
Abstract
Cysteine Supply in Different Parasitic Protozoa
GSH, glutathione.
Plasmodia rely exclusively on cysteine uptake from the host
Plasmodium falciparum, the causative agent of tropical malaria, multiplies in erythrocytes of its human host, thereby ingesting and degrading the host cell hemoglobin. However, hemoglobin degradation appears insufficient for its metabolic needs since the protein is a poor source of Met, Cys, Glu, and Gln and completely lacks Ile (78). In comparison to normal red blood cells, erythrocytes infected with P. falciparum have a dramatically increased permeability for Met, which is mainly attributable to new permeability pathways induced by the parasite in the host cell membrane (48). A wide range of neutral amino acids, including Cys, significantly compete with Met uptake providing a putative mechanism to supply the parasite with Cys. No genes for enzymes of the two cysteine biosynthetic pathways have been annotated in the genome of P. falciparum in accordance with earlier studies showing that different malarial parasites rely on exogenous cysteine supply (for reviews see refs. 175, 225).
Trypanosomatids obtain cysteine by synthesis and/or uptake
Trypanosomatids are a family of parasitic protozoa within the order Kinetoplastida (Table 1). The medically important trypanosomatids have digenetic life cycles that alternate between the human host and an insect vector.
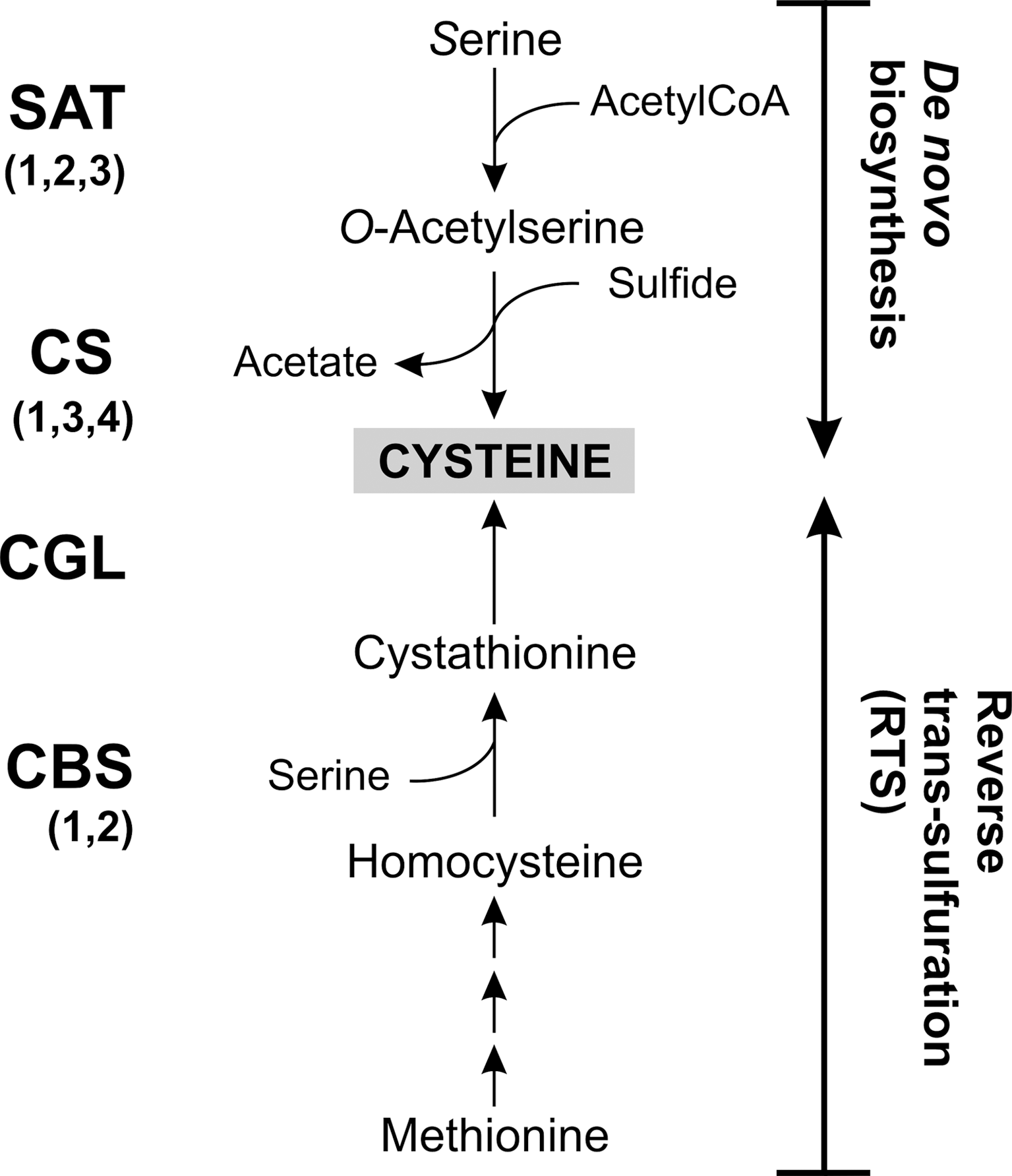
Pathogenic trypanosomatids differ in their mechanisms for cysteine supply. Leishmania major possesses enzymes for both de novo biosynthesis catalyzed by the concerted action of serine acetyl-CoA transferase (SAT) and cysteine synthase (CS) and by the RTS pathway composed of cystathionine-β-synthase (CBS) and cystathionine-γ-lyase (CGL) (Fig. 1) (277). L. major CBS has both CBS and CS activity. The source of sulfide for Cys biosynthesis is not clear since Leishmania lack genes for sulfur assimilation. This sulfate reduction occurs in Entamoeba as well as plants and bacteria. It has been proposed that the sulfide required for CS may be derived from 3-mercaptopyruvate by the action of mercaptopyruvate sulfurtransferase (MST; see Fig. 3 for the reaction) (276, 277). Cysteine transport into Leishmania can occur, but the rate is some 200-fold lower when compared to Trypanosoma brucei. Together with the high K m-value of 230 μM, this renders uptake unlikely as a main mechanism to supply the parasite with cysteine. Serine and methionine, but not cysteine, are essential for growth of promastigote Leishmania which underlines the importance of cysteine synthesis for this parasite (277).
Trypanosoma cruzi can also produce cysteine by de novo biosynthesis as well as by RTS from Met (177). Indeed, this parasite was the first protist shown to possess both pathways. The activity of CBS is about eight times higher in the insect epimastigote form than in the infectious amastigote and trypomastigote forms of the parasite, suggesting a stage-specific preference of the two synthetic pathways (177). Recently, an interactome analysis of tryparedoxin (Tpx) 1 (see below) in T. cruzi revealed CGL as an interacting protein (201). Thus, CGL is expressed in the parasite and may be redox regulated. T. cruzi epimastigotes possess a high-affinity and high-specificity transport system for L-cysteine (36). The maximum rate is, however, as low as that reported for Leishmania and the physiological role of cysteine uptake remains to be verified.
T. brucei encodes putative genes for CBS and CGL (277). However, cysteine is an essential nutrient (67), which suggests that the RTS pathway is inadequate to satisfy the cysteine requirements of African trypanosomes.
Microaerophilic protozoa developed individual pathways for their cysteine supply
Entamoeba histolytica, Trichomonas vaginalis, and Giardia lamblia (Table 1) share a variety of biological and biochemical peculiarities. These microaerophilic protozoa lack typical mitochondria, have an anaerobic carbohydrate metabolism, and use cysteine as the principal low-molecular-mass thiol (see next section) (71; for reviews see refs. 175, 244). However, the parasites employ divergent pathways for their cysteine supply. All three organisms lack the genes for enzymes of both the forward and RTS pathways and thus are unable to catalyze trans-sulfurations between Met and Cys (Fig. 1). Instead, E. histolytica is capable of sulfur-assimilatory de novo cysteine biosynthesis (146, 176). Three isoforms of SAT occur in E. histolytica, which show remarkable differences in their sensitivity to inhibition by cysteine (106). Overexpression of CS confers E. histolytica with a reduced sensitivity toward exogenous hydrogen peroxide, which is probably attributable to changes in membrane sensitivity (176), and suggests that cysteine biosynthesis plays a role in the oxidative stress defense. A recent metabolome analysis, however, indicates that the biosynthesis is not sufficient but cysteine uptake is essential (105).
De novo cysteine biosynthesis occurs also in T. vaginalis. Six genes encoding CSs have been identified, but SAT genes appear to be missing. Recombinant T. vaginalis CS1 accepts O-acetylserine (OAS), the common substrate of these enzymes, but also O-phosphoserine. This has led to the suggestion that O-phosphoserine, despite its high apparent K m-value, may be the physiological substrate of CS in T. vaginalis (267). The cellular level of CS is greatly reduced when the cells are grown in the presence of exogenous cysteine, suggesting that T. vaginalis can also salvage cysteine from its host (267).
Giardia duodenalis seems to rely exclusively on uptake for cysteine supply. Genes for both cysteine de novo synthesis and the RTS pathway are missing as it is the case in P. falciparum (175). This is in accordance with previous studies showing that the enzymatic activities are missing in homogenates of G. duodenalis and that exogenous cysteine is indispensable for both growth and protection against oxidative stress of the parasite (154).
The Microaerophilic Protozoan Parasites Use Cysteine as Key Antioxidant
Cultivation of E. histolytica requires high concentrations of cysteine in the medium that cannot be replaced by other thiols. Cysteine is required for growth, attachment, survival, and protection from oxidative stress (86). Recently, the cysteine metabolism of E. histolytica has been studied by metabolome and transcriptome approaches (104, 105). Cultivation of axenic trophozoites in the absence of cysteine results in nearly undetectable intracellular levels of cysteine and cystine. This strongly suggests that the endogenous biosynthesis is not sufficient but that the parasite greatly depends on cysteine uptake (105). Cysteine-depleted cells are highly sensitive toward paraquat and the intracellular levels of reactive oxygen species (ROS) increase three- to fourfold in accordance with cysteine being an important antioxidant. The metabolome analysis further revealed a dramatic accumulation of OAS and S-methylcysteine (SMC) and a decrease of S-adenosylmethionine (SAM) upon cysteine-depletion (Fig. 2A). SMC occurs in several legumes where it is considered to serve as a sulfur storage form (105). Surprisingly, depletion of cysteine does not result in an increased de novo synthesis from OAS. Instead, OAS is converted into SMC. The apparent failure to synthesize cysteine is not attributable to a lack of sulfide. It supports the conclusion that E. histolytica obtains cysteine primarily by uptake.
