
Abstract
Introduction
As we have started to understand, there are a host of epigenetic influences that modulate normal infant growth and development and may play an important role in maintenance of health and origins of disease (Fig. 1). There have been some recent reviews relating epigenetics to the field child health and neonatology (36, 76). However, an approach based on risk exposure in critical periods of development will greatly add to our understanding of the role of epigenetics in infant health and disease; and this review will attempt to do that.
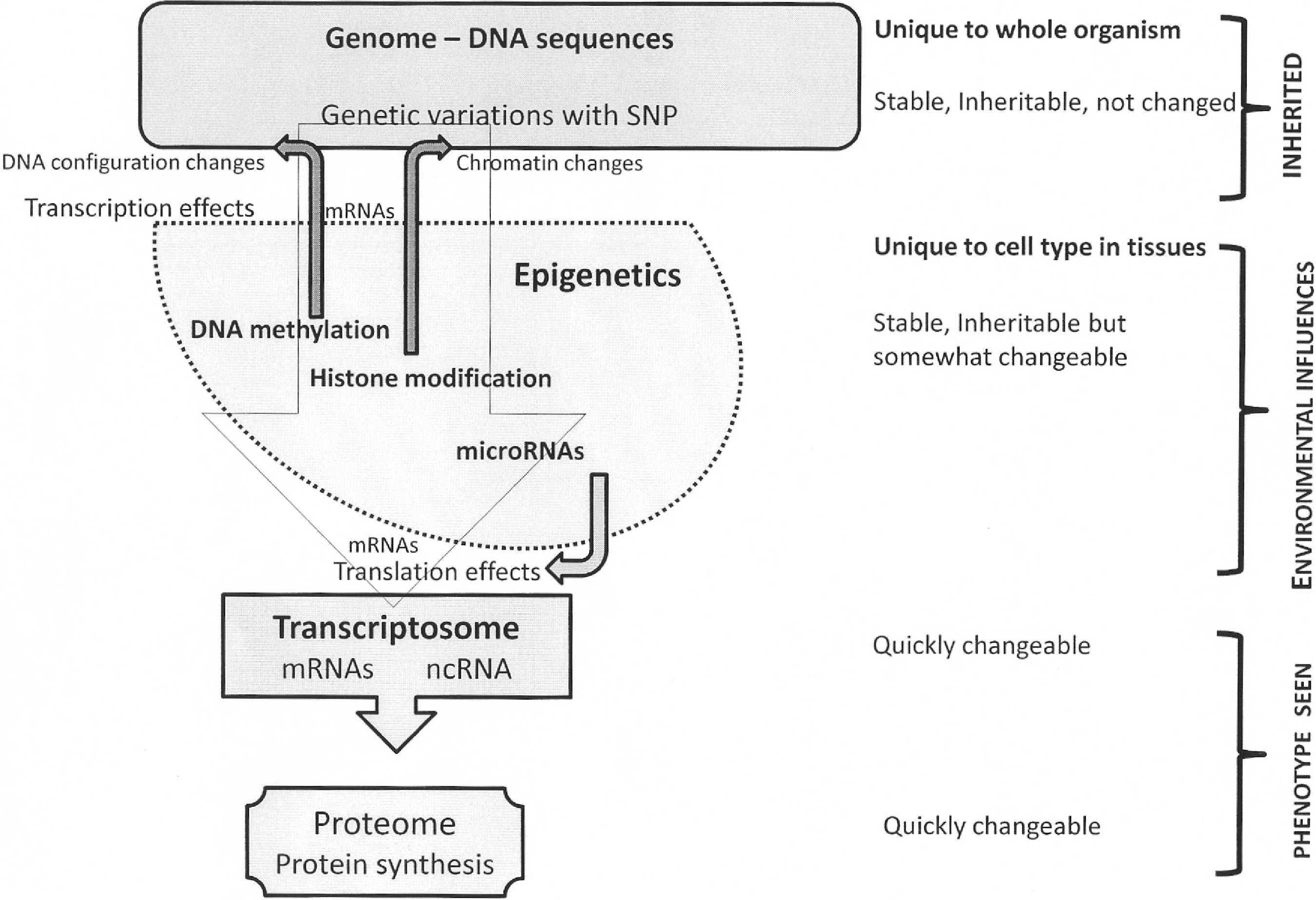
Mechanisms of Epigenetic Effects
The three fundamental mechanisms by which epigenetic changes are brought about include: 1. DNA methylation: This is the most well studied of epigenetic processes (54). In this, a methyl group is bound in the 5-carbon position of a cytosine base catalyzed by a family of DNA methyltransferases (DNMT). The cytosine most susceptible to methylation is within the dinucleotide CpG (Cytosine–phosphate–Guanine) sequence of the DNA. About 7% of the cytosine bases in the mammalian genome are known to be methylated. Gene promoter regions are usually rich in CpG sequences, also known as CpG islands, which span the 5’ end of the regulatory region of genes. Within CpG islands, methylation usually silences gene expression, and conversely demethylation activates gene expression, working as an ‘on-off switch’. Methylation patterns on CpG islands are conserved during mitosis from cell to cell (54). 2. Histone modifications: The genetic material of DNA is wrapped around a set of eight histone proteins. Based on the conformational changes to the histones brought about by presence or absence of other moieties, changes in chromatin condensation occur, leading to alteration in binding ability of promoters to gene transcription sites. Therefore epigenetic modifications of histone tails by lysine acetylation, arginine or lysine methylation, serine phosphorylation, or ubiquination can bring about changes in gene expression (46). 3. MicroRNA influences: MicroRNA (miRNA) are short 22-nucleotide, noncoding, single-stranded RNAs that are able to regulate gene translation by binding to sequence-specific base pairs in the 3′ untranslated regions of the target mRNA message. Their binding usually leads to inactivation of the mRNA. DNA methylation can influence miRNA levels, and conversely miRNA may target translation of enzymes involved in histone modification and DNA methylation (29, 33).
