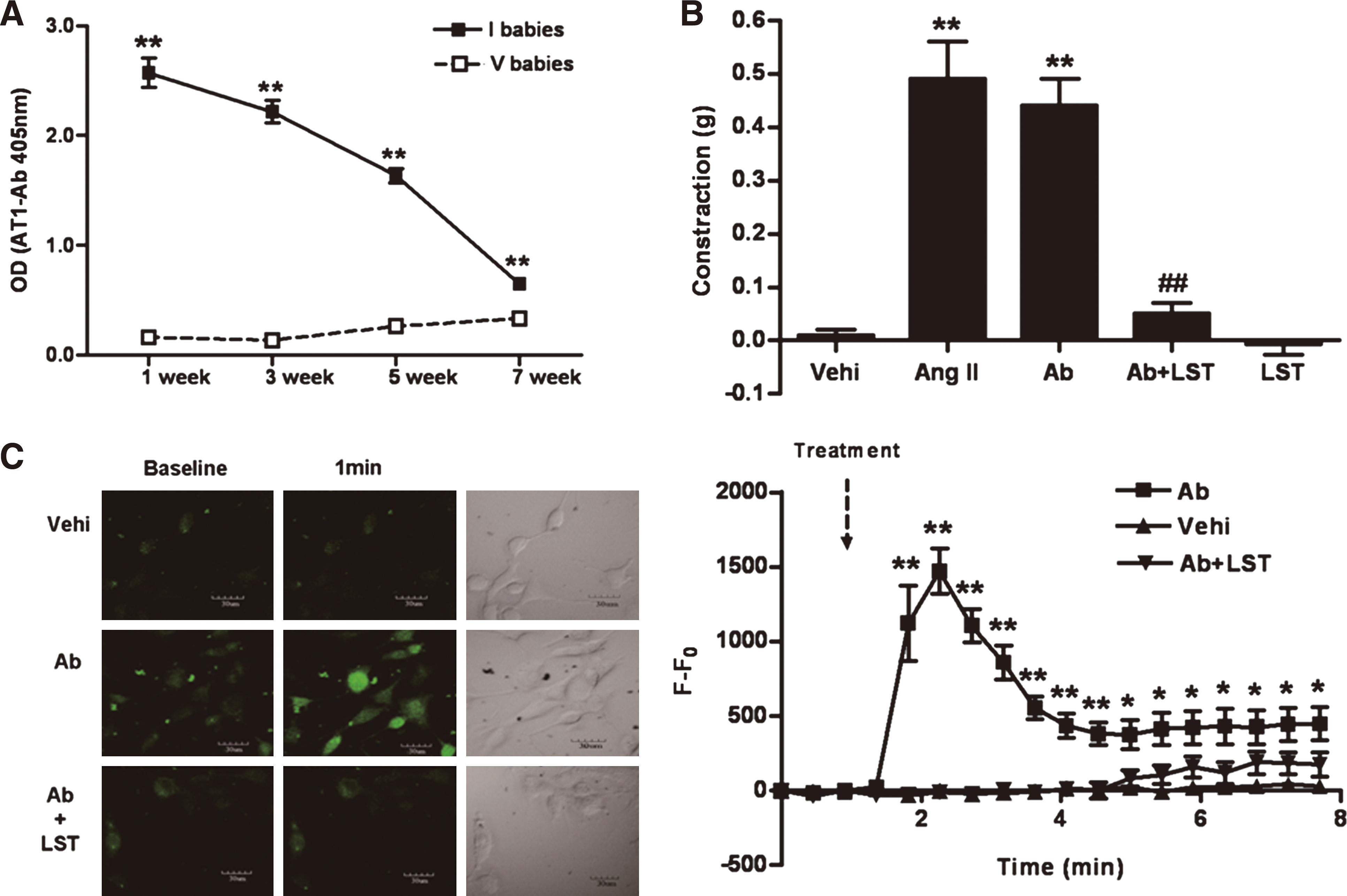
As shown in Figure 3B, 1 μM IgG fractions isolated from AT1-Ab-positive newborn rats induced significant vasoconstriction, similar to angiotensin II (AT1 receptor agonist) of the same concentration (contraction values, 0.44±0.05 g and 0.49±0.07 g, respectively). Furthermore, the vasoconstrictive response was markedly blocked (0.05±0.02 g, p<0.01 vs. IgG from immunized group offspring) by 10 μM losartan (AT1 receptor blocker). The IgG fractions from vehicle newborns demonstrated no vasoconstrictive effects, even at IgG concentrations of 5 μM.