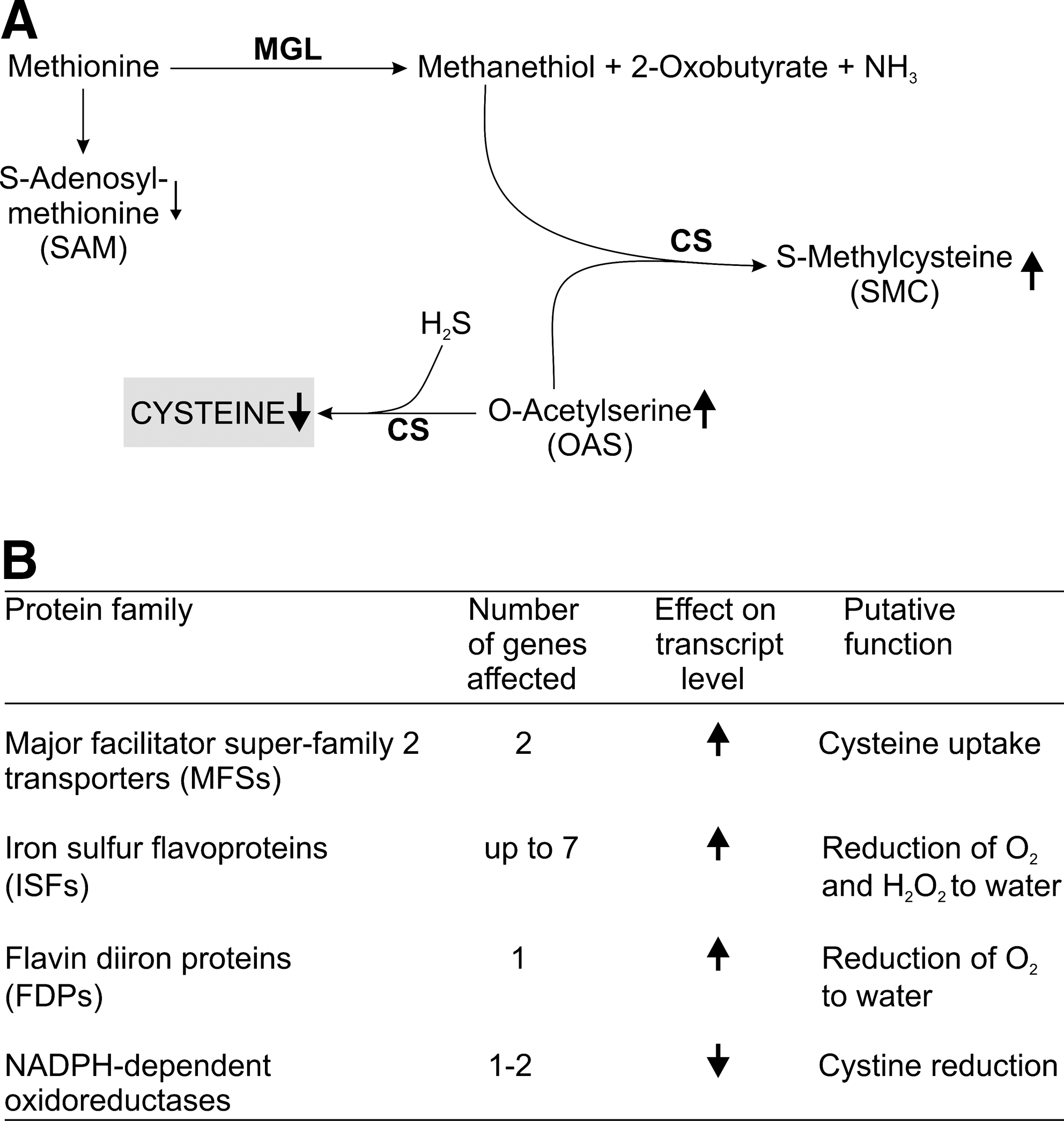
Cysteine deprivation affects the expression of a large number of genes (104), but the transcripts of most genes of the cysteine metabolism and of putative ROS- and reactive nitrogen species-detoxifying proteins are not significantly modulated in accordance with a previous study (257). This suggests that E. histolytica mainly relies on post-transcriptional mechanisms to adapt to oxidative stress. The most important changes caused by all three types of stresses are the induction of several major facilitator superfamily (MFS) transporters and iron–sulfur flavoproteins (ISFs) (104) (Fig. 2B). The E. histolytica genome encodes about 24 different genes for MFS proteins (104). It is tempting to speculate that under cysteine deprivation, the parasite may induce expression of cysteine or cystine transporters. ISFs are widely distributed in anaerobic prokaryotes where they may function in combating oxidative stress by reducing molecular oxygen and hydrogen peroxide (55). E. histolytica and T. vaginalis are the only known eukaryotes possessing ISF genes. Thus, an important in vivo role of cysteine is probably the regulation of different antioxidant enzymes. However, cysteine may also act as a direct antioxidant. This is corroborated by the fact that E. histolytica expresses an NADPH:flavin oxidoreductase and two other NADPH-dependent oxidoreductases, which all catalyze the reduction of cystine (32, 112). The expression of the latter enzymes is regulated by the availability of extracellular cysteine (112).
T. vaginalis possesses a full thioredoxin system, consisting of thioredoxin (Trx), thioredoxin reductase (TrxR), and Trx peroxidase, which probably represents a major antioxidant defense line (54, 139). The parasite does not produce GSH and lacks GSH metabolizing enzymes. Cysteine is present in the parasite at a concentration of about 600 μM and represents >70% of the total cellular thiols. Cysteine homeostasis in T. vaginalis is achieved by uptake and/or de novo synthesis as described above as well as by the catabolism of cysteine (Fig. 3). Cysteine can be degraded by the desulfurases methionine-γ-lyase (MGL) and cysteine desulfurase. The third mechanism is the concerted action of aspartate aminotransaminase (Asp-AT) and MST (266). T. vaginalis Asp-AT1 catalyzes the transamination of Asp and to a lesser extent of Cys. The observation that high levels of extracellular cysteine cause a three- to fourfold increase in MST activity suggests a role of this pathway in the redox homeostasis and antioxidant defense of T. vaginalis.
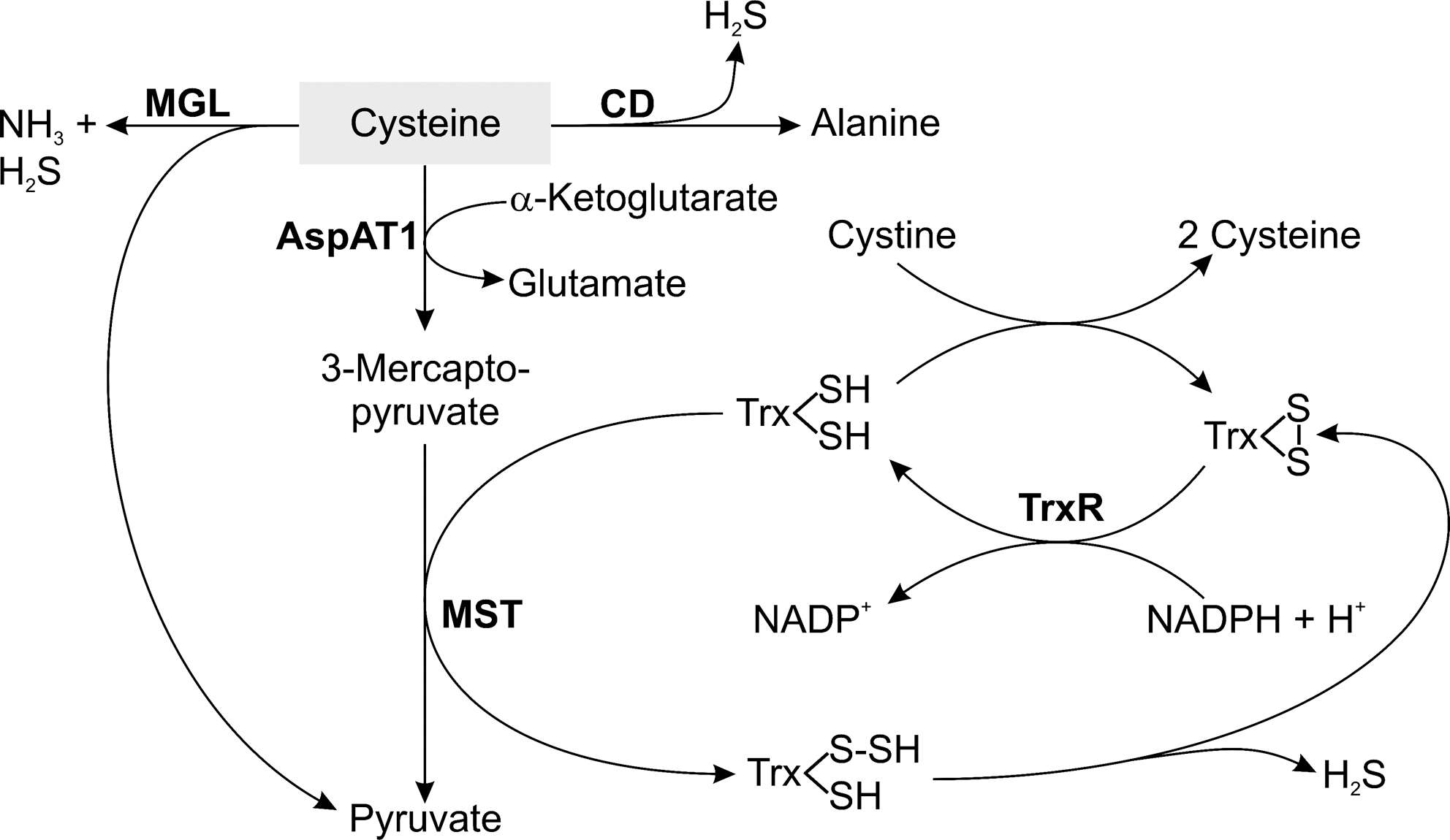
For most organisms, high concentrations of free cysteine are toxic and the cellular levels are maintained at 100–200 μM. In the presence of transition metal ions, cysteine rapidly undergoes autoxidation and the resulting hydroxyl radicals are highly toxic for cells and in particular for neurons (263). The use of cysteine as redox buffer in T. vaginalis may be an adaptation to its microaerophilic lifestyle. T. vaginalis is exposed to low levels of oxygen and does not produce ROS by aerobic respiration, which may increase its tolerance to comparably high cellular cysteine levels. It has even been suggested that the pronounced susceptibility of cysteine, compared with GSH, to autoxidation may serve to scavenge oxygen and thus protect the highly sensitive hydrogenosomal enzymes of the parasite from oxygen (266). The catabolism of cysteine is linked to the Trx system. Trx can act as sulfur acceptor in the reaction catalyzed by MST resulting in a cysteine persulfide. In the subsequent spontaneous reaction, sulfide is released and Trx is oxidized (Fig. 3). The dithiol form of Trx is then regenerated by TrxR. Persulfides are the source of sulfur for the biosynthesis of cofactors such as iron–sulfur clusters and lipoic acid or the sulfuration of tRNAs (167). The catabolism of cysteine via Asp-AT and MST may be a major source of sulfide also in these parasites. Another link between the cysteine and Trx systems is given by the fact that the parasite Trx spontaneously reduces cystine, suggesting that the Trx system is responsible for maintaining cysteine in its reduced state (54).
G. lamblia also contains cysteine as main low-molecular-mass thiol and GSH has been reported to be absent (30). The genome, however, encodes putative genes for the GSH biosynthetic enzymes γ-glutamylcysteine synthetase (GSH1) and glutathione synthetase (GSH2) (163). Recently, G. lamblia has been shown to possess a monothiol glutaredoxin (1-C-Grx) (202). These small redox proteins are characterized by a CGFS active site motif. They can dimerize by coordinating a [2Fe2S] cluster via the active site cysteines of each monomer together with the cysteines of two bound GSH molecules. 1-C-Grxs are involved in (probably the final step of) mitochondrial iron–sulfur cluster biosynthesis (143). The G. lamblia 1-C-Grx is localized in the mitosomes, highly reduced mitochondria-type organelles that harbor iron–sulfur cluster biosynthesis as only known function. The recombinant parasite 1-C-Grx has been shown to form a homodimeric iron–sulfur cluster complex that can be stabilized by GSH (202). It is therefore conceivable that G. lamblia contains at least low levels of GSH that may specifically be used in iron–sulfur cluster biogenesis.
G. lamblia possesses a flavin diiron protein (FDP) that efficiently reduces molecular oxygen to water (60). The primary function of the enzyme has been proposed to be the scavenging of oxygen, allowing the parasite to survive in the human intestine. A putative regulation of G. lamblia FDP expression by cysteine availability as it has been found in E. histolytica (see Fig. 2B) has not yet been studied. G. lamblia has an active Trx/TrxR system and encodes a gene for a putative Trx peroxidase. It is therefore likely that the parasite has a Trx-dependent peroxidase system as it is the case in T. vaginalis (54). G. lamblia TrxR has cystine reductase activity (31), which offers the possibility that the cysteine/cystine couple also acts directly in the redox homeostasis and antioxidant defense of the parasite.
Malarial Parasites Have a GSH Metabolism That Can Be Fueled by the Trx System
The antioxidant defense of P. falciparum is based on both a full GSH system as well as a Trx system (17, 59, 120, 169).
GSH synthesis and reduction in Plasmodia
P. falciparum is able to synthesize GSH (Fig. 4). The genes for GSH1 and GSH2 encode proteins with large insertions compared with the enzymes from other organisms, a phenomenon widely observed for plasmodial proteins (152, 160). Transcription of GSH1 is highest in the trophozoite state where hemoglobin degradation, and thus probably also the formation of ROS, is maximal (152). Recombinant P. falciparum GSH2 displays kinetic parameters comparable with the enzymes from other sources (160).