Epigenetic modifications may occur as isolated events or more commonly they may act in concert with genetic factors. For instance, DNA methylation of specific regions can lead to recruitment of histone deacetylases and subsequent gene silencing (48, 79). It is likely that one mechanism of epigenetic modification has influence on another, and there is probably also a contribution by purely genetic traits of an individual. The role of genetic factors on epigenetic modifications is illustrated by the observation that hypermethylation of gene promoters is seen in sputum cells of smokers and this methylation, in turn, is directly influenced by the number of single nucleotide polymorphisms that occur in genes involved in that individual's DNA repair mechanisms (57).
The timing of epigenetic modifications within an animals' lifespan is also of great significance. In pregnant adult mice exposed to Bisphenol-A, an estrogenic environmental endocrine disruptor from plastics, there were no significant changes seen in DNA methylation of the Hoxa10 gene. However, exposure in the developing fetus within these pregnant rats led to aberrant methylation in the promoter and intron of Hoxa10 gene and this altered methylation persisted after birth. These results suggest that individuals may be susceptible to epigenetic modifications only during certain vulnerable periods within a lifetime (17). It is likely that the earlier the epigenetic modification in the lifetime of an individual, the more extensive would be its effects. There are some well-defined critical periods in epigenetic programming and they occur most notably in the peri-conceptional period and in the early developing embryo (78).
Critical Periods in Epigenetic Programming
Epigenetic influences on the genetic code occur at all times during a lifespan, but there are certain critical periods of development where epigenetic marks have lasting effects not only during an individual's lifetime but in the persistence of these traits into subsequent generations (39). Some of these critical periods are: 1. Initial demethylation: At conception and in the immediate period before implantation of the embryo. 2. De novo methylation: Post-implantation development of the somatic, trophoblastic, and germ cells lines. 3. Sex-specific methylation changes: Generation of primordial germ cells and ‘imprinting’ of genes—first in utero and later during puberty and gametogenesis. 4. X chromosome changes: Two phases of X chromosome inactivation in females.
Epigenetic regulatory mechanisms are key processes during pre-implantation development of the embryo (22). Soon after fertilization the zygote undergoes rapid demethylation of the male genome (69, 83). The female genome too is demethylated passively during the mitotic divisions that follow (58). Global methylation then starts, and a number of CpGs are methylated prior to the blastocyst stage of embryogenesis (88). Demethylation at this stage affects almost all genes except the ‘imprinted genes' (which are epigenetically modified at a different critical period of gametogenesis). The important distinction in genes that are ‘imprinted’ is that the methylation patterns are established in the gametes before fertilization and these areas are excluded from the gene-wide demethylation that occurs after conception (15). DNA methylation is a complex process and in mammals a key set of enzymes, the DNA methyltransferases (DNMT), are involved. De novo DNA methylation is catalyzed by DNMT-3a and -3b enzymes, and DNMT-1 is responsible for maintenance of established CpG dinucleotide methylation patterns during mitosis. During this initial critical phase in the early embryo, besides DNA methylation, histone modifying proteins also play an important role. Their role has been well studied in mechanisms that help in changes from pluripotency to the differentiated cell state. In early embryogenesis, a set of histone modifying proteins termed PcG (Polycomb group complex) proteins play an important role in silencing cell determination genes such as Pax, Hox, and Dlx and keep the cell mass in the pluripotent state (4, 106).
The next critical phase is methylation of the genome following the initial wave of demethylation. The environmental milieu and availability of one-carbon moieties are especially critical in this phase in establishing a normal methylation pattern. During this phase, the pattern of somatic, trophblastic, and primordial germ cells methylation is different and occur at different rates (78) (Fig. 2).
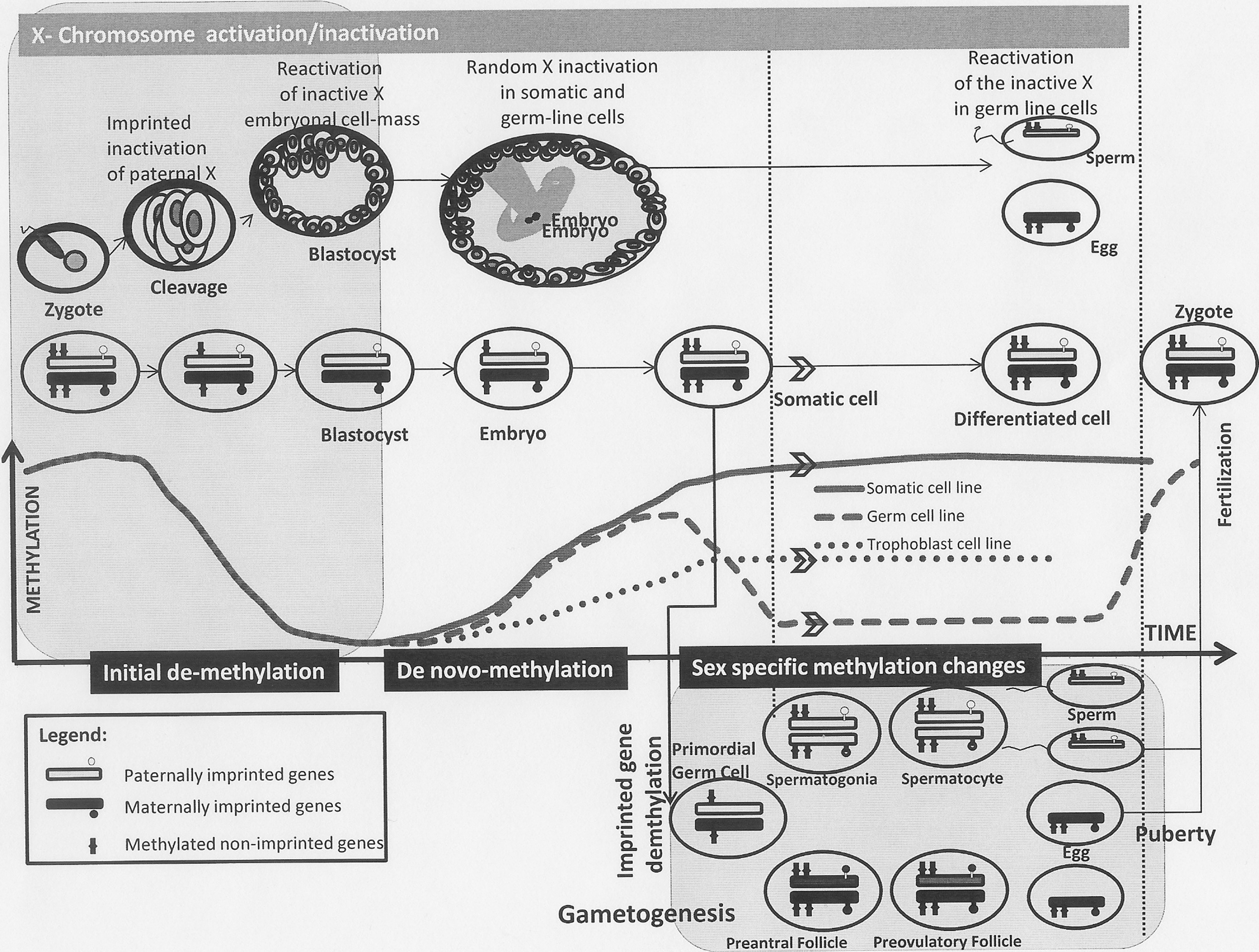
A series of DNA methylation is seen within the germ cell line and occurs during two critical stages of life. The first part of germ cell epigenetic modification occurs during primordial germ cell development within the fetal gonadal ridges where demethylation of all single copy genes and some repetitive elements occurs (56). A second wave of methylation in germ cells occurs after birth, starting at puberty, and is important in gametogenesis. Germ cell methylation is most significant for the ‘imprinted genes' whose methylation patterns are established during this process. The imprinted genes in turn have a major role to play in the epigenetic changes related to DNA demethylation and methylation that occur during the normal course embryogenesis (114). Germ cell epigenetic modifications are especially important in transgenerational changes that are seen as a result of maternal–fetal environmental or nutritional disruption.