As summarized in Figure 3C and Supplementary Video S1, within 8 min, a significant increase of intracellular Ca2+ level in cultured aortic smooth muscle cells (SMC) was apparent after stimulation with IgG fractions from the immunized group newborn rats (1 μM). Maximal change of fluorescence occurred after 1 min (p<0.01 vs. vehicle IgGs) and gradually decreased. Cellular preincubation with losartan (10 μM) for 20 min caused markedly reduced and delayed Ca2+ uptake, while vehicle IgGs (1 μM) had no effect on intracellular Ca2+ at any time. These in vitro experiments suggested that the circulating AT1-Abs in offspring might activate the AT1 receptor in a pathophysiological fashion.
Offspring body weight at age 0 and 3 weeks
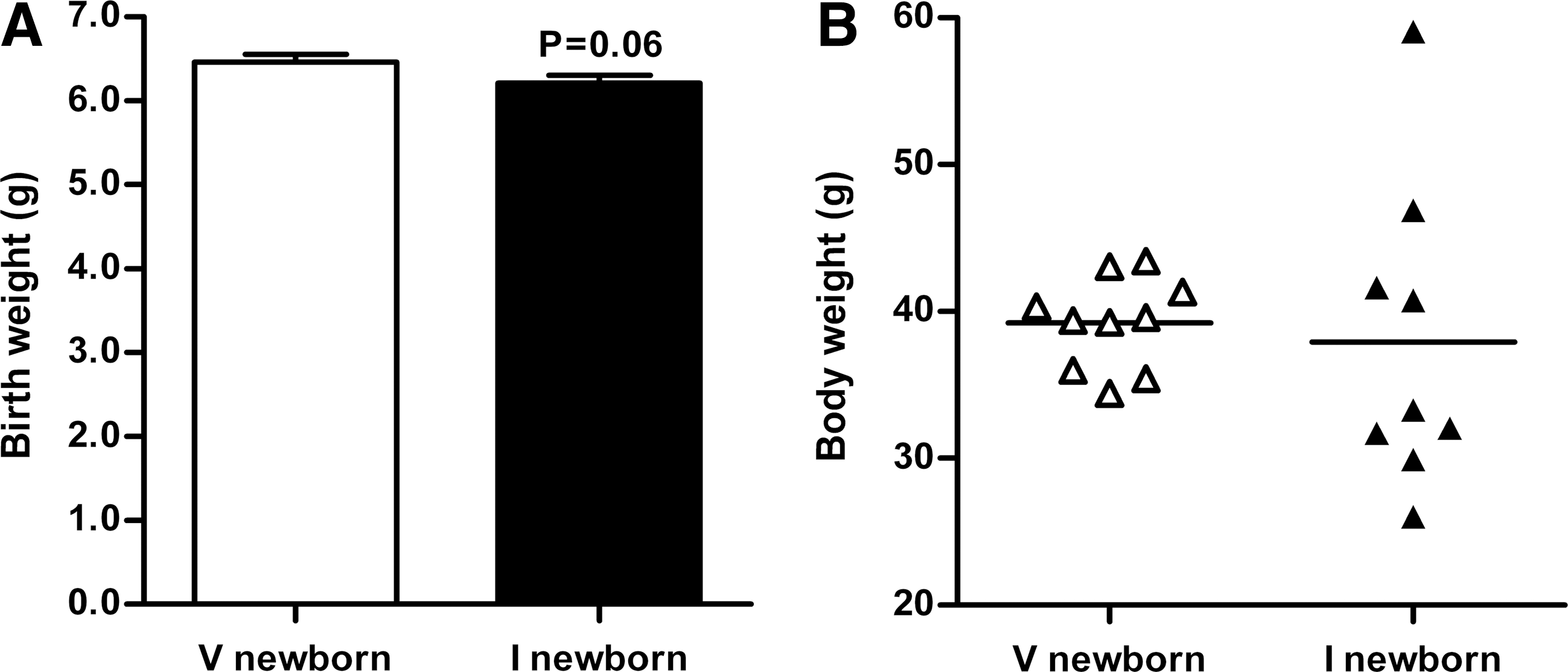
Immunized group newborn rats' birth weights tended to be slightly decreased compared with the vehicle group, but not statistically significant (p=0.06, 6.21±0.09 g vs. 6.46±0.09 g, Fig. 4A). Unexpectedly, the immunized group offspring body weight values demonstrated much greater variability compared with the vehicle group. Figure 4B and Supplementary Figure S2 revealed the body weights of both groups of 3-week-old offspring.