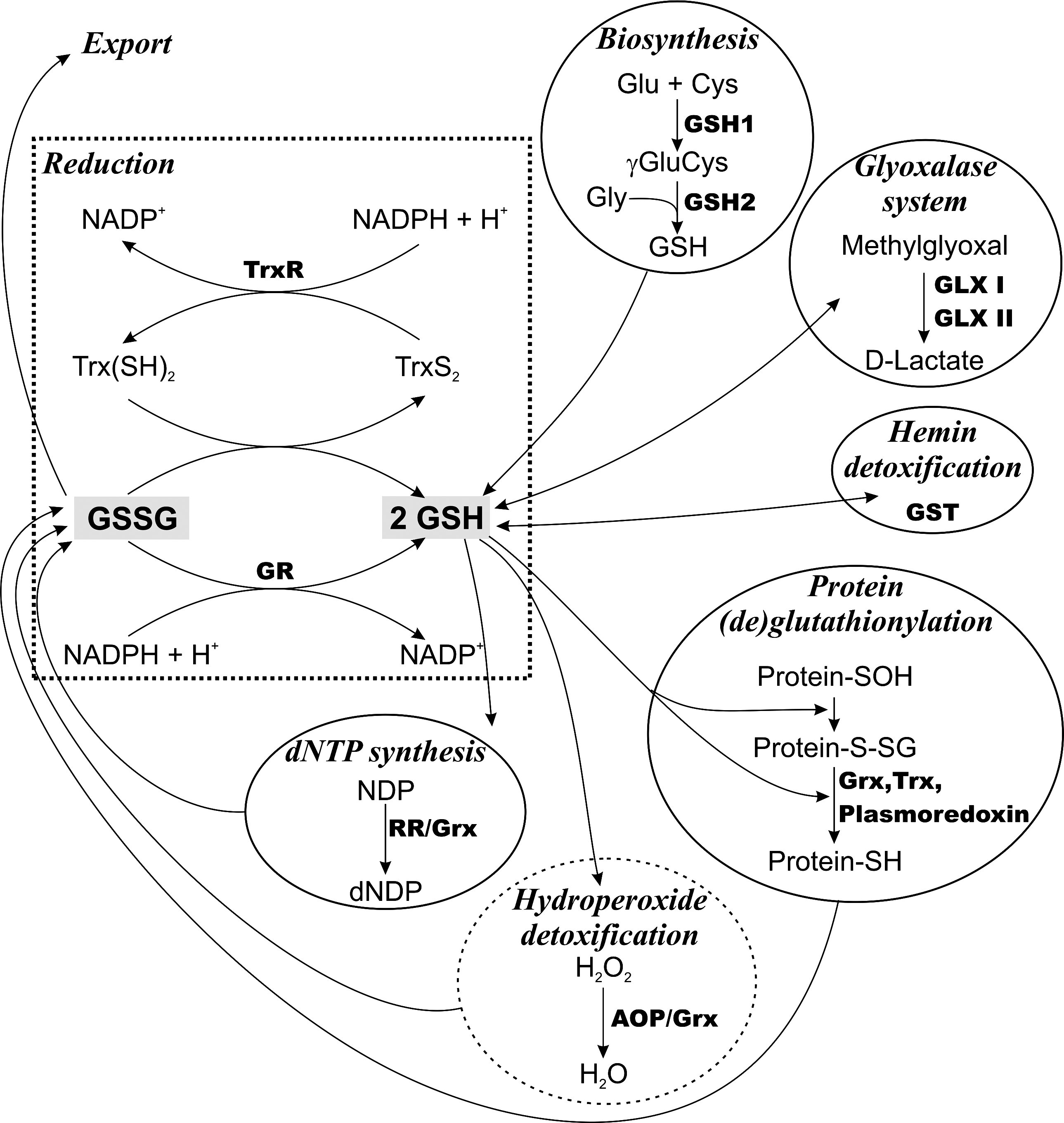
The total GSH content in erythrocytes infected with trophozoite P. falciparum is only halve that in noninfected red blood cells. This is probably mainly due to loss of glutathione disulfide (GSSG) from the infected erythrocytes, which occurs at a rate about 50-fold higher than in the case of uninfected cells (13, 153, 169). Decreased GSH levels were also reported for erythrocytes from patients infected with Plasmodium vivax (21). In other studies, the levels of GSH in intact P. falciparum–infected erythrocytes remained unchanged (for a review see ref. 17). In contrast, mice erythrocytes infected with Plasmodium berghei or Plasmodium vinckei have more than twice the GSH content found in uninfected erythrocytes (68, 232). This indicates that data obtained in rodent malaria models do not necessarily reflect the situation in the human pathogens (see below).
Treatment of P. falciparum–infected red blood cells with sub-lethal concentrations of buthionine sulfoximine (BSO), a specific inhibitor of GSH1, results in about 50% loss of the total GSH within 2.5 h. In contrast, an identical treatment of uninfected erythrocytes lowers the levels only slightly, 50% of GSH being lost after 4–5 days. This indicates that infection of erythrocytes by P. falciparum results in a strongly increased consumption of GSH (153). BSO inhibits the proliferation of cultured P. falciparum, suggesting that GSH biosynthesis is essential for parasite survival. However, the plasmodicidal effect of BSO is not due to specific inhibition of the parasite enzyme but to the altered metabolic needs for GSH in the infected cell (153).
By targeted gene disruption, GSH1-deficient mutants of the rodent malaria parasite P. berghei have been generated (256). These mutants are able to infect mice but show significantly reduced proliferation compared with wild-type parasites. Interestingly, the most dramatic effect is observed in the insect vector. Infection of Anopheles with GSH1-deficient P. berghei results in a reduced number of oocysts with aberrant morphology that do not produce sporozoites, thus interrupting transmission to the mammalian host.
P. falciparum glutathione reductase (GR) has first been isolated from infected erythrocytes (129). The gene has been cloned and overexpressed (74) and the crystal structure of the recombinant protein has been solved (215). Parasite and host cell GRs are studied as putative targets for antimalarial drug development (for reviews see refs. 111, 125). A recent thorough analysis of the compartmentation of the thiol redox proteins in P. falciparum revealed a dual subcellular localization of GR (120). Alternative translation initiation results in either the cytosolic or an apicoplast form of the enzyme. Together with several other redox proteins in this organelle, the data suggest the presence of an independent GSH-based antioxidant metabolism in this nonphotosynthetic plastid organelle in addition to the common cytosolic system.
Disruption of the GR gene in P. berghei only slightly affects proliferation of the asexual blood-stage parasites in cultured as well as in infected mice (33, 190). However, any attempt to generate mutants that lack both GR and GSH1 was not successful. This is evidence that the blood-stage parasites require either de novo GSH biosynthesis or the ability to regenerate GSH from GSSG. As observed for the GSH1-deficient parasites (256), GR-deficient P. berghei show the most severe phenotype in the insect stages. The lack of GR totally prevents formation of infective sporozoites. Taken together, in the case of the rodent malaria species either GSH biosynthesis or GSH reduction seems to be sufficient for parasite proliferation in the mammalian host. However, it is not known if this is also true for the human pathogens provided the difference in the GSH levels upon erythrocyte infection by individual malarial parasites described above. In addition, the P. falciparum Trx/TrxR couple has been shown to efficiently reduce GSSG and thus may replace GR activity especially at low substrate and enzyme concentrations (Fig. 4) (117).
GSH-dependent pathways in malarial parasites
Studies on the specific role(s) of GSH in P. falciparum revealed a number of GSH-dependent pathways (Fig. 4) (17, 119, 253). Upon infection, the glucose consumption of the red blood cell increases up to 75-fold (226). Both the host cell and the parasite rely on glycolysis for energy production. This is inevitably linked to the spontaneous formation of methylglyoxal, which is detoxified by conversion into D-lactate. The nearly ubiquitous glyoxalase (GLX) system, composed of GLX I and GLX II, uses GSH as a regenerated cosubstrate, not as reducing agent (for a recent review see ref. 283). Both the erythrocyte and the parasite harbor a functional GLX system (for a recent review see ref. 253).
P. falciparum has a single glutathione-S-transferase (GST) that, despite structural similarity to μ class enzymes, cannot be assigned to any known GST class (79). In addition to the common reactions catalyzed by these enzymes, in the presence of GSH the parasite GST can bind hemin (ferriprotoporphyrin IX) and thus contribute to the protection of Plasmodia toward this heme-derived toxic product (101, 142). The P. falciparum GST undergoes a dimer to tetramer transition that is accompanied by a loss of the ability to sequester hemin. GSH or GSSG prevents the tetramerization, suggesting that the protection of the parasite from hemin toxicity is independent of the redox status of the cell (141).
P. falciparum possesses a classical glutaredoxin (Grx) that catalyzes the GSH-dependent reduction of hydroxyethyl disulfide and can serve as electron donor for the synthesis of deoxyribonucleotides catalyzed by ribonucleotide reductase (RR) (203). A recent interactome analysis identified 17 target candidates for the P. falciparum Grx (235), but the specific physiological role of the protein remains to be elucidated since 10 of these interactions were also observed with Trx. A reaction usually catalyzed by Grxs is protein (de)glutathionylation. Formation of protein-GSH mixed disulfides is a reversible reaction allowing the positive or negative regulation of protein functions. In addition, the mechanism can protect thiol groups from over-oxidation and serve as a cellular storage form of GSH (reviewed in ref. 58). Nearly 500 proteins undergoing glutathionylation have been identified in P. falciparum suggesting a wide regulatory role of GSH in the parasite. Not only Grx but also Trx and the Plasmodium-specific plasmoredoxin can catalyze protein deglutathionylation in vitro (119).
Classical selenocysteine-containing GSH peroxidases as well as catalase are missing in P. falciparum. A total of five putative peroxidases have been described (for reviews see refs. 59, 173). The most important enzyme for hydrogen peroxide removal may be a 2-Cys-peroxiredoxin (2-Cys-Prx), which uses Trx as electron source (3, 118, 132). However, knockout parasites show only growth retardation when cultured in the presence of paraquat but proliferate normally in the absence of exogenous stressors (122). This may suggest that the 2-Cys-Prx is not essential for the detoxification of endogenously produced ROS at least under culture conditions (59). The nonselenium GSH peroxidase of P. falciparum also prefers Trx instead of GSH as reducing substrate. However, the low rate of hydrogen peroxide reduction in the presence of either thiol system points to a biological role distinct from antioxidant defense (238). The so-called antioxidant protein (AOP) of P. falciparum shows high sequence similarity to human Prx V and thus is an atypical 2-Cys-Prx. In vitro, AOP prefers Grx over Trx and accepts lipid-derived hydroperoxides as substrates although the overall peroxidase activity is low (173). The recent finding that the parasite Grx is located in the cytosol whereas AOP resides in the apicoplast (120) necessitates further studies of a possible in vivo interaction of both proteins. Overall, GSH seems to play only a minor role for hydroperoxide detoxification in malarial parasites (Fig. 4).
For yeast cells GSH has recently been shown to play an essential role in the cytosolic iron–sulfur cluster assembly (135). In Plasmodia, iron–sulfur cluster biogenesis has not yet been studied in detail. Several reactions in the plastid of the parasite require iron–sulfur clusters and an active assembly pathway has been suggested to be present in the organelle (134), suggesting another role of GSH in these parasites.