Another important epigenetic modification that occurs during early embryogenesis is the X chromosome inactivation in females that was first described by Lyons (64). This process recently has been found to be due to epigenetic inactivation of the extra X chromosome by interplay of DNA methylation and histone modification (64, 118). The process of X-chromosome inactivation is complex. Paternally acquired X chromosome inactivation in females is initiated after fertilization, during the cleavage-stage embryos. In cells that form trophoblasts, the inactivation persists. But within the embryonic cell mass, both X chromosomes become active for a period of time corresponding to embryonic day 3.5 (E 3.5) and E 5.5 in the mouse (118). Subsequently, there is a random inactivation of either the paternally or maternally derived X chromosome starting from E 5.5 (in mouse) and is maintained through the adult somatic and germ cell lines. The next phase of reactivation of the X chromosome occurs in the developing germ line so that both X chromosomes are again active during oogenesis. A similar process probably happens in humans.
Early Life Influences on Epigenetic Changes in Humans
Given that the ‘critical periods' of epigenetic modifications are mostly in early intrauterine life of humans, we will now detail the effect of various influences on this process and its potential impact for future health and disease.
Preconception environment
Pre-conceptional maternal health, 1-carbon metabolism status, especially folate levels, and maternal diet appear to have important influences, as have been shown by animal experiments (21, 47, 122).There has also been some epidemiological evidence in humans (43, 67, 116, 119). A number of reviews on this subject have been published, and the importance of nutrition, especially protein availability, calorie intake, and the availability of one-carbon moieties have been well described (12, 21, 35, 62–63, 96, 122). The specific effects from folate supplementation in diet will be discussed later in this review.
Assisted reproductive technology
Embryos from in vitro conception are associated with lower mean methylation at CpG sites in placenta and higher mean methylation at CpG sites in cord blood. There are also variations in gene-specific DNA methylation between the in vivo and the in vitro conception groups (51). There is epidemiological evidence that conditions related to improper imprinting, such as Beckwith-Wiedemann syndrome, are more common in babies born after in vitro fertilization. In this condition, DNA methylation of the IGF2 gene promoter within the chromosome 11p15 region is thought to be responsible (25). Other specific genes such as PEG10 and L3MBTL are upregulated and PHLDA2 downregulated in ART-conceived offspring but the significance of these findings are not yet known (30). There is also evidence that cloned animals subjected to embryonic cell replication in vitro develop different disease patterns than genetically identical donors (51).
Folate and 1-carbon availability related effects
Nutrients that contribute to methyl-group one-carbon metabolism, such as folate, can influence epigenetic effects during critical periods of development. These have been well illustrated using the agouti mouse model and the modification of the coat colors by various nutritional restrictions and supplements (21, 122). In human studies, folate antagonists use during pregnancy was associated with increased risk of cleft lip and palate (CLP) and increased neural tube defects(NTD) (42). Dietary supplementation with folic acid in the general population has decreased the occurrence of both CLP and NTD (119). Other similar substances that affect one-carbon metabolism, especially pyridoxal phosphate, choline and symmetric di-methyl-arginine, have also been associated with differing frequency of CLP (97). Methylation patterns in IGF2 promoter P2 of maternal blood were associated with vitamin B12 levels but not folate levels during pregnancy (5). The probable relationship of this finding to fetal growth in utero is intriguing. Lower maternal folate status in early pregnancy may impair fetal brain development and thus increase likelihood of hyperactivity and peer problems in children (96).
Conversely, modification of one-carbon group availability at these critical periods of development may also be implicated in disease causation. In utero supplementation with methyl donors appears to enhance allergic airway disease in mice and this effect is transgenerational, implicating epigenetic changes (43). There have been some recent reports implicating folate fortification of diet and increased allergic human disease (116). There is no clear evidence of these effects and the causal relationships of these phenomenon are still being debated (67).
Environmental agents
A number of environmental agents and stressors are known to cause epigenetic changes (13, 108). Some of environmental agents that have been shown to cause epigenetic modifications include endocrine disruptor chemicals (EDC), diethylstilbestrol (DES), tobacco smoke, and alcohol. Other agents that humans are commonly exposed to such as marijuana, opiates, and serotonin uptake inhibitors are being studied for their ability to cause epigenetic modifications (31, 81).