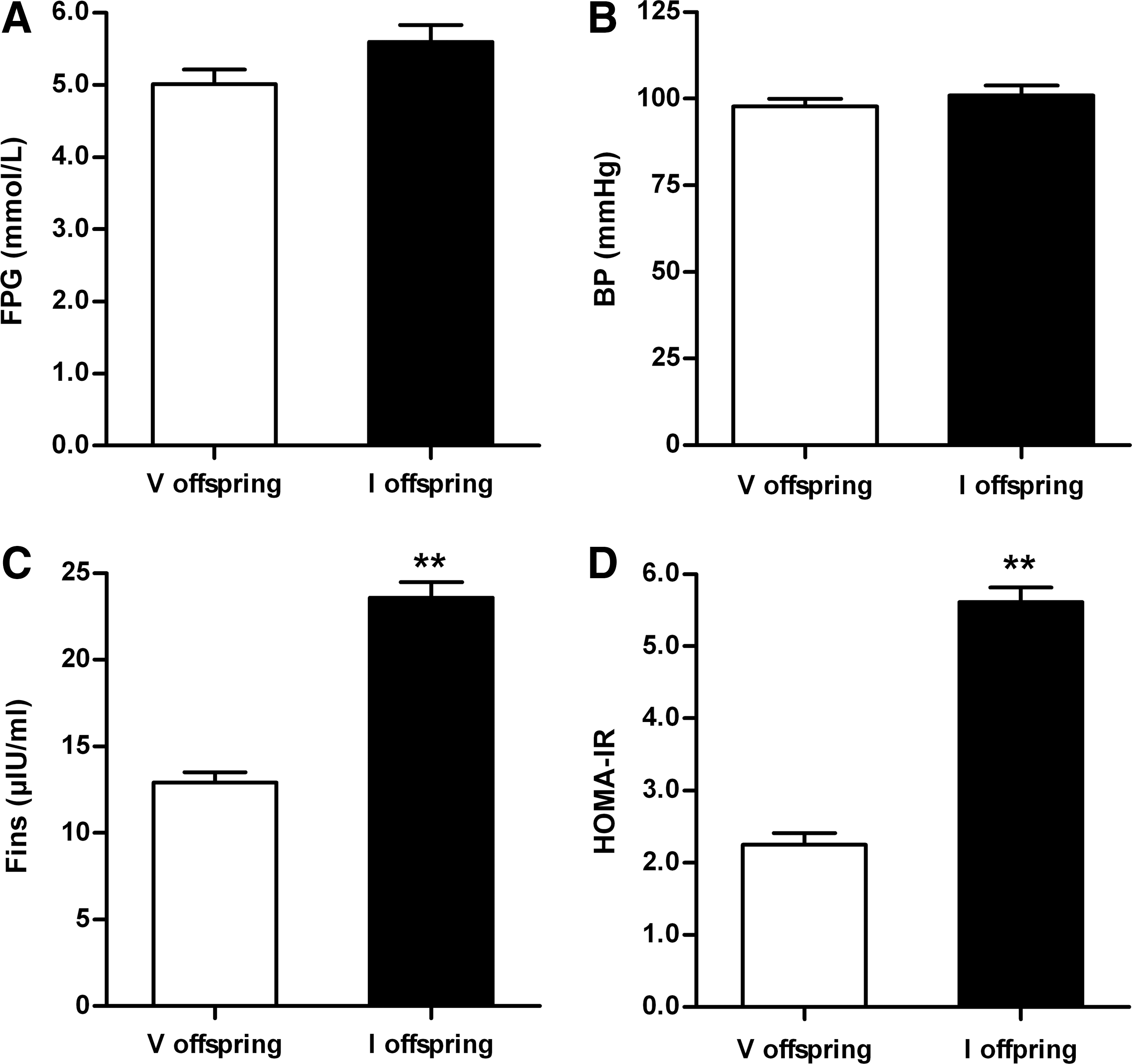
The presence of insulin resistance in 40-week-old immunized group offspring given a normal diet
The offspring of AT1-Ab-positive/negative pregnant rats were fed and raised in typical fashion until they reached 40 weeks of age. No significant difference was observed between the two groups with regard to fasting plasma glucose (FPG, 5.60±0.23 mM vs. 5.01±0.21 mM, p>0.05, Fig. 5A) or blood pressure (97.8±2.2 mmHg vs. 100.9±2.9 mmHg, p>0.05, Fig. 5B). However, fasting insulin in the offspring of AT1-Ab-positive mothers was greatly increased (23.6±0.9 μIU/ml vs. 12.9±0.6 μIU/ml, p<0.01, Fig. 5C), and the homeostasis model assessment for insulin resistance (HOMA-IR) was also elevated (5.60±0.20 vs. 2.30±0.16, p<0.01, Fig. 5D).