Trypanosomatid Parasites Have a Unique Trypanothione-Based Antioxidant Metabolism
Trypanosomatids show a large number of biochemical, morphological, and genetic peculiarities. The thiol and polyamine pathways are directly linked in these parasites, transforming their unique redox metabolism in one of the most distinctive features. The genome sequencing projects of T. brucei (19), T. cruzi (70), and L. major (110) revealed that trypanosomatids lack genes for GR and TrxR. While in most eukaryotic organisms the GSH/GR and Trx/TrxR systems maintain the intracellular thiol redox homeostasis, trypanosomatids possess a redox metabolism that is based on trypanothione [N1,N8-bis(glutathionyl)spermidine; T(SH)2] (72) and trypanothione reductase (TR), which keeps the dithiol in the reduced form (for reviews see refs. 73, 126). T(SH)2 is a much more efficient reducing agent than GSH although the redox potentials of these thiols are very similar (73) (Table 2). Since T(SH)2 is a dithiol, formation of an intramolecular disulfide is kinetically favored when compared with the intermolecular oxidation of two GSH molecules (85, 166). Moreover, the positive charge in the spermidine (Spd) bridge shifts the thiol pK-value to the physiological pH range (166) (Table 2), which renders T(SH)2 highly reactive in thiol-disulfide exchange reactions (85).
T(SH)2, trypanothione; TS2, trypanothione disulfide.
TR is the only enzyme that connects the NADPH- and the thiol-based redox systems in these parasites. The enzyme has been biochemically characterized in the insect parasite Crithidia fasciculata (224), as well as the pathogenic T. cruzi (127), Leishmania (56), and T. brucei (115, 236). TR is a representative of the flavin adenine dinucleotide (FAD)-cystine-oxidoreductases and shares many physical and chemical properties with GR. However, both enzymes have a mutually exclusive specificity toward their disulfide substrate (127, 224). Although Leishmania donovani promastigotes expressing 15% of wild-type TR activity grow normally in axenic culture, they are highly sensitive toward oxidative stress and exhibit a significantly impaired ability to survive inside activated macrophages (66, 248). In conditional knockout cell lines of bloodstream T. brucei, the TR activity could be lowered to less than 10% of wild-type cells (131). This caused a remarkable growth arrest and later the appearance of revertants. In addition, the TR-deficient parasites became highly sensitive toward exogenous hydrogen peroxide and were not able to infect mice (131). Therefore, TR plays an essential role in trypanosomatids and is considered to be an interesting drug target (130).
Trypanothione biosynthesis in different trypanosomatid organisms
The biosynthesis of trypanothione can be dissected into three parts: Spd, GSH, and trypanothione syntheses (Fig. 5). The pathways for GSH and Spd biosynthesis correspond to those in mammals. The last step is parasite specific and comprises the amide bond formation between the glycine residues of two GSH molecules and the primary amines of Spd.
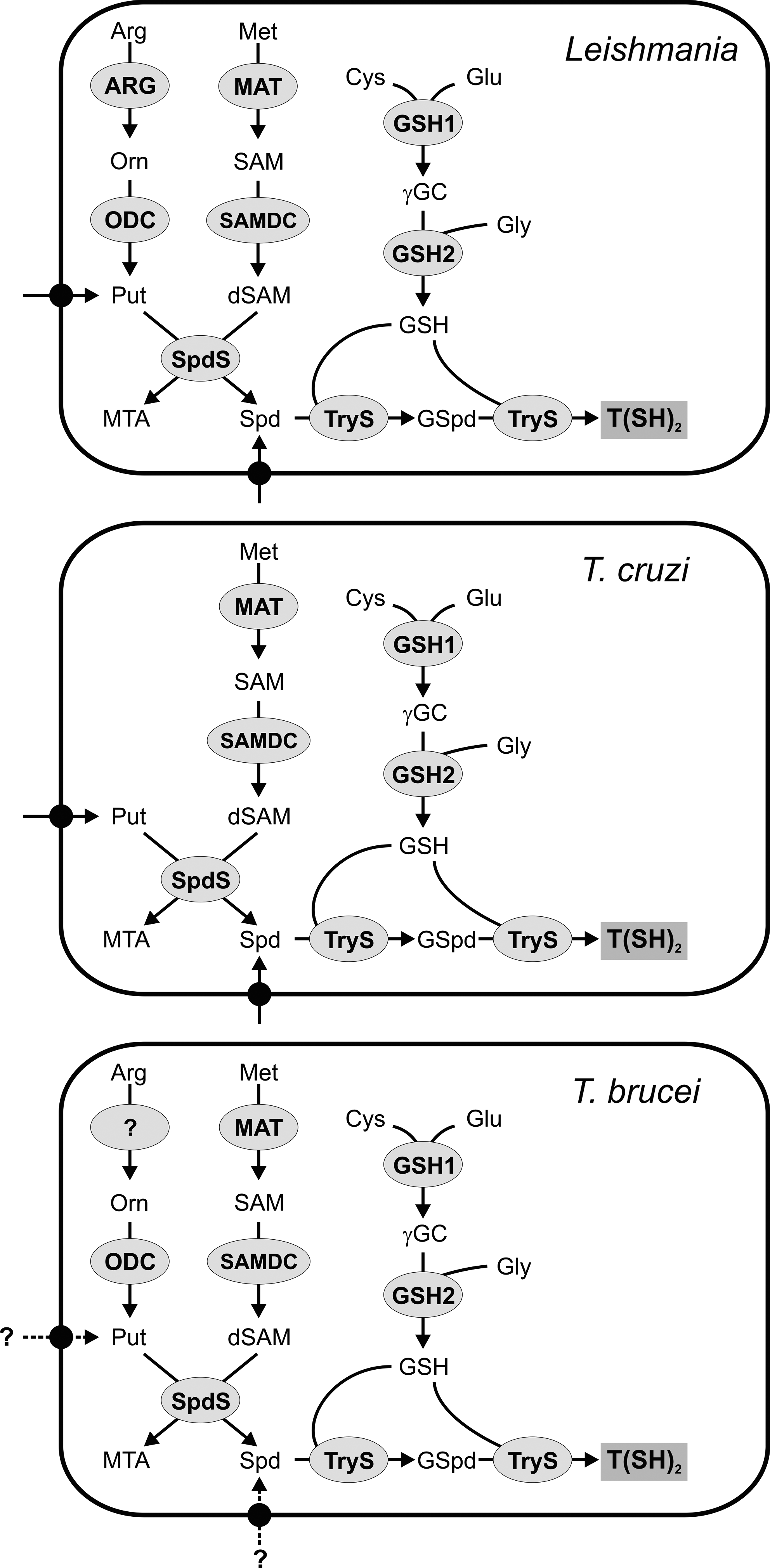
The polyamine pathway begins with the transformation of L-arginine into ornithine catalyzed by arginase (ARG, Fig. 5). The enzyme is present in Leishmania amazonensis (57), Leishmania mexicana (213), and L. major (209). ARG null mutants of promastigote L. mexicana and L. major fail to grow in semi-defined media (209, 213), but the proliferation defect can be rescued by supplementation with putrescine, Spd, or ornithine. The ARG null mutants retain the ability to infect Balb/c mice, although the animals show a slower progression of the lesions (82, 209). This suggests that the ornithine and/or polyamine levels inside the parasitophorous vacuole of the host cell are sufficient to sustain parasite growth. In trypanosomes, the origin of ornithine is still a matter of debate. Early studies were not able to show ARG activity in T. cruzi, T. conorhini, and T. mega cell extracts (35). A homolog of the functionally characterized leishmanial ARG is absent from the trypanosome genomes. However, a gene encoding a putative agmatinase/ARG-like protein is present in all trypanosomes and Leishmania (19, 70, 110). The functionality of the latter protein needs to be assessed before concluding that T. brucei is auxotrophic for ornithine.
L-Ornithine is converted into putrescine by ornithine decarboxylase (ODC, Fig. 5). This enzyme occurs in Leishmania (94) and T. brucei (197) but not in the American trypanosome (39, 103), which renders T. cruzi auxotrophic for putrescine. In human cells ODC undergoes a rapid turnover with half-lives of less than 1 h (84). In contrast, in T. brucei (83, 196) and Leishmania (94) the turnover of ODC is much slower (for a review see ref. 195). The different half-lives of the mammalian and T. brucei gambiense ODC form probably the basis for the specificity of the drug eflornitine (difluoromethylornithine [DFMO]), which acts as an irreversible inhibitor of ODC. The genetic disruption of the ODC gene is lethal for promastigote L. donovani (113) or procyclic T. brucei (140), but the parasites grow normally in putrescine-supplemented media. More recently, RNA interference in bloodstream T. brucei bloodstream showed that ODC is also essential in the infectious mammalian stage. Interestingly, wild-type proliferation is rescued by putrescine but not Spd supplementation (282). This suggests that putrescine is not only required for Spd production, but may play a so far unknown role for parasite survival. L. donovani ODC null mutants are severely compromised in their ability to establish an infection in Balb/c mice. The average parasite load in the liver and spleen of the animals is 3–6 orders of magnitude lower after infection with ODC knockout compared to wild-type parasites (26). This shows that L. donovani amastigotes are not able to scavenge enough putrescine or Spd from the host to properly sustain proliferation.
SAM is provided by methionine adenosyltransferase (MAT, Fig. 5) (for a review see ref. 210). The partially purified enzyme from T. brucei presented biphasic kinetics for ATP and methionine and, unlike the mammalian counterpart, was only poorly inhibited by SAM (284). Recombinant MATs from Leishmania infantum (208) and L. donovani (193) have been cloned and functionally expressed. It has been proposed that MAT may be implicated in the resistance of Leishmania against methotrexate (65, 93).
In the next step, SAM is decarboxylated by S-adenosylmethionine decarboxylase (SAMDC, Fig. 5), a self-processing enzyme with an active site pyruvoyl group. SAMDC catalyses the production of decarboxylated SAM (dSAM), which provides the aminopropyl group in the synthesis of Spd (69). SAMDC activity has been studied in T. brucei extracts (23, 243) and the L. donovani (212, 240) and T. cruzi (121, 194) enzymes have been cloned and characterized. In contrast to the short half-life of the mammalian isoform, the SAMDCs from Leishmania and T. brucei seem to turn over much more slowly (212, 222). Although trypanosomal SAMDCs are partially activated by putrescine (20, 194, 243), the enzyme requires the formation of a heterodimer with a catalytically inactive homolog to become fully active (275). Pharmacological inhibition of SAMDC by irreversible inhibitors can cure T. b. brucei and T. b. rhodesiense infections in mice (15, 22). Moreover, SAMDC null mutants of L. donovani revealed that the enzyme is an absolute prerequisite for cell growth in the absence of Spd (212).
The final step of Spd biosynthesis involves the transfer of an aminopropyl group from dSAM to putrescine catalyzed by spermidine synthase (SpdS, Fig. 5). The enzyme has only been biochemically characterized in T. brucei (24, 242). RNA interference-mediated silencing showed that SpdS is essential and that the proliferation of the parasites could not be recovered by Spd supplementation (242). More recently, using a different medium composition, another group demonstrated that the lethal phenotype caused by SpdS knockdown in bloodstream T. brucei can be overcome by supplementation with 100 μM Spd (282). Taking into account that the plasma Spd concentration is 300 nM (254), it is, however, likely that in vivo SpdS is essential. L. donovani SpdS null mutants are auxotrophic for Spd (87, 211). Moreover, the ability of L. donovani SpdS null mutants to infect mice is severely compromised. The average parasite load in the liver and spleen of infected animals is 2–3 orders of magnitude lower when compared to an infection with wild-type parasites (87), which makes SpdS an interesting drug target. Little is known about T. cruzi SpdS. The conversion of [3H]putrescine into Spd and spermine as well as of [3H]cadaverine into aminopropylcadaverine has been established in the American trypanosome (103). A 2.5Å X-ray structure of T. cruzi SpdS has been deposited at the protein data bank (3BWB).
The polyamine requirements of the parasites could also be satisfied by acquisition from the environment. T. cruzi is auxotrophic for polyamines. The epimastigote insect form possesses a specific putrescine transporter whose protein levels are regulated by the extracellular putrescine concentration (89, 138). A T. cruzi Spd–specific transporter, TcPAT12, was cloned and biochemically characterized (38). The presence of two different transport systems for Spd and putrescine was also observed in L. donovani and L. mexicana (16, 116). It was proposed that L. mexicana promastigotes and amastigotes possess different polyamine transporters that are adapted to work at different pH optima (16). A polyamine permease, LmPOT1, was cloned from L. major. The protein is present exclusively in promastigotes and can efficiently transport both putrescine and Spd (95). Finally, in T. brucei at least a low efficient transport system for Spd and/or putrescine should exist to explain the growth recovery of the ODC and SpdS knockdown mutants when cultured in polyamine-supplemented media (282). However, due to the low polyamine levels in the plasma it is very likely that under in vivo conditions bloodstream T. brucei rely on the biosynthesis of polyamines.