There are many examples of exposure to endocrine-disrupting chemicals and epigenetic programming during in-utero development that may result in later adult-onset disease (121). Methoxychlor and vinclozoline are well-studied EDC chemicals found in the environment. Methoxychlor causes DNA hypermethylation in the ovary associated with increase DNA methyltransferase activity, specifically DNMT3B (121). Another EDC, vinclozoline, has been shown to epigenetically alter the sperm epigenome for up to the third generation after in utero exposure (37, 98). Many human pregnancies have vinclozoline exposure and its relation to fetal endocrine disruption and development of genital tract defects such as hypospadias has recently been demonstrated (109).
Diethylstilbestrol (DES) was one of the first therapeutic agents that were shown to cause transgenerational problems in mothers and their offspring. The transgenerational effects of DES in causing breast and genital tumors in women is an example of epigenetic changes transmitted through germ line alterations of the female fetus in utero. Specific link to an epigenetic marker has been shown in animal models of DES exposure. Enhancer of Zeste Homolog 2 (EZH2) is a histone methyltransferase that has been linked to breast cancer risk and epigenetic regulation of tumorigenesis. Mice exposed to DES in utero showed a > 2-fold increase in EZH2 expression and protein levels in adult mammary tissue compared with controls (26). Similarly, mice exposed to DES in utero showed increased mammary histone H3 trimethylation. Developmental programming of EZH2 may be the mechanism by which in utero exposure to endocrine disruptors such as DES leads to epigenetic regulation of the mammary gland (26).
Maternal smoking is an important risk factor for poor fetal growth and poor lung function, and these effects may be epigenetically regulated. Maternal tobacco smoking and prenatal smoke exposure of the fetus affecting global DNA methylation can be studied by evaluating some surrogate markers such as interspersed DNA repetitive elements. The most commonly studied of these repetitive elements are long interspersed nucleotide element (LINE1) and a short interspersed nucleotide element termed AluYb8. Maternal tobacco smoke exposure has shown demonstrable differences in global DNA methylation, especially within AluYb8 but not LINE1 (16). There are also demonstrable effects of tobacco exposure on gene-specific DNA methylation, the significance of which is under study (16). Global DNA methylation rates vary in women based on many factors, including maternal smoking during pregnancy, later age at menarche, and later age at first birth among multiethnic women in New York (105). Maternal smoking also affects insulin-like growth factor 2 (IGF2), which is an important regulator of fetal growth. Alteration in methylation patterns in IGF2 promoter P2 were associated with exposure to passive smoking during pregnancy (5).
Maternal alcohol consumption is known to have serious effects on the fetus encompassed within the diagnosis of fetal alcohol spectrum disorders. One mechanism for this may be related to the effect that alcohol has on DNA methylation pathways. Alcohol has been shown to reduce the expression of the DNMT3b mRNA associated with an increase (10%) in genomic DNA methylation in human subjects (14). There are also increased levels of homocysteine in patients with alcoholism and this 1-carbon metabolite may have important epigenetic influences in utero (11). Recent studies in ethanol-treated rats has demonstrated that CREB binding protein (CBP) which is widely expressed in granule and Purkinje neurons of the developing cerebellar cortex, have reduced levels in pups born to alcohol exposed pregnancies (38). CBP is a histone acetyltransferase, and a component of the epigenetic mechanism controlling neuronal gene expression. Consequently, acetylation of both histone H3 and H4 was reduced in the cerebellum of ethanol-treated rats (38). This may help explain the motor coordination deficits that characterize fetal alcohol spectrum disorders.
Maternal nutrition during pregnancy
Maternal nutritional deprivation, as best illustrated by pregnancies during natural famines, has been studied extensively in the cohort from Amsterdam that survived the ‘Dutch Hunger Winter’ of WWII. Within this cohort, there have been demonstrable changes in glucose metabolism and risk for adult onset disease such as hypertension, coronary heart disease, and stroke (41). Some of these effects have been explained by changes in methylation of the IGF2 gene and its persistence into adulthood (41). More specific epigenetic changes related to intake of other nutrients are being extensively studied in animal models (62, 63).
Stress and steroids
Prenatal environment may program our stress response throughout life (68) (Fig. 3). The effect can be due to influences acting not only on the embryo and the fetus in the womb but also in the nurturing that is done in the newborn period and early infancy (99, 114).