Development of MetS in adult offspring of the immunized group subject to high-sugar diet challenge
Results presented in Figure 5 demonstrated that the offspring of AT1-Ab-positive pregnant rats developed IR under normal living conditions. To determine whether an impaired insulin response in these animals may contribute to the development of MetS when the animals were fed with high-sugar food, additional experiments were performed. A high-sugar diet (20% sucrose) was administered to the offspring during postnatal weeks 40–48. FPG in the offspring of AT1-Ab-positive pregnant rats moderately increased at 44 weeks (6.14±0.27 mM vs. 5.56±0.11 mM, p=0.067) and markedly increased at 48 weeks (6.28±0.23 mM vs. the vehicle group 5.28±0.18 mM, p<0.01, Fig. 6A). Dyslipidemia accompanied the hyperglycemia observed. Compared with the vehicle group, an increased ratio of visceral fat weight to body weight (VF/BW, 8.1%±0.4% vs. 6.0%±0.5%, p<0.01, Fig. 6B), elevated triglycerides (TG, 6.69±0.38 mM vs. 2.57±0.31 mM, p<0.01, Fig. 6C), and decreased high-density lipoprotein cholesterol (HDL-C, 0.47±0.03 mM vs. 0.29±0.03 mM, p<0.05, Fig. 6D) were observed in the immunized group offspring, indicative of MetS.
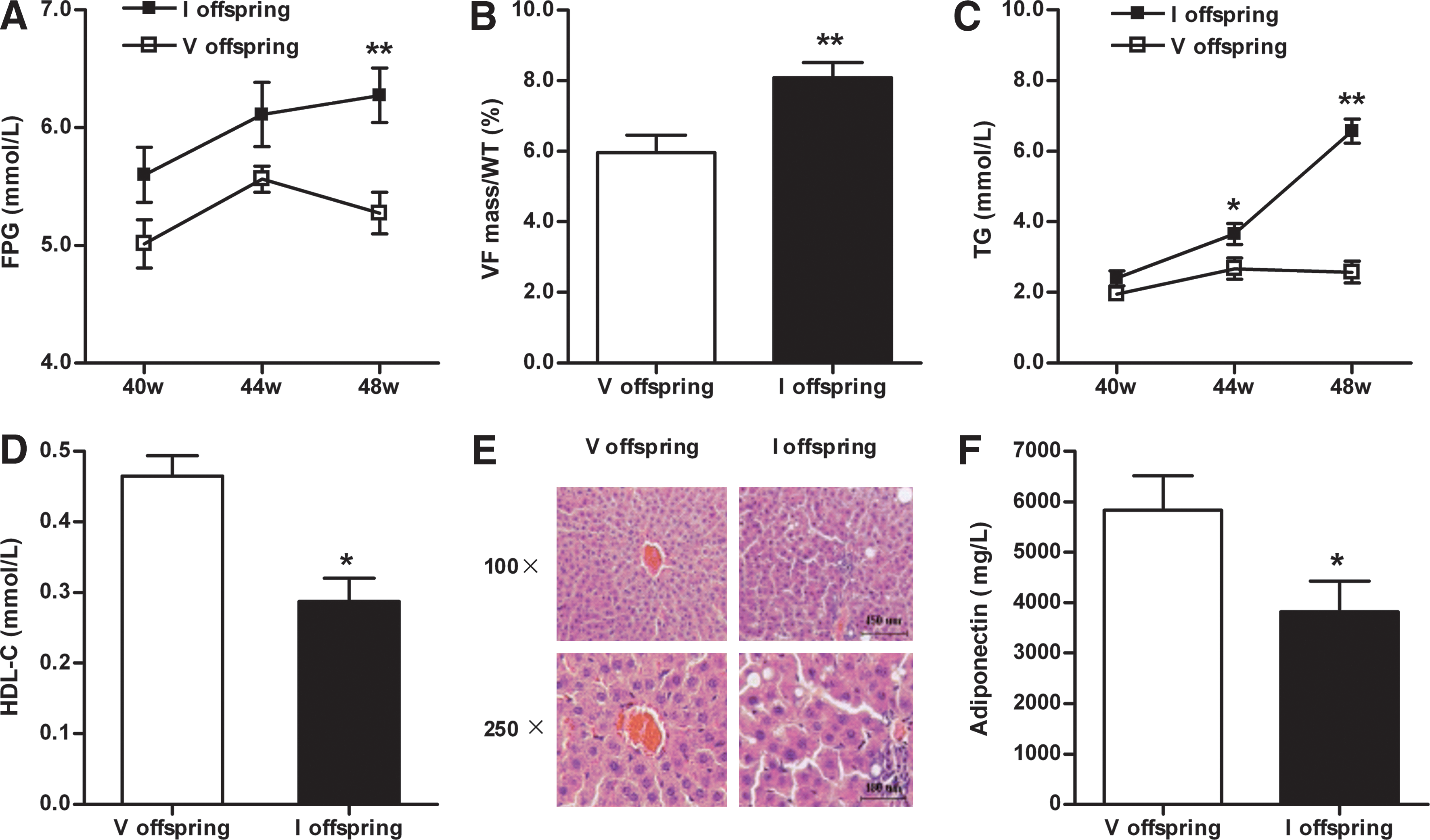
Additionally, hepatic fat accumulation in the rats was determined. In the immunized group offspring, numerous lipid droplets of various sizes were observed in the hepatocyte cytoplasm. In severe cases, hepatocyte nuclei were peripherally displaced, similar to adipocyte morphology. Portal region vascular dilatation, congestion, and significant inflammatory cell infiltration were observed (Fig. 6E). Vehicle offspring manifested grossly normal lobular architecture. Hepatocytes were arranged in a radial distribution surrounding the central vein. However, mild vascular dilatation, congestion, and inflammatory cell infiltration were observed in some portal regions (Fig. 6E).