GSH in Trypanosoma and Leishmania is synthesized by two enzymes that are common to mammalian cells. The first step catalyzed by GSH1 (Fig. 5) ligates Cys and Glu to produce γ-glutamylcysteine. The reaction is considered to be the rate limiting step of GSH biosynthesis because overexpression of GSH1 in bloodstream T. brucei or Leishmania tarentolae is associated with a significant increase in the GSH and T(SH)2 levels (91, 223). The single-copy gene of T. brucei GSH1 has been cloned and the recombinant enzyme has been functionally characterized. As the mammalian enzyme, T. brucei GSH1 is inhibited by GSH (150). Different genetic strategies revealed that GSH1 is essential for T. brucei and L. infantum (107, 168). This is in agreement with the fact that BSO, a potent inhibitor of GSH1, can cure or prolong survival of mice infected with T. brucei (12).
The second step in the biosynthesis of GSH, namely, the linkage of γ-GC and glycine, is catalyzed by GSH2 (Fig. 5). In antimonite-resistant L. tarentolae, GSH2 expression is upregulated when compared to a sensitive strain (93). The crystal structure of T. brucei GSH2, solved at 3.15Å resolution, showed that the active sites of the trypanosomal and mammalian enzymes are almost identical (80). More recently, the biochemical characterization of recombinant T. cruzi GSH2 revealed a normal hyperbolic kinetic (178) in contrast to the cooperative binding of γ-GC, typical for the mammalian enzymes (155).
The last step in the biosynthesis of T(SH)2 is the conjugation of two GSH molecules with the N1- and N8-terminal amino groups of Spd (Fig. 5). Originally, the mechanism was studied in C. fasciculata which was shown to possess a glutathionylspermidine synthetase (GspdS) and a trypanothione synthetase (TryS) to catalyze the two consecutive steps (49, 246). However, since the TrySs of T. cruzi, T. brucei and L. major catalyze the conversion of GSH and Spd into T(SH)2 (181, 184, 185), it is likely that in the pathogenic trypanosomatids T(SH)2 biosynthesis is accomplished by a single enzyme. No GSpdS homolog has been found in the T. brucei genome and the one present in L. major is a pseudogene (184). Nevertheless, putative genes for GSpdS were described for T. cruzi and L. infantum though the functionality of the proteins remains to be proved (183). T. cruzi TryS displays a rather broad substrate specificity. It also accepts aminopropylcadaverine and spermine, thus producing homotrypanothione [Nl,N9-bis(glutathionyl)aminopropylcadaverine] and Nl,N12-bis(glutathionyl)spermine, respectively (10, 103, 185). Nonetheless, the preferred physiological product is T(SH)2. GSpdS and TryS are composed of a C-terminal synthetase domain and an N-terminal amidase-domain that can hydrolyze the conjugates to restore GSH and Spd (81, 181, 182). The paradox activity, rather than being a futile cycle, has been suggested to regulate the intracellular polyamine and/or GSH levels under specific conditions (27, 227). Despite some attempts to verify this hypothesis, the fact that the amidase activity is not required for the viability or infectivity of bloodstream T. brucei (280) leaves its physiological role an open question. T. brucei TryS is essential for parasite survival and has been validated as a drug target by both inverse genetic (9, 50) and chemical approaches (247).
Despite a lot of work, it is still not clear whether the initial conjugation of GSH occurs at the Nl- or N8-position of Spd. N8-GSpd is almost undetectable in crithidial extracts but in vitro studies showed that the k cat/K m ratio of C. fasciculata TryS for N8-GSpd is an order of magnitude higher than that for the Nl-isomer. This has led to the suggestion that both isomers are generated during the normal catalytic cycle of the enzyme (97). However, when taking into account that Escherichia coli GSpdS exclusively produces N1-GSpd (239) and that modifications of the 3-aminopropyl but not of the 4-aminobutyl substituent abrogate the activity (27), it is likely that the parasite TrySs also mainly generate N1-GSpd. This assumption is strengthened by the fact that N8-acetylspermidine is almost as efficiently used by T. cruzi TryS as Spd while the N1-acetyl derivative is only a poor substrate (10).
Trypanothione-dependent pathways in trypanosomatids
T(SH)2 spontaneously reduces dehydroascorbate (DHA) at least two orders of magnitude faster than GSH (128) (Fig. 6). T(SH)2 can also directly reduce GSSG and ovothiol disulfide (8, 165, 228). Furthermore, this unique dithiol displays much higher propensity than GSH to intercept NO in a dinitrosyl-trypanothionyl-iron complex that does not need to be sequestered by proteins, because it is harmless to the parasite even in its free form (25). T(SH)2 may be involved in the defense against toxic xenobiotics. In fact, the eukaryotic elongation factor 1B complex from L. major (259), localized to the endoplasmic reticulum (ER) surface, displays trypanothione-S-transferase activity as well as low peroxidase activity toward hydrophobic peroxides (259). Finally, T(SH)2 participates as a cofactor in the detoxification of reactive ketoaldehydes such as methylglyoxal. In contrast to the GSH-dependent mammalian GLXs, the trypanosomatid GLX I and GLX II enzymes are trypanothione-dependent (109, 258). T. cruzi (90), L. major (11, 258), and L. donovani (186, 187) possess a complete GLX system. In contrast, a GLX I gene is absent from the genome of African trypanosome (19) and GLX II does not seem to be essential for the proliferation of bloodstream and procyclic T. brucei (265). As recombinant T. brucei GLX II hydrolyzes thioesters that can be derived by GLX I-independent reactions, a function not related to methylglyoxal detoxification has been proposed (265).
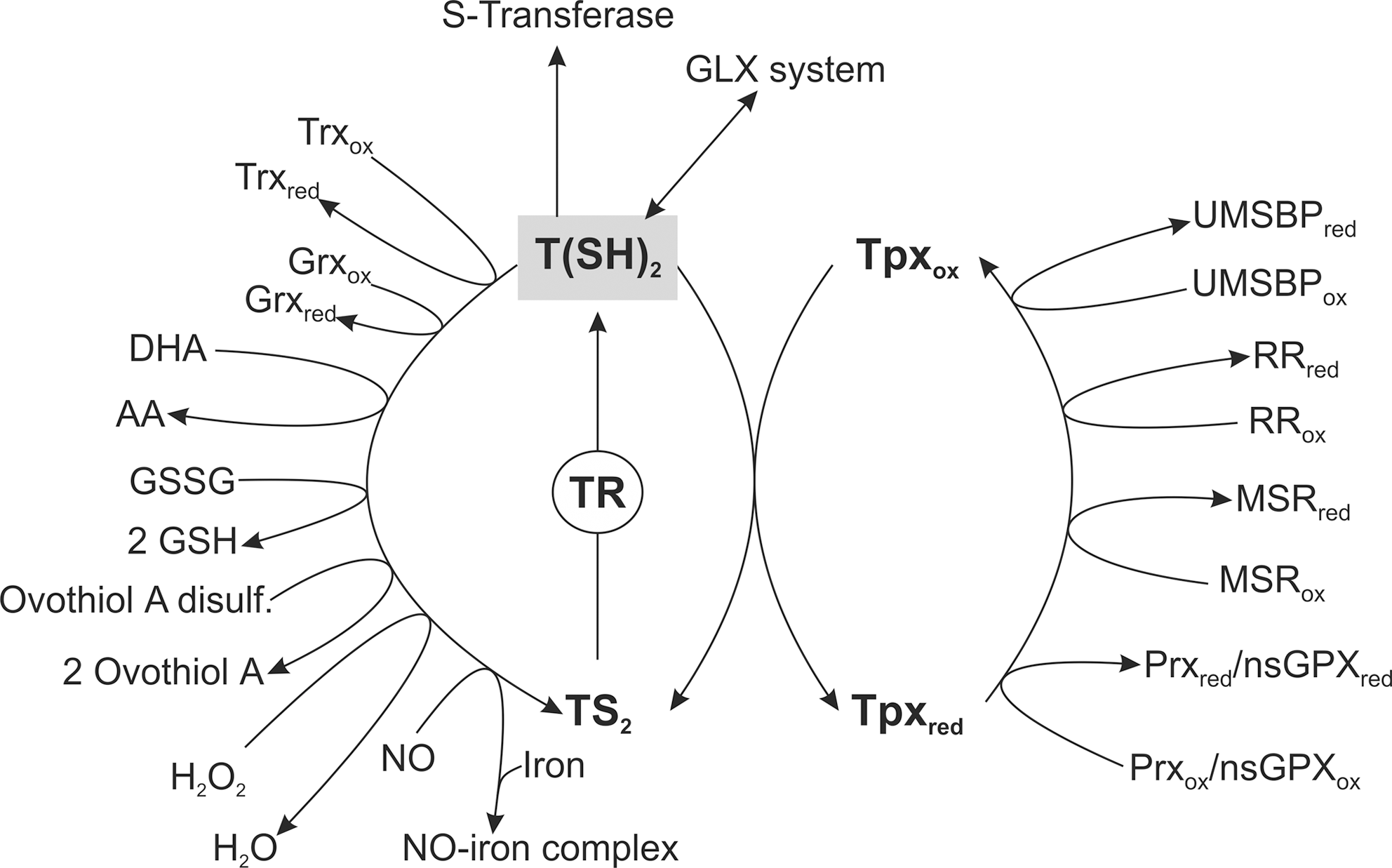
Many reactions of trypanothione are mediated by Tpx
Although at high concentrations T(SH)2 directly undergoes in a wide variety of chemical reactions, many of the reactions are accelerated by the presence of Tpx. The protein is a distant relative of the superfamily of Trx-type thiol disulfide oxidoreductases (42, 88, 148, 149). Tpxs and Trxs share the same core structure (5, 102) but show only 13% overall sequence identity. Moreover, the trypanosomatid-specific Tpxs have a WCPPCR active site motif [instead of the typical WCG(A)PC motif of Trxs] and with 16 kDa are substantially larger (162). Functionally, Tpxs share many properties with Trxs and Grxs (88, 149, 174, 231). In vitro Tpx can be reduced by mammalian TrxR. However, the enzyme is absent and T(SH)2 is the in vivo reducing agent. Both the redox potential and the thiol pK-value of Tpx are every similar to the respective values of T(SH)2 (Table 2) and the Tpx/T(SH)2 couple is the determining factor for the intracellular redox milieu of the parasites (206). Two types of Tpx occur in trypanosomatids. The first class includes the functionally characterized mitochondrial and cytosolic isoforms in L. infantum (mTpx and cTpx) (42) and T. brucei cTpx (149,206,245). The class II Tpxs are unlikely to function as normal Tpxs owing to a number of amino acid substitutions (41).
In T. brucei, downregulation of the cTpx expression affects cell growth (51, 268) and is accompanied by transiently elevated levels of low-molecular-mass thiols (51). The cTpx is also essential for L. infantum (214), but the mTpx homozygous knockout promastigotes do not show any growth retardation or morphological alterations when compared to wild-type parasites (41). Functionally, Tpx acts as a multipurpose oxidoreductase that transfers reducing equivalents from T(SH)2 to a variety of protein targets such as (i) RR, (ii) the universal minicircle sequence-binding protein (UMSBP), (iii) methionine sulfoxide reductase (MSR), and (iv) 2-Cys-Prxs and nonselenium glutathione peroxidase-type enzyme (nsGPX) (Fig. 6). T(SH)2, but not the monothiols GSH or GSpd, can serve as direct donor of reducing equivalents for T. brucei RR. Nevertheless, the presence of Tpx increases the RR activity almost 10 times, which suggests that in vivo the T(SH)2/Tpx couple acts as electron donor in the synthesis of DNA precursors (64). UMSBP is a zinc finger protein that plays a key role in the replication of mitochondrial DNA in trypanosomatids. The binding of UMSBP to the kinetoplast DNA is significantly affected by the redox state and Tpx has been shown to act as the reducing agent (161, 164, 179, 220). MSRs are involved in protein repair by catalyzing the reduction of methionine sulfoxide residues to methionine. Recently, T. brucei and T. cruzi have been shown to possess Tpx-dependent MSRs (7).