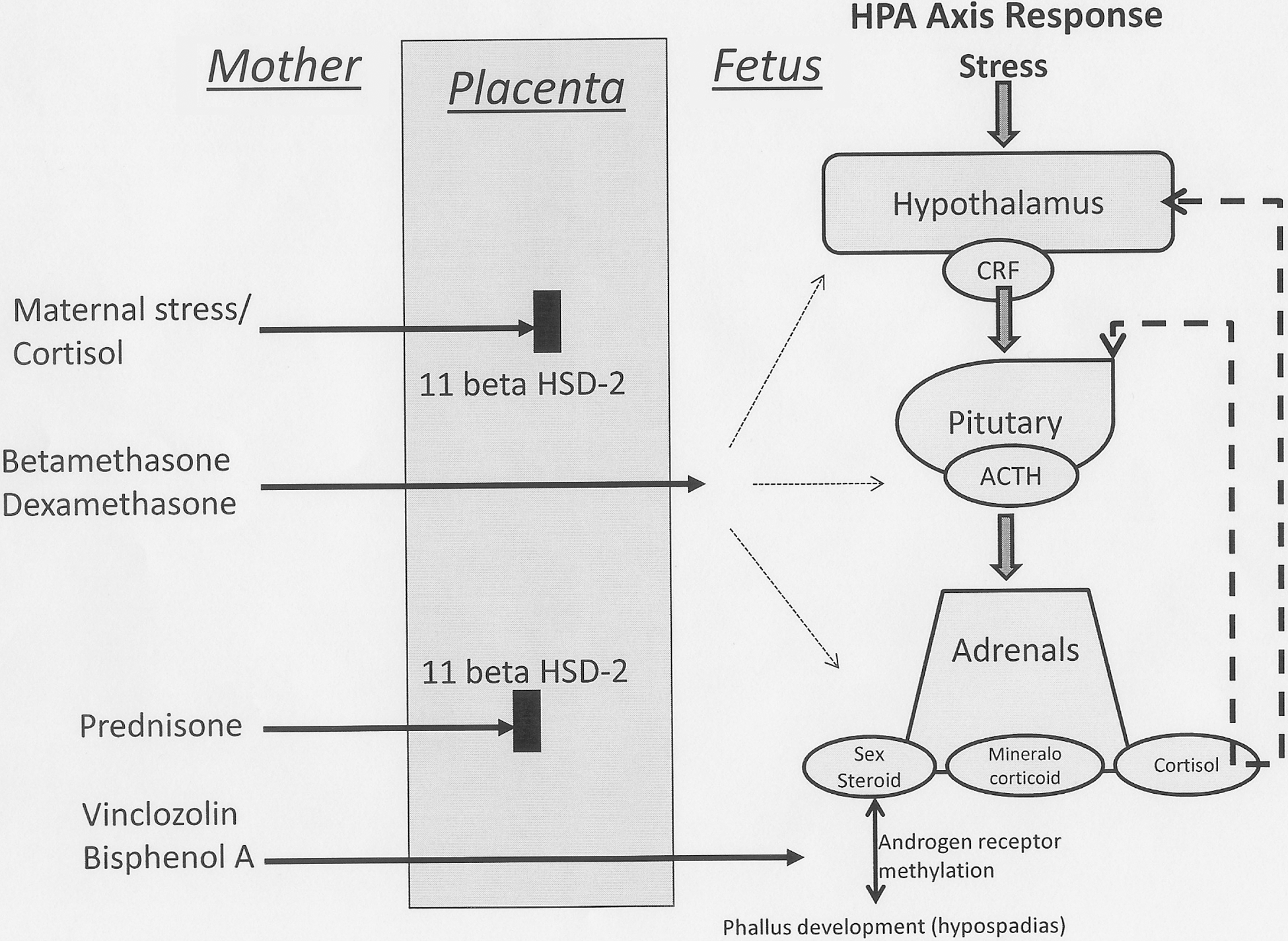
The effects of maternal stress on the fetus has been correlated with alterations in its hypothalamic–pituitary–adrenal axis (HPA axis) (86). Studies of maternal stress and its effects on fetal nervous system development and function show that some of these effects in offspring persist into later childhood and adulthood (93, 94). The mechanism for this persistent effect is not clearly elucidated but epigenetic modifications may play an important role. There is evidence to suggest that selective methylation/demethylation of specific cytosine residues occurs in the glucocorticoid receptor I7 promoter in response to stressful stimuli (112). An epigenetic modification of the transcription nerve growth factor inducible protein-A gene is thought to be responsible for this effect (71, 113).
Stress after birth in the newborn or in early infancy is associated with increased arginine-vasopressin (AVP) gene expression. In animal studies, this has been found to occur due to hypomethylation of the region where methyl CpG-binding protein-2 binding occurs (MeCP2); and MeCP2 is known to silence gene expression (77). Therefore a sustained increase in AVP gene expression in the hypothalamic paraventricular nucleus (PVN) results in persistently high activity of the HPA axis (77). Interestingly, MECP2 gene mutations that lead to altered methylation and changes in chromatin structure in humans cause Rett syndrome (developmental delay/autism) (19).
Another piece of evidence for postnatal epigenetic modifications comes from studies of the brains of childhood abuse victims who committed suicide. These brains showed increased methylation (silencing) of the NR3C1 exon 1F which is known to regulate glucocorticoid receptor (GR) expression in the hippocampus (70). There was also a corresponding decrease in GR mRNA levels in these brains (70).
Intrauterine growth restriction
The role of intrauterine growth in the development of adult disease was initially enunciated in the ‘Barker's hypothesis' (6). Subsequently, there has been a body of work that has emerged to support this hypothesis, and epigenetic modifications have been shown to be responsible for fetal programming in intrauterine growth restriction (IUGR). A host of adult-onset diseases have been linked with IUGR on the basis of epigenetic modification and some specific conditions are briefly mentioned below.
Adult-onset noncommunicable diseases such as cardiovascular disease, obesity, and metabolic syndrome have been associated with changes that occurred during early fetal development. The population in Amsterdam during WWII that was exposed to the ‘Dutch Hunger Winter’ has been a source of many studies looking at the persistence of epigenetic changes relating to human exposure to famine conditions. In those individuals exposed to famine conditions in utero, decreased methylation of the maternally imprinted IGF2 gene has been shown to persist for many decades (41). The close association of higher incidence of schizophrenia and coronary artery disease in famine survivors awaits further study (84, 100). Also of interest is the observation that endocrine or nutritional interventions during early postnatal life may be able to reverse some epigenetic and phenotypic changes (39).
Obesity in later childhood or adulthood is more common in growth-restricted infants. Changes in global DNA methylation of repetitive elements such as LINE1 are nonspecific epigenetic markers, and recently it has been found that significant differences in cord blood LINE-1 methylation occurs among newborns with low and high birth weight, as well as among prematurely born infants (72). In another study, the methylation status of 68 CpGs in the 5' region from five candidate genes in umbilical cord tissue DNA were studied and correlated with childhood adiposity at 9 years of age. Methylation varied greatly at particular CpGs but the epigenetic state of a single CpG site in the RXRA transcription factor promoter region was strongly associated with later childhood obesity in both sexes (35).
Hypertension has been correlated with low birth weight and intrauterine growth restriction, suggesting fetal programming. In rats with adult onset hypertension, modification of the renin-angiotensin system by changes in methylation of the AT1b promoter has been demonstrated to be induced by changes in diet during pregnancy (12). Epigenetic modifications affecting somatic angiotensin converting enzyme function has been demonstrated in humans (89).