Recent clinical and experimental studies have demonstrated that plasma levels of adiponectin, an adipokine with strong antidiabetic and cardioprotective properties, is significantly reduced in obesity and diabetes. To determine whether the offspring of AT1-Ab-positive pregnant rats may have altered adiponectin production, serum adiponectin levels were determined. As summarized in Figure 6F, serum adiponectin levels decreased significantly in the immunized group offspring compared with the vehicle group (3818±612 μg/L vs. 5837±678 μg/L, p<0.05, Fig. 6F).
Discussion
In the current study, we emphasize several important observations, the first of which is that AT1-Abs can be transferred from mother rats to offspring via the placenta and milk. Previous studies have demonstrated that IgG passes through the human placental barrier (yolk sac membrane in rats) and the neonatal enterocyte membrane (where milk is absorbed) through specific major histocompatibility complex class I–related Fc receptor (FcRn) binding. Among four rat IgG subclass types (IgG1, IgG2a, IgG2b, and IgG2c), IgG2a and IgG2b exerted greater affinity for FcRn, and crossed both the placental barrier and the neonatal enterocyte membrane more easily (8, 27). Our current study demonstrated that AT1-Abs in the sera of immunized female rats were predominantly IgG2b, with a very minor portion being IgG2a. These data provide credible theoretical support for the transference of AT1-Abs to offspring via the placenta and milk.
The rat yolk sac, similar to the human placenta, engages in maternal IgG transport by way of the syncytiotrophoblast and endothelial monolayer of villus, both of which express FcRn (17). Our current results illustrated that the sera of fetal rats born to antibody-positive pregnant rats contained high levels of AT1-Abs before labor onset; the vehicle group fetuses harbored no AT1-Abs. Such observations are identical to a recent clinical report (10) and suggest the ability of AT1-Abs to cross the rat placenta. Thus, since our current study provided only indirect evidence for antibody transfer, we sought specific morphological evidence identifying placental crossing of AT1-Ab. First, the immunogen, AT1R-ECII peptide, was conjugated to horseradish peroxidase (HRP). We located the target antibody in the yolk sac tissue via immunohistochemistry. Similar to general IgGs, AT1-Abs are distributed within the syncytiotrophoblast cell layer and vascular endothelium of the rat placenta in the immunized rat group, providing the first direct evidence for AT1-Abs transference via the placenta.
Maternal milk ingestion is another basic IgG transport pathway. Milk IgGs are specifically recognized by FcRn and transported across the newborn rat's intestinal epithelium into the circulation. Due to the difficulty associated with collection, we extracted maternal milk from the stomachs of newborn rats (7). ELISA showed AT1-Ab-positivity in the immunized group milk, making it potentially possible for antibodies to be transferred via milk ingestion. However, infant rat's stomach milk had a variety of confounding factors, including saliva, gastric juices, blood, and so on; therefore, the gastric contents might not completely represent pure mothers' milk composition. We then designed a cross-feeding experiment to confirm the transfer of AT1-Abs through milk (5). Due to the difficulty involved in distinguishing neonatal rats (the lack of hair and the tendency of mother rats to eat them when marked with ink or dye), we first separated the neonatal rats from both the immunized and vehicle groups by gender in order to facilitate the distinction among different subgroups. The exchange of the same sex newborns between two groups served as an experimental variable, and the un-exchanged offspring served as a control. AT1-Abs were present in the sera of the vehicle group neonatal rats cross-suckled by the immunized group mother rats after 1 week. Conversely, the AT1-Ab levels in the immunized group offspring markedly decreased after cross-feeding by the vehicle mother rats. Altogether, the data just provided fully demonstrated AT1-Ab transferability via milk, further demonstrating the transference mechanisms of AT1-Ab from mother to young.