Trypanosomatids lack catalase and classical GPXs (reviewed in ref. 77). The parasite hydroperoxide metabolism is linked to T(SH)2, but trypanothione-dependent peroxidases were not detected (37, 96, 191, 192). Later it was shown that hydroperoxide detoxification occurs by a sophisticated cascade of reactions in which T(SH)2, TR, and Tpx play central roles as the carriers of reducing equivalents from NADPH onto two types of peroxidases, namely, 2-Cys-Prxs and nsGPXs (100, 174; for reviews see refs. 45, 124) (Fig. 6).
The genomes of T. brucei, T. cruzi, and L. major (19, 70, 110) encode multiple, almost identical copies for cytosolic 2-Cys-Prxs at one locus and on another chromosome, a single copy gene for the mitochondrial analog. The cytosolic and mitochondrial localization of the proteins has been experimentally verified in T. cruzi (274), T. brucei (245), L. amazonensis (144), and L. infantum (43). In Leishmania spp., one of the genes carries a glycosomal targeting signal, suggesting that the enzyme may also localize to this peroxisome-like organelle (45). Trypanosomatid 2-Cys-Prxs preferably reduce H2O2 and some of the enzymes are even inactivated by lipid-derived hydroperoxides (34, 40, 76). The 2-Cys-Prxs take also part in the decomposition of peroxynitrite (ONOO−) (6, 198, 249). In comparison to Tpx and T(SH)2, which react with ONOO− at rates that are within the range of typical thiols, the T. brucei and T. cruzi 2-Cys-Prxs reduce peroxynitrite 100 times faster (249; for a review see ref. 250).
In the infective form of T. brucei, the cytosolic, but not the mitochondrial, 2-Cys-Prx is crucial for parasite survival and protection against H2O2-mediated oxidative stress (268). However, T. cruzi epimastigotes overexpressing either the cytosolic or the mitochondrial isozyme are more resistant toward hydrogen peroxide, t-butyl hydroperoxide, and peroxynitrite (6, 198, 274). Due to the correlation between the 2-Cys-Prx protein levels in different T. cruzi strains and the parasitemia of infected mice, the enzymes are thought to be virulence factors (199). This hypothesis is strengthened by the fact that T. cruzi overexpressing the cytosolic 2-Cys-Prx can overcome the macrophage-derived cytotoxicity and display higher parasitemia and more prominent inflammatory tissue infiltrates in a mice model (6). In L. infantum, overexpression of the cytosolic and mitochondrial 2-Cys-Prx increases the resistance of promastigotes toward H2O2 and t-butyl hydroperoxide, respectively (43). Very recently, the mitochondrial 2-Cys-Prx has been shown to play an essential role for the infection of the mammalian host by Leishmania. Interestingly, the protein functions as a chaperon in a reaction independent of peroxidase activity (44). Moreover, the leishmanial Prxs seem to be implicated in drug resistance. Overexpression of the cytosolic enzyme in L. tarentolae increases resistance toward antimonials (281) and increased Prxs levels have been found in antimony unresponsive L. donovani field isolates (279).
The second class of peroxidases in trypanosomatids that obtain their reducing equivalents from the T(SH)2/Tpx couple are nsGPXs. A genomic locus encodes a cluster of two or three nearly identical nsGPXs and at a separate locus an additional open reading frame for a distantly related nsGPX. As proposed by Castro and Tomas (45), the two types of enzymes will be called nsGPXs-A and nsGPXs-B, respectively. The only partially characterized parasite nsGPX-B is the T. cruzi protein. It is localized in the ER. The recombinant protein has been reported to have low GSH-dependent peroxidase activity with linoleic acid and phosphatidylcholine hydroperoxides and not to accept Tpx as reducing agent (273). It has been hypothesized that the protein plays a role in preventing or minimizing lipid peroxidation within the ER.
In trypanosomes and Leishmania, the nsGPXs-A are almost identical proteins, the only differences are the absence/presence of mitochondrial and glycosomal targeting signals. In T. cruzi, the nsGPX-As are found in the cytosol and glycosomes (270). In T. brucei, a cell fractionation revealed nsGPX-As clearly in the mitochondrial and cytosolic fractions (216).
Like mammalian GPX4, the trypanosomal and leishmanial nsGPX-A proteins are monomers (123, 216) and are probably mainly responsible for lipid hydroperoxide detoxification (61, 269). RNA interference studies proved that the enzymes are essential in both bloodstream and procyclic T. brucei (216, 268). The lethal phenotype of bloodstream cells in which the genes for the cytosolic nsGPX-As have been deleted is abolished by supplementing the culture medium with α-tocopherol or Trolox (61). The main physiological role of the trypanosomatid nsGPX-As appears to be the prevention of lipid peroxidation as it is the case for mammalian GPX4 (219, 285).
T(SH)2 is also the physiological reducing agent for the parasite Trx and Grxs
Despite the lack of GRs and TrxRs, T. brucei, T. cruzi, and L. major encode genes for Trxs as well as Grxs (19, 70, 110). The Trx gene is transcribed in both bloodstream and procyclic T. brucei (207), but the protein was not detectable by Western blot analysis even not in cells that expressed an ectopic copy of the gene at a level 30-fold higher than the wild-type transcript (217). In addition, deletion of both trx alleles in bloodstream T. brucei did not result in a growth phenotype at least under culture conditions (217). In T. cruzi epimastigotes, Trx was also only detectable after enrichment by immunoprecipitation. Immunohistochemistry showed a diffuse cytoplasmic staining with extension to the flagellum (200). In vitro, T. brucei Trx is an electron donor for RR (207, 218), 2-Cys-Prxs (207), and nsGPXs (100) as well as for 1-C-Grx1 (75). T(SH)2 reduces the disulfide form of Trx, but the rate is at least 100 times slower than that of the reaction with Tpx (88, 218). Taking into account that all reactions observed for Trx are also accomplished by Tpx, which is present at a much higher intracellular concentration, the role of Trx in trypanosomatids remains to be elucidated.
Structurally, Grxs have been classified into three categories: (i) classical Grxs with the CPYC active site motif and Trx/Grx fold; (ii) GST-related proteins that possess Grx activities but have a structural organization similar to GSTs; (iii) 1-C-Grxs that contain a single cysteine in their active site (usually CGFS). Two dithiol Grxs have been characterized in T. brucei. Grx1 is a cytosolic protein, while Grx2 is probably located in the intermembrane space of the mitochondrion. Both proteins are reduced by T(SH)2 and strongly catalyze the dithiol/disulfide exchange between T(SH)2 and GSSG. While Grx2 is more active as protein disulfide reductase, Grx1 can form an iron–sulfur complex and prefers protein-GSH mixed disulfides as substrates (46). A dithiol Grx has also been characterized from T. cruzi. The protein seems to be involved in the dithiol/disulfide exchange between T(SH)2 and GSSG and is likely to take part in the reduction of protein-GSH mixed disulfides (157). Three 1-C-Grxs are encoded in the T. brucei, T. cruzi, and L. major genomes (19, 70, 110). The trypanosomatid 1-C-Grx1 and 1-C-Grx2 are single domain proteins with a predicted mitochondrial presequence, whereas 1-C-Grx3 is a Trx/1-C-Grx fusion protein with no evident targeting signal. In bloodstream and procyclic T. brucei, 1-C-Grx1 and 1-C-Grx3 are abundant proteins that present highest intracellular concentration when the parasites are grown to the stationary/starvation phase (52). Complementation studies in Grx5-deficient Saccharomyces cerevisiae targeting the T. brucei proteins into the mitochondria revealed that only 1-C-Grx1 modestly rescues the mutant phenotype (75). Moreover, 1-C-Grx1 can coordinate an iron–sulfur cluster using GSH as ligand and plays a crucial role in the iron and redox homeostasis of the parasites (52). The Tritryp genomes (19, 70, 110) also encode genes for putative GST-related Grxs [T. brucei (Tb927.7.3500), L. major (LmjF.14.1480), and T. cruzi (Tc00.1047053508265.10)], but none of the proteins has been characterized so far.
Some Trypanosomatid Parasites Contain Ovothiol A
Ovothiols are a group of 4-mercaptohistidines that were first discovered in marine invertebrate eggs giving them their trivial name (252). Ovothiol A stands for 1-methyl-4-mercaptohistidine (Fig. 7), and ovothiol B and C are the respective Nα-methyl and Nα,Nα-dimethyl derivatives. Upon fertilization, sea urchin eggs form a protective envelop that is cross-linked by dityrosyl residues, a reaction requiring H2O2 as extracellular oxidant. The eggs contain 5 mM ovothiol C, which reacts with H2O2, and the resulting disulphide is then reduced by GSH. Thus, ovothiol can act as a nonenzymatic GSH peroxidase system protecting the early embryo from oxidative stress (251).
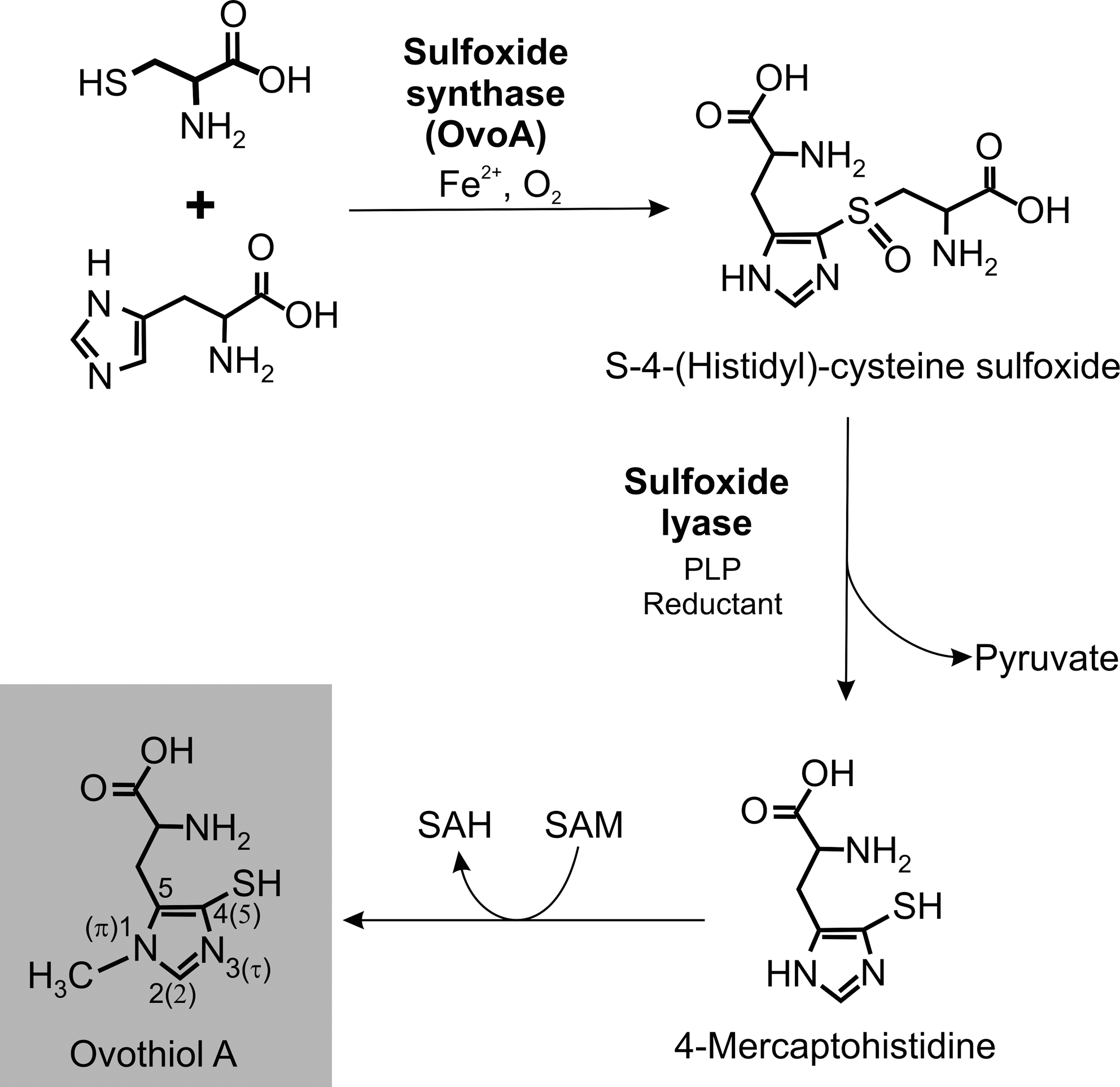
In 1994, Steenkamp and Spiess reported for the first time on the occurrence of ovothiol A in a trypanosomatid organism (229). Shortly after its detection in the insect parasite C. fasciculata, it became obvious that the aromatic thiol is also present in Leishmania (229) and other pathogenic trypanosomatids (8). The thorough analysis of its prevalence in different Leishmania species, T. cruzi and T. brucei, revealed ovothiol A in all insect stages with cellular concentrations ranging from >1 mM in Leishmania to at least 10-fold lower levels in trypanosomes (8). Interestingly, the mammalian stages of the parasites contain significantly less ovothiol A and bloodstream T. brucei as well as amastigote L. major lack detectable amounts.