Mental and motor function modifications have been correlated with epigenetic changes in growth-restricted babies. Autism spectrum disorder (ASD) has been associated with IUGR, especially in premature infants (44). Recent evidence suggests a role for changes in DNA methylation and genetic imprinting in ASD (52). Fetal body size and symmetry have been associated with differences in infant motor development at 9–15-week age in a population cohort from Rotterdam (107). Epigenetic changes have also been shown to affect motor functioning in animal models (66). Epidemiological evidence is accumulating in supporting a role for epigenetic influences in the causation of major psychoses (92). Schizophrenia has been correlated with epigenetic changes caused by the ‘Dutch Hunger Winter’ (100). In studies with monozygotic twins that share a similar genotype but express different mental health phenotypes, the role of epigenetics is well illustrated (32). Genes involved in schizophrenia such as dopamine D2 receptor and the catechol-O-methyltransferase gene have shown differences in methylation in monozygotic twins with and without the diagnosis (73, 85).
Reproductive function is altered in IUGR infants as manifested by increased incidence of hypospadias in boys and ovarian dysfunction in girls. The incidence and severity of hypospadias is directly related to the timing and degree of severity of IUGR (45). This may be related to altered steroid metabolism in utero in IUGR infants. In human studies, the steroid androgen receptor (AR) gene in target tissues from patients with hypospadias was more methylated (silenced) than in control children, resulting in a decreased expression of the AR. The mechanism underlying the modulation of the AR gene expression seems to be mediated by DNMT-3A. This epigenetic alteration of the AR gene might be involved in the pathogenesis of hypospadias (Fig. 3) (110). In females, there may be a similar correlation of IUGR with epigenetic changes affecting sex steroid action and development of polycystic ovary syndrome in later life (60).
Placenta and Epigenetic Modifications
One of the mechanisms underlying fetal origins of adult disease may be an alteration of the epigenome in the placenta as well as the fetus (7). Maternal-placental and baby phenotypes have been correlated with the risk of future lung cancer from a cohort born in Helsinki between 1924 and 1944. In this study it was suggested that a complex mechanism involving low amino acid–high glucose delivery to the fetus in early gestation, impairment of placental antioxidant system, and subsequent vulnerability to agents such as tobacco smoke may have been responsible (7). Epigenetic regulation of the placenta may also have played a part. Some of the placental elements that are known to have epigenetic modifications are the human growth hormone gene cluster (hGH) and the 11 beta hydroxyl-steroid dehydrogenase (HSD) enzymes. The hGH gene cluster contains a single human pituitary hormone gene (hGH-N) and four placental specific paralogs. Activation of the cluster depends on 5’ remote regulatory elements that are known to be regulated by histone methylation and acetylation (53). Placental 11 beta hydroxyl-methyltransferase (HSD-1 and HSD-2) are important in the regulation of maternal steroid cross-over from the mother to the fetus in utero. Specifically, 11 beta HSD-2 deactivates maternal stress steroids and other medications such as prednisolone but has no action against betamethasone or other environmental endocrinal agents such as vinclozoline and bisphenol-A (Fig. 3). HSD-2 enzyme levels in the placenta that are responsible for preventing unregulated stress steroids crossing over from mother to fetus, have also been shown to be subject to epigenetic modifications. Differential methylation has been shown of the CpG islands influencing the promoter and exon 1 regions of the HSD11B2 gene, both in human cells in culture and in vivo in rats (1). Therefore the role of the placenta in epigenetic modifications of the fetus could be direct by changes in its own activity as in hGH, or indirect by its role in allowing transfer of bioactive agents.
Birth, Rearing, and Early Life Effects on Epigenetic Modifications
Not only the fetal period of life but conditions at birth and in early infancy may be important in modulating behavior and function through epigenetic modifications. Even the mode of delivery may have an effect. Global methylation levels in fully differentiated WBC from cord blood are higher in those born by C-section versus normal vaginal delivery. But the methylation status reverts to no difference within 3–5 days (95, 102). This suggests a dynamic epigenome that is modifiable during life. Animal experiments show that some of these effects are reprogrammable with use of methyl group modifiers such as trichostatin-A and L-methionine (111). Early life social adversity leaves its impression on the epigenome of an individual and may influence the person's social and biological adaptations for a lifetime (101). Early grooming and nurturing behavior may have lifelong consequences because of epigenetic modifications (111, 112). For example, variations in maternal grooming influences HPA axis stress reactivity via long-term changes in expression of glucocorticoid receptor expression. In animal studies, this is related to increased hippocampal seretonergic tone along with increase histone acetylase activity, histone acetylation, and DNA methylation mediated by the transcription nerve growth factor-inducible protein-A (111).
Epigenome in Genetically Identical Twins
Genomic pattern is uniform among different cells of a complex organism, but the epigenome varies from tissue to tissue. This difference gets larger with increased differentiation of the tissue. Similarly, monozygotic twins start with similar epigenomes but accumulate epigenetic differences over their lifetime (32). As mentioned earlier, genes involved in schizophrenia have differences in methylation patterns in monozygotic twins with and without the diagnosis (73, 85). Twin studies have also shed light on the epigenetic influences in the disease phenotype of primary biliary cirrhosis (PBC) (75). Primary biliary cirrhosis is an autoimmune chronic cholestatic liver disease with a strong genetic susceptibility due to the high concordance in monozygotic (MZ) twins and a striking female predominance. It has been shown that in this condition epigenetic factors influencing PBC onset are complex, and not only methylation differences at X-linked promoters but variably three inactivated X-linked genes may be influenced by partial promoter methylation and biallelic transcription (75).
Genetic Imprinting and Epigenetics
There is an increasing list of diseases related to alterations in genetically imprinted genes (Table 1). Genetic imprinting is a form of epigenetic modulation but is different in that the methylation changes in DNA occur during gametogenesis and thus the factors that influence these effects have a different critical window.