A second important finding of the current study is that AT1-Abs in newborn rats remain biologically active and exert agonist-like effects through AT1 receptor activation. As seen in patients harboring autoantibodies (26, 33), the exogenous administration of AT1-Ab-positive sera IgGs obtained from baby rats induced significant thoracic aortic ring contractions and enhanced intracellular Ca2+ in cultured aortic SMC in vitro, similar to angiotensin II. These results suggested that early-life exposure to AT1-Abs (including the intrauterine and early postnatal stage) might be dangerous for offspring rats. Thus, excessive activation of the AT1 receptor might result in multiple pathophysiological effects, such as destroying vascular endothelial cells (35), inducing inflammation (4), promoting the secretion of aldosterone, and inhibiting the metabolic effects of insulin (24).
Third and the most clinically significant finding of the present study is that the AT1-Ab-positive offspring harbors several pathological alterations. First, birth weight is an important indicator reflecting fetal health. Abnormal birth weight shares a close relationship with multiple adult-onset diseases according to epidemiological data (2, 6). A nonstatistically significant downward trend was observed in AT1-Ab-positive pregnant rat fetal birth weights (p=0.06). Interestingly, there was distinctly greater body weight variability in the immunized group newborns compared with the vehicle group during weaning. Second, on reaching 40 weeks old (equivalent to human middle-aged life), having been fed a normal diet, AT1-Ab-positive pregnant rats' offspring exhibited normal FPG and blood pressure, but elevated serum insulin levels and increased HOMA-IR, indicating IR, while the vehicle group's metabolic parameters remained normal. Moreover, IR is a precursor to MetS, which may be due to genetic changes and/or adverse environmental factors (13, 18). The present study's data suggest that offspring born to AT1-Ab-positive mothers may have increased susceptibility to metabolic derangement by middle age. Finally, the offspring of AT1-Ab-positive pregnant rats were more sensitive to a high-sugar diet challenge. By 48 weeks postnatally, offspring of AT1-Ab-positive mothers demonstrated impaired fasting glucose, impaired glucose tolerance (Supplementary Fig. S3), increased visceral fat deposits, high TG, and low HDL-C, with normotension (data not shown), meeting the criteria for MetS criteria (1). In comparison, rats of the vehicle mothers subjected to the same conditions were of a nearly normal metabolic profile.
Direct/indirect impaired effects of AT1-Abs on the development of the placenta may explain the lower birth weight of rat offspring in the present study (4, 12, 32). AT1-Abs might contribute to uteroplacental ischemia/hypoxia, which is followed by increased fetoplacental vascular resistance and finally resulting in poor nutrient supply to the fetus (23). Then, for survival, the fetus would adapt its physiological development, including downregulating its growth, resetting the endocrine systems and metabolic homeostasis (28). Although beneficial for survival in utero, the fetus would be maladaptive and sensitive to metabolic and CVD when exposed to mismatched postnatal environments, such as catch-up growth, an atherogenic diet, and other risk factors.
Recently, both clinical and animal research suggest that the epigenetic modulation of transcription is partially responsible for the links between prenatal and postnatal influences and an increased risk of adult-onset diseases. Nutrient restriction in early life may directly induce epigenetic changes in growth and regulatory-related genes, which will cause permanent structural and functional alterations of important organs, such as the liver, pancreas, kidney, and the hypothalamic-pituitary-adrenal axis. Additionally, the phenotypic effects of epigenetic modifications during development may not manifest until later in life, especially when responding to environmental challenges (9, 14). Therefore, in our study, we speculated that the large degree of variation in offspring birth weights may be due to interactions between variations in the environment, adaptive epigenetics, and individual behavior. Furthermore, the mechanisms underlying the close association between early-life exposure to AT1-Abs and increased susceptibility to adult-onset MetS may be destroyed tissues and organs via various pathways (such as restricting nutrition, activating inflammatory cytokines) and/or epigenetic modulation of key genes. In addition, the oxidative stress and inflammation-related signal pathways (mitogen-activated protein kinase pathway, nuclear facor-κB signaling pathway, and so on) may play critical roles in the process (3, 4, 11, 25). However, the precise underlying mechanisms responsible are the subject of ongoing study.
In summary, we have demonstrated that AT1-Abs can be transferred from mother rats to their offspring and increase the offspring's risk of developing IR and MetS in middle and late life. These results strongly suggest that the AT1-Ab-positivity of mothers during pregnancy is a previously unrecognized “silent” risk factor for the development of MetS in their offspring. Since MetS has become one of the major public-health challenges worldwide, therapeutic interventions that block the adverse effects of AT1-Abs during pregnancy, or preventive actions (e.g., a low-sugar, low-calorie diet) taken by the descendants of AT1-Ab-positive pregnant women, may be novel strategies in the battle against MetS and its cardiovascular complications.