The biosynthesis of ovothiol A has partially been unraveled (Fig. 7). In the presence of His, Cys, Fe2+, and pyridoxal phosphate, cell free extracts of C. fasciculata catalyze the formation of 4-mercaptohistidine (260). The methyl group of ovothiol A is most likely derived from SAM (228, 230). Very recently, putative genes for S-4-(histidyl)-cysteine sulfoxide synthase (OvoAs), the first enzyme of the biosynthetic pathway, have been identified in different eukaryotic kingdoms and bacterial phyla. The recombinant enzymes from the γ-proteobacterium Erwinia tasmaniensis as well as from T. cruzi catalyze—although with modest efficiency—the iron-dependent formation of OvoA from His and Cys (29). The enzymes catalyzing the following steps, namely, the cleavage into 4-mercaptohistidine and the final methylation still await their characterization.
The physiological role of ovothiol A in trypanosomatids is not known. With a thiol pK-value of 1.42 (264), ovothiol is in the thiolate state throughout the physiological pH range. Although ovothiols can act as nonenzymatic scavengers of hydrogen peroxide, the reaction is less efficient than that by T(SH)2 (8). In addition, the parasites possess efficient trypanothione-dependent peroxidases (see above). These findings render ovothiol A unlikely to play a major role in the cellular H2O2 metabolism of trypanosomatids. The disulphide form of ovothiol is not a substrate of TR but is readily reduced in the presence of trace amounts of T(SH)2, suggesting that ovothiol A is kept reduced by the spontaneous reaction with the dithiol (8). This is corroborated by the fact that the redox potential of ovothiol is −92 mV (264) and thus 150 mV more positive than that of trypanothione (−242 mV) (73). Leishmania as well as T. cruzi proliferate within macrophages and have to deal with the innate immune system of their mammalian host. In mice, expression of inducible nitric oxide synthase is essential for the elimination of L. donovani (172), suggesting an important role for NO in the elimination of intracellular trypanosomatids. Ovothiols are efficient radical scavengers. Ovothiol A decomposes S-nitrosoglutathione with a second-order rate constant three orders of magnitude higher compared with GSH. This has led to the suggestion that ovothiol A in synergy with T(SH)2 provides the parasites with an efficient system for the nonenzymatic decomposition of nitrosothiols (261). Since the mammalian stages of the parasites contain only very low levels of ovothiol (8), the physiological significance of this reaction remains to be elucidated. In addition, as described above, T(SH)2 itself can sequester NO in form of a harmless and stable iron complex (25).
Metazoan Parasites Have GSH-Based Redox Systems
Filarial worms secrete several GSH-dependent enzymes
The antioxidant defense in filarial nematodes such as Wucheria bancrofti, Brugia malayi, and Onchocerca volvulus largely corresponds to that in higher organisms (221). The main low-molecular-mass thiol is GSH (Table 1). The GSH1 and GR from O. volvulus exert structural and kinetic properties comparable to those of the enzymes from other origins (151, 170).
Nematodes possess superoxide dismutases, nsGPXs, GSTs, and Trx. As a peculiarity, these redox proteins, together with other proteins, are partially secreted as a potential immune evasion strategy of these parasites (18, 99, 136, 221). The nsGPX of Brugia pahangi is glycosylated. It is specifically expressed after infection of the mammalian host and represents the major surface protein of adult lymphatic filarial parasites (53, 241). The recombinant enzyme reduces lipid-derived hydroperoxides with GSH as electron donor and is inactive with a bacterial Trx system as reducing system (241). The GSTs of nematodes are a focus of recent antifilarial drug development approaches (14).
The GSH and Trx systems of flatworms are linked by a single reductase
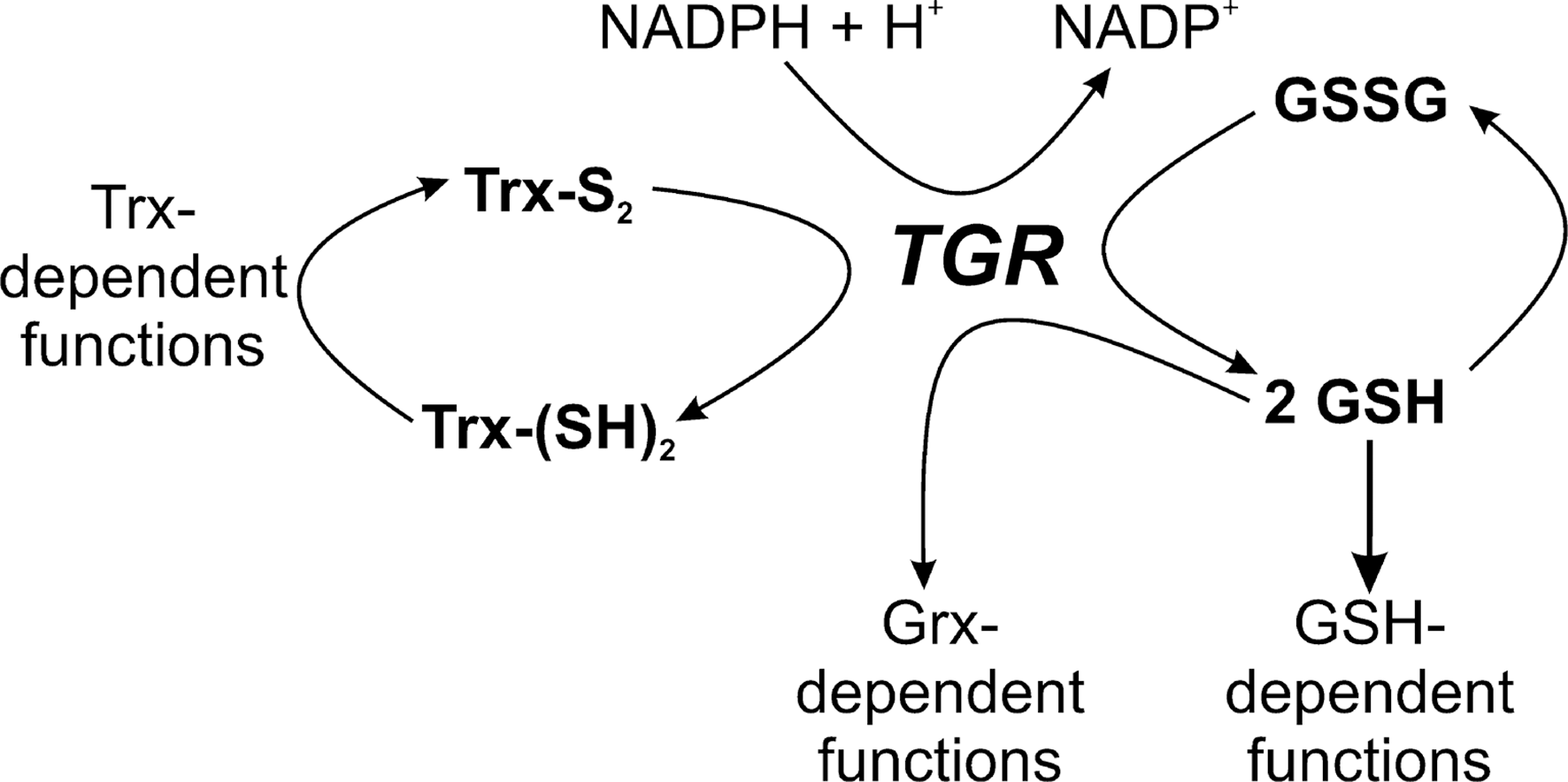
In platyhelminths (flatworms), to which belong Schistosoma, Echinococci, and Taenia, the causative agents of schistosomiasis, hydatic disease, and cysticercosis, respectively, cellular redox homeostasis and antioxidant defense are based on both GSH and Trx systems. However, parasitic platyhelminths—but not free living flatworms (180)—lack conventional GRs and TrxRs. Instead, the parasites rely exclusively on a single enzyme that provides reducing equivalents for both pathways (4). This thioredoxin glutathione reductase (TGR) is a fusion protein composed of an N-terminal Grx domain that is linked with a TrxR/GR-type NADPH-oxidoreductase. TGR has GR, TrxR, and Grx activities (Fig. 8) (4). In mammals this type of oxidoreductase is specifically found in testes (237). As mammalian TrxRs, TGRs are selenoproteins with a C-terminal Gly-Cys-SeCys-Gly motif. The parasite TGR occurs in the cytosol and mitochondria of the worms (2, 28, 92). It is essential for Schistosoma survival and is currently investigated as a target for specific antischistosomal chemotherapy (137).
In parasitic helminths, GSH probably plays an important role in the removal of heavy metals. Recently, a Schistosoma mansoni phytochelatin synthase (PCS) has been characterized (205). The enzyme catalyzes the conversion of GSH into phytochelatines. These oligopeptides with the general formula (γ-GC)n-Gly (n=2–11) efficiently sequester heavy metals. PCS occurs in all mammalian stages of S. mansoni and its expression increases in response to the presence of heavy metals. It has been hypothesized that the enzyme plays a role in the detoxification of iron generated by the breakdown of host hemoglobin. Three PCS transcripts are produced in S. mansoni by alternative splicing, probably generating two cytosolic and a mitochondrial version of the enzyme (205). PCS genes exist also in nematodes such as B. malayi as well as many other organisms but not in mammals.
S. mansoni possesses a selenocysteine-containing GPX that catalyzes the GSH-dependent reduction of hydrogen peroxide (159). Adult worms express the highest activity, which renders them more resistant to oxidative stress compared with other developmental stages. The physiological function of the parasite enzyme is probably that of a phospholipid hydroperoxide GPX (156).
Ascorbate as an Antioxidant in Parasitic Organisms
Ascorbate can directly metabolize ROS, maintain α-tocopherol (vitamin E) in its reduced state, and mediate the electron transfer to ascorbate-dependent peroxidases (272). Most eukaryotes can synthesize ascorbate. Among mammals, only humans, some other primates, and guinea pigs have lost this ability because they lack an active gulonolactone oxidase that catalyzes the final step of mammalian ascorbate biosynthesis. Few studies have been published on the role of ascorbate in parasitic organisms. Mouse erythrocytes infected with the rodent malarial parasite P. vinckei show an increased uptake of ascorbate compared with uninfected cells (234), whereas both normal and infected cells rapidly take up DHA and reduce it to ascorbate. Parasitized but not control cells release most of the ascorbate formed that could in part explain the higher plasma levels of ascorbate found in infected animals (108). Mice erythrocytes infected with P. vinckei contain higher levels of ascorbate when compared with uninfected cells, and it has been suggested that in late stage infections the ascorbate content and redox state is altered to keep vitamin E reduced and thus to protect the cell from hemolysis (233). Nevertheless, the physiological significance of ascorbate in malarial infection is not clear. Knockout mice that are unable to synthesize ascorbate develop the same parasitemia upon infection with the rodent parasite P. berghei as wild-type mice (98). In the case of the human malarial parasite P. falciparum, ascorbate has been even reported to be destructive to different intraerythrocytic stages (158).