X-Chromosome and Epigenetics
The presence of two X chromosomes in females results in the random inactivation or silencing of one of them in somatic cells as per Lyon's hypothesis (64). The silencing of this entire chromosome involves a complex set of epigenetic modifications involving both DNA methylation and histone modification changes (118) (Fig. 2). However, there may be important effects of the pattern of X-chromosome inactivation. As mentioned above, some disease such as primary biliary cirrhosis illustrate this effect well (75).
Transgenerational Effects
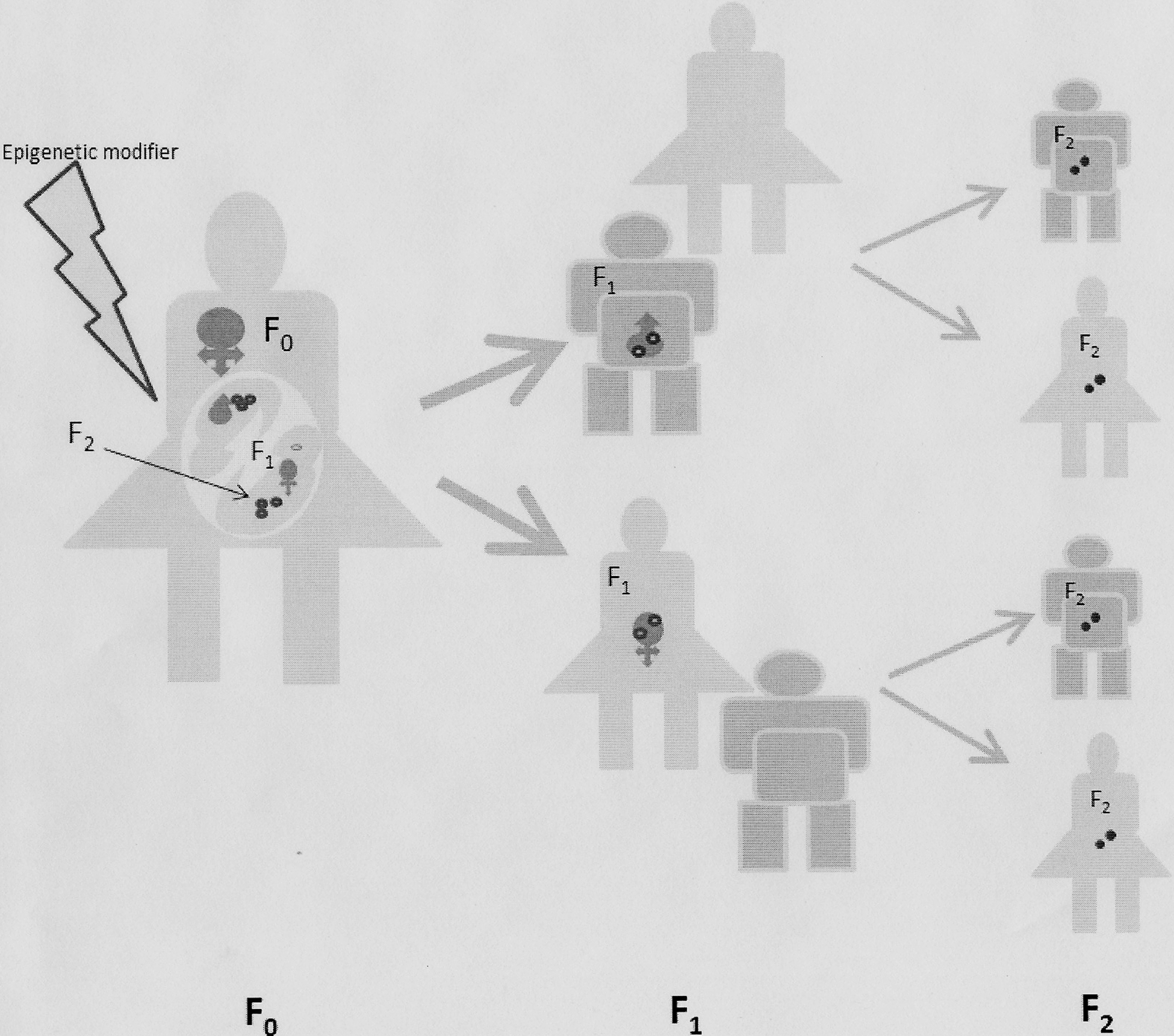
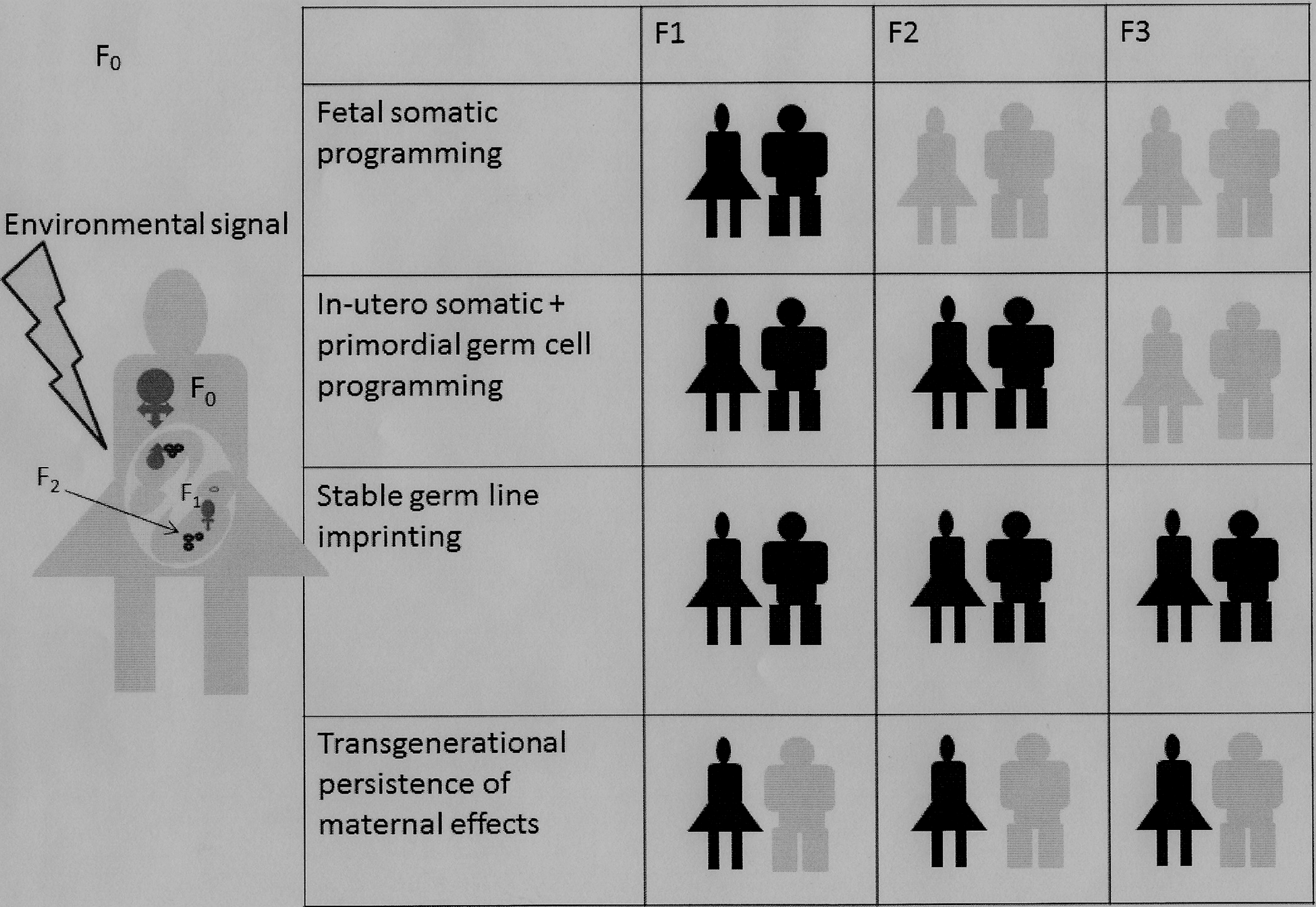
There are a number of conditions in which the epigenetic influences have been shown to persist into second and third generations after the initial modifications.(Fig. 4). Animal experiments with vinclozolin and persistence of changes in the third generation epigenome of the sperm is one such example (37, 98). In humans, maternal smoking and asthma have been associated in not only offspring but in maternal grandchildren (59, 87). The ‘uterine effect’ transgenerational programming usually stops at the third generation, as has been demonstrated in animals exposed to dexamethasone and others subjected to maternal undernutrition (10, 40). Human grandparent effects have also been shown on mortality related to cardiovascular effects and diabetes (49). There are important epigenetically influenced sex differences in the transmission of characteristics in offspring when maternal stress and or diet is modified (27). This is because of germ-line inheritance that is seen in both sexes versus transgenerational perpetuation of maternal effect that is only seen in female and not in male offspring (27). However, it is important to remember that not all transgenerational epigenetic effects are a result of epigenetic inheritance (120) (Fig. 5).
Role of Epigenetics in Development Beyond Infancy
In the continuum of growth and development from infancy to adulthood, there appear to be ongoing epigenetic changes, albeit not as profound as those during the ‘critical periods' that may be responsible for the phenotypic effects of aging. Epigenetic drift may be involved in the aging processes (9, 20). Moreover, contrary to widely held belief, there appears to be ongoing neuronal production in the adult hippocampus. There are suggestions that there is dynamic and reversible DNA methylation in hippocampus in adults that may be important in memory and neurogenesis (23). The role of epigenetics dysregulation in oncogenesis is beyond the scope of this review.
Focus for Future Innovations