Study limitations
We used an active immunization rat model in the present study. As a classic method for autoimmune disease research, active immunization with a given autoantigen is easy to operate and widely applied. However, the levels of antibodies produced in the experimental animals are difficult to control, because the process depends on not only the dose of the antigen, but also the individual's immune status. The AT1-Ab levels in the mother rats in our study are a little higher than those in pre-eclamptic patients (36). We will use an alternative rat model that will be passively immunized with AT1-Ab in future studies. Furthermore, with a new model, various potential risk factors for MetS development, such as insulin, leptin, adiponectin, aldosterone, tumor necrosis factor-α, and glucocorticoid, should be dynamically monitored throughout the process. Another phenomenon which needs further exploration is that there is no hypertension in the offspring. It may be due to the complex regulation of blood pressure in vivo (such as depressor reflex) or our failure to capture the transient increase in blood pressure.
Materials and Methods
Animals
AT1-Ab-negative female (for establishing an active immunization model) and male (for mating) Wistar rats (8 weeks old) were used. All research procedures complied with the ‘‘Guiding Principles in the Use and Care of Animals'’ published by the National Institutes of Health (NIH Publication No. 85-23, Revised 1996) and were approved by the Institutional Animal Care and Use Committee of the Shanxi Medical University. The normal rat diet contained 280 g farina tritici, 125 g wheat bran, 375 g corn flour, 65 g soybean meal, 25 g fish meal, 2.5 g calcium lactate, 1.25 g probiotics, and 10 g salt per kilogram. The high-sugar diet was supplemented with 20% sucrose. The rat food was provided by the Experimental Animal Center of the Shanxi Medical University, China (production license No. SCXK[Jin] 2009-0001).
Immunization protocol
The peptide corresponding to the sequence of human AT1R-ECII was synthesized as previously reported (35). Twenty-four female Wistar rats were randomly assigned to two groups. The immunized group rats were actively immunized with synthetic AT1R-ECII peptide every 2 weeks as per the previous protocol (19). After 8 weeks, immunized female rats were mated with normal male rats (nonimmunized). During pregnancy and nursing periods, the female rats received monthly immunization. Thus, the mother rat models that possessed a relatively stable level of AT1-Ab would be established (35). Caudal vein blood was drawn before each immunization, and serum was collected to monitor antibody generation using the ELISA method as previously reported. The level of AT1-Ab was expressed as OD value, which is also called absorbance (A). A λ=lg(I 0/I), where I is the transmitted light intensity at a specified wavelength λ and I 0 is the incident light intensity. The absorbance is proportional to the concentration of the absorbing species in the sample and usually used to reflect the relative concentration of a sample. Acting as a reference, a control sample is essential. The positivity of the sample to AT1-Ab was also defined as P/N ≥2.1, while the negativity was defined as P/N ≤1.5 (35). The vehicle group rats were treated with a mixture of Freund's complete/incomplete adjuvant (without antigen) and saline in an identical manner.
Preparation of IgG and the subclasses identification
Total IgGs from the sera of immunized female rats containing maximal AT1-Abs were extracted by Mab Trap Kit (Amersham). Extraction purity was assessed by conventional SDS-PAGE. The IgGs from the vehicle group sera were obtained by an identical protocol. The purified IgGs subclasses were identified by ELISA methods. The dilution ratios of biotinylated goat anti-rat IgG1, 2a, 2b, and 2c antibodies (AbD Serotec) were 1:6000, 1:6000, 1:8000, and 1:8000, respectively.
HRP-AT1R-ECII probe preparation
AT1R-ECII antigen peptides were conjugated with HRP utilizing a Glue-B-type-activated HRP conjugation kit (Galaxy Bio). Briefly, the AT1R-ECII antigen peptide was diluted in double-distilled water to a concentration of 1 mg/ml. One hundred microliters of antigen solutions were added to the activated HRP (the HRP/antigen molecular molar ratio was maintained at around 30:1). The reaction body was regulated at pH 9.5 by a starting agent and incubated overnight at 4°C or at 37°C for 2 h. Then, 10 μl of stop solution was added and mixed for 15 min to cease the reaction. The mixtures were dialyzed for 48 h by dialysis bags with MW10000 interception. The water was changed six times to completely remove free AT1R-ECII peptides. Antigen molecules-HRP conjugates in liquid state were adjusted to pH 7.0 by 1 M phosphate buffer. This HRP-AT1R-ECII probe could specially recognize AT1-Abs, and was stored at −40°C until needed.