In extracts of T. vaginalis ascorbate peroxidase activity has been measured, but the protein has not been characterized (188). Trophozoites of E. histolytica require cysteine and ascorbate in the medium for optimal attachment and growth (86), but the underlying molecular mechanism has not been revealed. B. malayi depends on exogenous vitamin C for larval molting and it has been suggested that the filarial parasite does not synthesize ascorbate. During larval development, ascorbic acid does not seem to act as a general antioxidant but to play a specific role (204). A distinct function of ascorbate or the (in)ability for biosynthesis of ascorbate in these parasites has not yet been reported.
Ascorbate supply and metabolism in trypanosomatids
Trypanosomatids have the capacity to synthesize vitamin C (Fig. 9). T. brucei arabinonolactone oxidase (ALO) and T. cruzi galactonolactone oxidase (GAL) share 60% of all residues (147, 272). The enzymes are localized in the glycosomes, unique peroxisome-like organelles that harbor the first seven enzymes of glycolysis as well as ether lipid biosynthesis and some other pathways in trypanosomatids (272). The so far uncharacterized Leishmania ALO homolog lacks an obvious glycosomal targeting motif. The recombinant T. brucei and T. cruzi enzymes accept both D-arabino-1,4-lactone and L-galactono-1,4-lactone as substrates (133, 147, 272). Recently, the cofactor of T. cruzi GAL has been identified as FAD (133) and not flavin mononucleotide as suggested previously (147). Despite the double-substrate specificity of the parasite ALO/GAL, a genomic analysis suggests that ascorbate is synthesized via L-galactose and L-galactono-1,4-lactone as it is the case in plants (Fig. 9) (272). In bloodstream T. brucei, deletion of both alleles of ALO results in cells that, after a transient growth defect, show wild-type in vitro proliferation and can produce a lethal in vivo mice infection. Depletion of ascorbate from the medium causes a transient growth defect in both mutant and wild-type T. brucei. Taken together, the data suggest that ALO is not essential for African trypanosomes but that the parasites can take up ascorbate from its environment (272). This seems not to be the case for T. cruzi, which has been reported to be incapable of scavenging exogenous ascorbate (147). Both epimastigote and trypomastigote T. cruzi contain ascorbate, the concentration in the latter mammalian infective form being two- to four-fold higher than in the insect form (47). In trypanosomatids, ascorbate is most probably kept in its reduced state spontaneously by T(SH)2, (128).
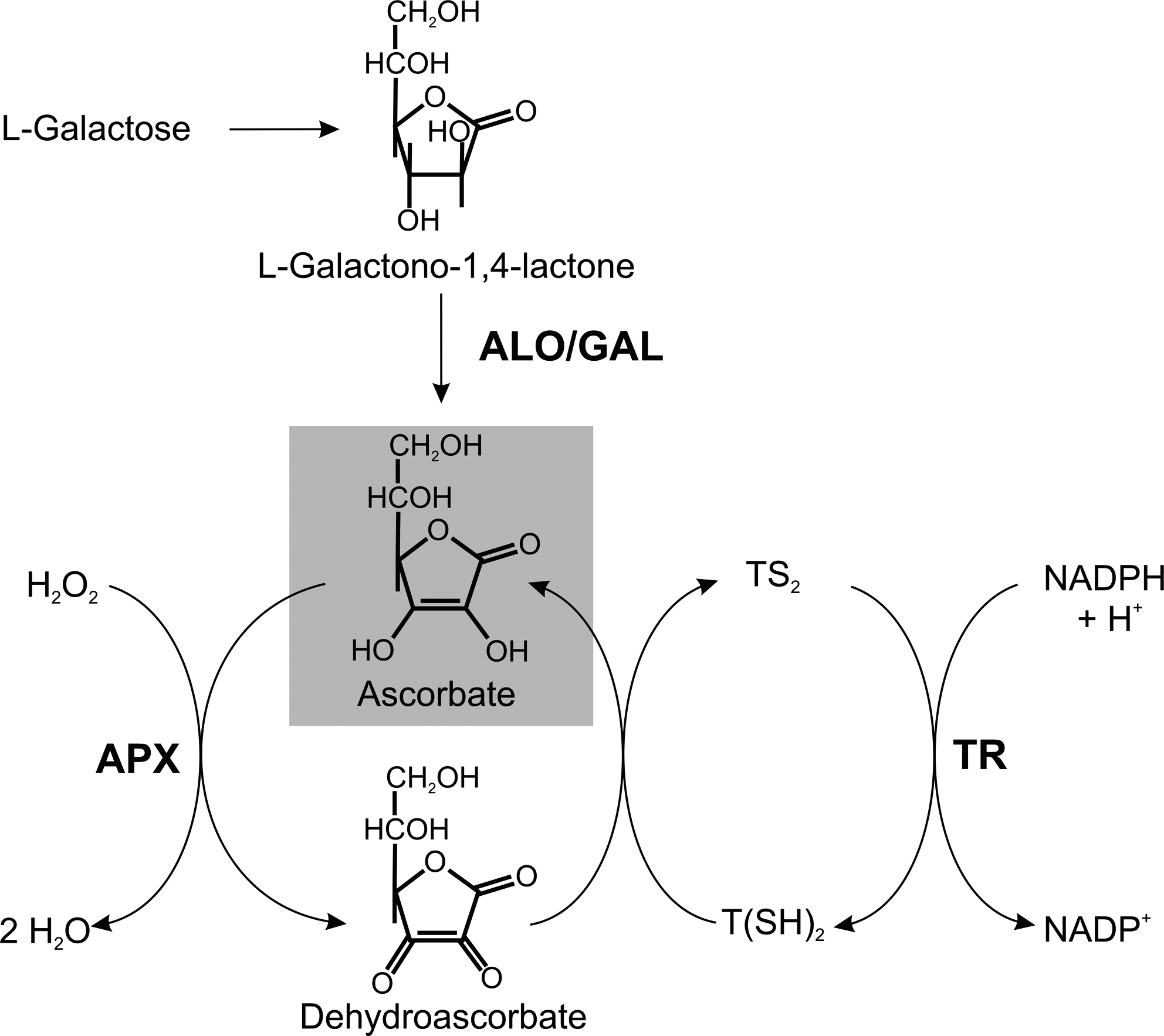
T. cruzi and L. major, but not T. brucei, possess a plant-like ascorbate-dependent heme peroxidase (APX) (1, 271). In epimastigote T. cruzi, APX colocalizes with the chaperone protein BiP, suggesting a localization in the ER (271). L. major APX has an overall sequence identity of 63% with the T. cruzi peroxidase. It has been shown to be a mitochondrial protein, localized in the intermembrane space side of the inner membrane (62). T. cruzi APX catalyzes the ascorbate-dependent reduction of hydrogen peroxide but not of cumene or t-butyl hydroperoxides (271). Both T. cruzi and Leishmania APX undergo a time-dependent inactivation by hydrogen peroxide (1, 271).
Wild-type L. major promastigotes cultured in the presence of 250 μM H2O2 express APX at levels about sixfold higher compared with unstressed cells (62). Overexpression of APX in T. cruzi or L. major results in an increased resistance toward exogenous H2O2 (63, 271). In addition, APX overexpression in L. major results in a significantly reduced intracellular H2O2 concentration and renders the cells resistant to cardiolipin oxidation induced by different mitochondrial ROS-generating drugs (62).
Recently, APX knockout mutants of L. major have been generated. The cells contain higher cellular concentrations of H2O2 and are more susceptible to exogenous H2O2 (189). Interestingly, the Leishmania knockout mutants exhibit a much higher internalization rate and improved survival in macrophages and cause a more severe mice infection compared with wild-type parasites. In contrast, APX-overexpressing parasites are practically avirulent. This may be due to the following mechanism. The virulent inoculum of Leishmania promastigotes contains a high ratio of annexin A5-binding apoptotic parasites. When these cells are depleted from the virulent population, the remaining parasites are no longer infectious (255, 262). This suggests that apoptotic promastigotes, in an altruistic way, enable the intracellular survival of the viable parasites. Indeed, cultures of APX-deficient L. major promastigotes contain a higher ratio of metacyclic and apoptotic cells compared to wild-type or APX-overexpressing cells. APX may thus play a major role in cellular differentiation of L. major and protect the parasite from apoptosis. Downregulation of APX in Leishmania seems to be crucial for disease development (189).
Summary and Outlook
Parasitic protozoa to which belong the causative agents of numerous, mainly neglected tropical diseases employ distinct small thiols as main cellular antioxidants. The redox potentials of these compounds, namely, cysteine, GSH, and trypanothione, are very similar and thus cannot account for this diversity (Table 2). In the case of Entamoeba, Trichomonas, and Giardia, life under microaerophilic conditions probably does not require a specific low-molecular-mass thiol, and Cys can act as both direct antioxidant and regulator of antioxidant enzymes. Metazoan parasites as well as Plasmodia, the causative agents of malaria, possess a GSH metabolism as it is the case in the mammalian host. In contrast, trypanosomatids developed a unique trypanothione-based system. T(SH)2 with a pK-value close to the physiological pH is a highly reactive natural dithiol that undergoes a variety of spontaneous reactions that are not fulfilled by GSH or occur at rates that are by orders of magnitude lower.
It is tempting to speculate if or which of the parasite (specific) pathways might be exploited for an antiparasitic drug development. As outlined in this review, several parasites are capable of synthesizing cysteine, but these pathways do not appear to be first choice targets. Either, as shown for Leishmania and T. cruzi, two biosynthetic pathways exist or, in the case of E. histolytica and T. vaginalis, cysteine can alternatively be taken up from the environment.
In the case of malaria, lowering the GSH levels in infected erythrocytes is supposed to mimic the protecting effect observed in natural glucose-6-phosphate dehydrogenase deficiency. In the rodent malarial parasite P. berghei, gene disruption approaches revealed that the intraerythrocytic forms require either GSH biosynthesis or GSSG reduction. This would suggest that interference with one of these pathways may not be sufficient to impair parasite proliferation. Yet, it is not clear if this is also true for human malarial parasites. The development of P. falciparum is severely affected in erythrocytes depleted of GR activity (286, 287) or GSH de novo synthesis (153). On the other hand, the fact that the activity has to be lowered by >90% renders GR a difficult target molecule (117, 286, 287). Nevertheless, a very recent study shows that GR plays a crucial role in the bioactivation of antimalarial naphthoquinones (171).
In trypanosomatids, nearly all enzymes involved in the synthesis and metabolism of T(SH)2 are essential and thus fulfill a prerequisite for a drug target molecule. With more than 60 publications, TR is so far the most intensively studied putative target protein. A drawback is that, as with GR and malaria, the activity of TR has to be lowered by >85% to impair parasite proliferation, which requires the development of highly potent inhibitors (131). Other enzymes of the trypanothione metabolism such as those involved in the synthesis of the dithiol may be even more suitable targets (178). Indeed, BSO, a specific inhibitor of GSH1, cures rodents from infection with T. brucei (12). Importantly, DFMO, a drug in clinical use for the treatment of late-stage sleeping sickness caused by T. b. gambiense is a specific inhibitor of ODC. Impairment of T(SH)2 biosynthesis is at least one crucial consequence of ODC inactivation (see Fig. 5). An attractive approach may be the development of compounds that simultaneously interfere with two steps of the pathway, for instance with TryS and TR.
A promising current drug development approach against Schistosoma and related platyhelminths focuses on TGR (137), an enzyme essential for parasite survival. Since inhibitors of the parasite TGR such as several gold complexes also interfere with human TrxR and mammalian testes express a TGR-type enzyme as well (137), it remains to be shown if parasite-specific compounds can be developed.
In conclusion, essentiality is a prerequisite but by no means sufficient for a protein to be a drug target. Other aspects such as the degree of inhibition required, the availability of a suitable in vitro test system, oral availability of the drug, and, in some cases, the ability of the compound to cross the blood brain barrier are crucial points that must be taken into account when aiming at a clinically successful drug (178, 278). Future work will show which of the parasite redox systems indeed fulfils these demands.