An important consequence of our understanding of the epigenetic process is the discovery of agents that may be used in its modification for the promotion of health and treatment of disease. These may range from simple dietary modifications focused on the ‘critical periods' to the use of agents that actively alter the epigenetic state. For some noncommunicable disease it has been found that endocrine or nutritional interventions during early postnatal life may be able to reverse the epigenetic changes that were programmed in utero (39). Some animal studies have demonstrated efficacy of treatments in altering epigenetic modifications. Dietary modifications in rats fed protein-rich diets during pregnancy have shown alteration in CpG sites in the promoter regions of the PPARα gene that has effects on development of obesity and metabolic syndrome. However, this effect was only seen in the liver but not in skeletal muscle or adipose tissue (62). There have also been demonstrable changes in the expression of DNA methyltransferase in these animals (63). One-carbon metabolites such as trichostatin-A and L-methionine have been used in laboratory animals to modify certain epigenetic changes that occur related to the birth process (111). There are many agents under study that have the ability to alter methylation patterns on DNA or can modify histones (28). Some agents undergoing Phase I trials that may effect this by DNA methylation include: 5-azacytidine, 5-aza-2’-deoxycytidine, epifallocatechin-3-gallate (EGCG), and specific antisense oligomers for specific DNA regions (28). There are also some histone deacetylases undergoing Phase I and II trials, including phenylbutyric acid, suberoylanilide hydroxamic acid (SAHA), depsipeptide. and valproic acid (28). Some of these agents are known to have developmental effects on the fetus and the key would be to use them to target specific critical periods of methylation/demethylation during development. Another strategy may be to use a sequential combination to silence or unsilence a gene with one agent and once a gene is in an altered state to make another alteration with another agent to prevent it from reverting back to its original state (28). Obviously, more study in this field needs to be done.
Conclusion
Epigenetic influences modulate the genetic code of an organism in response to its environment and are important biological mechanisms that help in individual adaptation. However, adverse environmental signals are shown to cause epigenetic changes that may persist through many generations. An important aspect of epigenetic modification is its potential for reversal. A full understanding of the risk factors and the critical periods in which they act will inform us of potential therapeutic approaches to reverse the undesirable changes. The potential for therapeutic modification of epigenetic signals may be the next frontier for drug discovery and development.
Footnotes
Acknowledgment
I would like to thank Tasneem Hussain, MOTR/L for her support in preparing this review.
Author Disclosure Statement
The author has no commercial associations that might create conflict of interest in connection with the submitted manuscript file. No competing financial interests exist.