Immunohistochemistry
Location of AT1-Abs
Twenty-day pregnant (in labor) rats were intraperitoneally anesthetized with 10% chloral hydrate (3 ml/kg), and caesarean operations commenced. The placentas were removed and fully perfused with phosphate-buffered saline (PBS, pH 7.2) via the umbilical vein. The tissues were then fixed in 4% paraformaldehyde solution, and embedded in paraffin. Sections were cut at 8 μm and mounted onto glass slides. After dewaxing and hydrating, 3% H2O2 was added, and the samples were incubated for 10 min at room temperature. After three washings with PBS (5 min per time), the slides were subjected to high-pressure antigen retrieval (0.01 M sodium citrate buffer, pH 6.0, 270°C, 10 min), followed by another three PBS washings. The AT1R-ECII peptides conjugated to HRP (diluted in 5% PBS-Milk-Tween(PMT) solution [5% bovine serum albumin or 5% skimmed milk in PBS with 0.1% Tween-20], 1:100) were added and incubated overnight at 4°C. After another PBS washing, the slides were subjected to a diaminobenzidine (DAB) staining reagent.
Distribution of general IgGs
Placental samples were treated with rabbit anti-rat IgG antibody (diluted 1:200 in PMT; Bioss. Inc.) and incubated overnight at 4°C. Then, biotinylated goat anti-rabbit IgG secondary antibody was added and incubated at 37°C for 20 min. The process was repeated with HRP-labeled avidin under the same conditions. A PBS washing was performed following every step, concluding with a DAB staining reagent. The appearance of intracytoplasmic brown granules indicated the target antibody presence.
Milk collection and cross-feeding design
Mother rats' milk was collected from the stomachs of newborns after nursing as per a classically utilized technique (7). The AT1-Ab content of the milk was determined by ELISA via the same process as serum antibody detection.
In the cross-feeding experiment protocol, the immunized group and vehicle group mother rats that delivered on the same day were paired. Their newborns were separated by gender. In one pair of mother rats, male newborns were fed by their mother, and female newborns were exchanged for cross-feeding by the opposite group's mother. There were four subgroups: the vehicle group male newborn rats; the immunized group female newborn rats fed by the vehicle mother rats; the immunized group male newborn rats; and the vehicle group female newborn rats fed by the immunized mother rats. After 7 days of feeding, the serum AT1-Ab levels were compared among the subgroups. In another pair of mother rats, the female newborns were fed by their mother, and the male newborns were exchanged for cross-feeding.
Preparation of thoracic aorta rings
To determine whether the AT1-Ab present in offspring sera was biologically active, total AT1-Ab-positive/negative IgGs in the offspring sera were purified by the previous method. The effects of the IgGs on thoracic aorta and intracellular-free Ca2+ in SMC were observed in vitro. The vasoconstriction of thoracic aorta rings was performed as previously described (33).
Determination of intracellular-free Ca2+
Male Wistar rats were anesthetized with 10% chloral hydrate (3 ml/kg, intraperitoneal injection). The thoracic aortic SMC were isolated and primarily cultured. Intracellular-free Ca2+ concentration in SMC was monitored utilizing Fluo-3/AM, a common fluorescent Ca2+ indicator, in the dark (34). The changes in intracellular Ca2+ were defined as the difference between the fluorescence intensity at each recording time and the baseline fluorescence value. Ten cells in each group were randomly selected for image analysis.
Metabolism test
After a 12-h overnight fast, the levels of blood glucose, insulin, TG, and HDL-C were analyzed. Plasma glucose levels were determined by an automated glucose oxidase method (One Touch Ultra; LifeScan). Serum insulin was detected by a commercial double-antibody solid-phase radioimmunoassay (Coat-A-Count; DPC). Serum TG and HDL-C were assessed with an autoanalyzer (VetAce). HOMA-IR was calculated as (FPG [mM]×fasting insulin [μIU/ml]/22.5) (16). Both sides of the perirenal and epididymal fat weights were recorded as the visceral fat weight, and VF/BW was calculated. In addition, serum adiponectin was detected by an ELISA kit (R&D). Blood pressure was determined by tail-cuff plethysmography.
Statistical analysis
All data were described as mean±standard error of mean. Statistical analysis was performed with the SPSS 15.0 program. The t-test was used to compare two independent sample means, and one-way ANOVA was utilized for comparing means of more than two samples. The intracellular Ca2+ variations in different groups were compared by repeated-measures ANOVA. All pregnant rats and their offspring were randomly assigned to different experiments. A p-value <0.05 was considered statistically significant.
Footnotes
Acknowledgments
The authors are grateful to Lindsey Devillier for correcting the English spelling and grammar. This study was supported by grants from the Key Laboratory of Medical Electrophysiological Open Foundation of the Sichuan Province, KZ201110025023 from the Science and Technology Plan Project of the Beijing Municipal Education Commission, the NSFC 30900584, the NSFC 81070263, and the Basic Research Project of the Shanxi Province 2008011076-2.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.