
Abstract
Introduction
Innovation
Carbamylation by cyanate is known to occur non-enzymatically when urea is dissociated, and also by myeloperoxidase catalyzed oxidation of thiocyanate. Carbamylation of low-density lipoprotein (LDL) is suggested to increase atherosclerosis. In this study, we showed that immunization with carbamyl-mouse-LDL without adjuvant resulted in specific immunoglobulin G (IgG) immune response in mice. IgG antibodies to carbamylated proteins were also found in humans, and they were associated with conditions with enhanced carbamylation, uremia, and smoking. IgG antibodies to carbamyl-epitope might serve as a novel indicator of carbamylation in vivo in uremic patients or smokers, and need to be further investigated.
Although carbamylated proteins have been demonstrated to exist in vivo in humans and to predict the future risk for cardiovascular events (46), the presence and role of this modification as an antigenic trigger for the human humoral immune system is largely undisclosed. Recently, post-translational modification by carbamylation was demonstrated to result in CD4+ T cell activation, proliferation, and chemotaxis, subsequently influencing experimental erosive arthritis in mice (29), which suggests involvement of the immune system in recognition of carbamylated epitopes. Indeed, serum immunoglobulin G (IgG) antibodies recognizing homocitrullinated (carbamylated ɛ-amino groups of lysines) antigens have been detected in sera of patients with rheumatoid arthritis and with no cross-reactivity to citrullinated proteins (38).
Immunizations with modified LDL (3, 18, 31, 35, 49) and antibodies generated against various modifications of LDL have been suggested to modulate atherogenic processes in animal models of atherosclerosis, including LDL receptor-deficient (LDLR −/−) mice (3, 10, 18). Modifications of homologous LDL, such as those generated during carbamylation, make it highly immunogenic by inducing antibody formation specific for the derivatized lysine residues (40). The aim of the present study was to investigate activation of the primary humoral immune system to recognize carbamyl-haptenated proteins, especially carbamylated LDL, and the role of this immune recognition in atherogenesis in LDLR −/− mice. In addition, the presence of autoantibodies binding to carbamylated proteins in humans under conditions of enhanced carbamylation was investigated.
Results
IgG immune response in mice immunized with carbamyl-mouse LDL
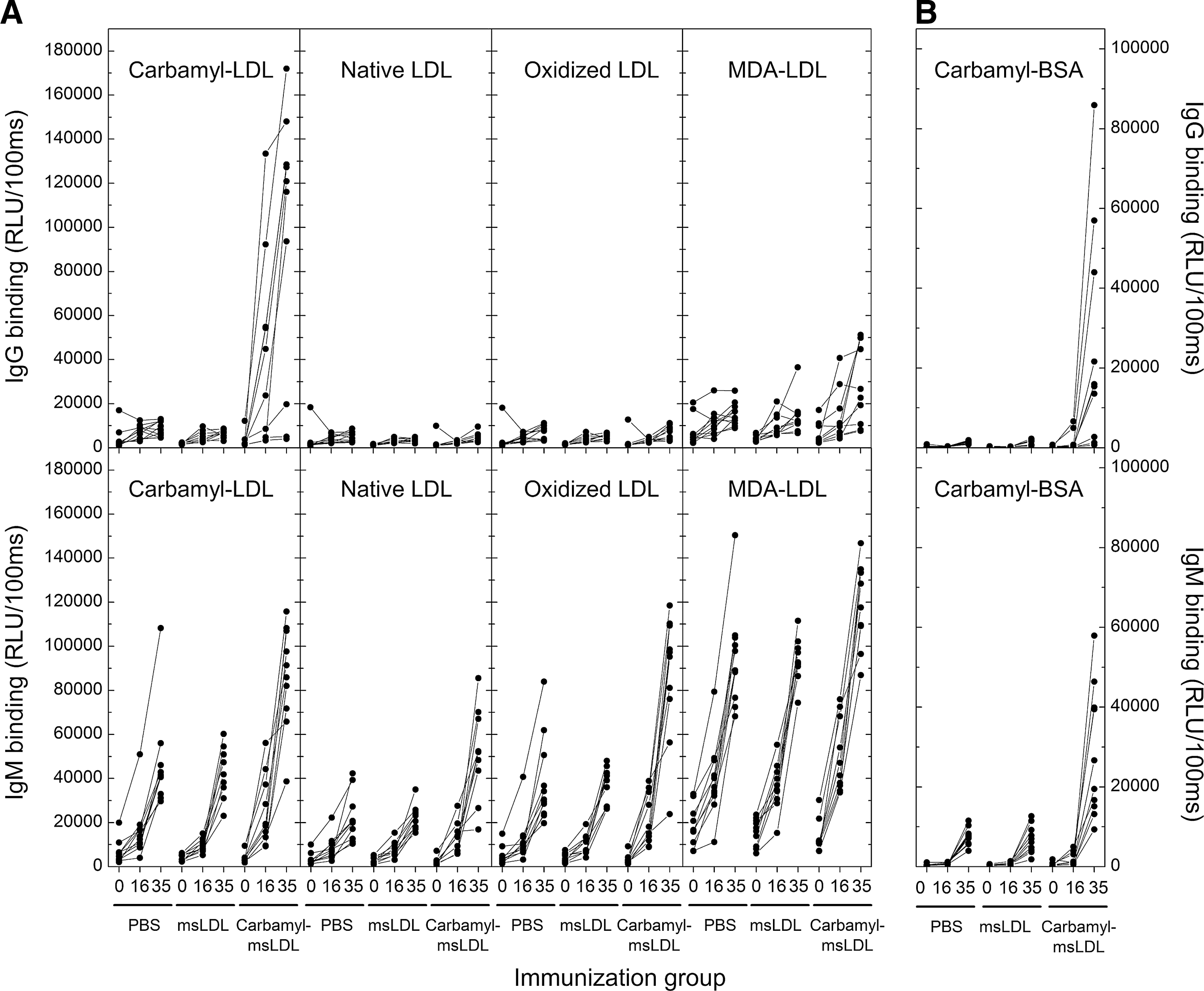
LDLR −/− mice were immunized with carbamylated mouse LDL (msLDL) (n=10), native msLDL (n=10), or phosphate-buffered saline (PBS; n=11). Plasma dilution curves for IgG binding to carbamyl-LDL for one mouse from each immunization group are shown in Figure 1A–C. The carbamyl-msLDL-immunized mouse demonstrated high plasma IgG binding to carbamyl-LDL (Fig. 1A), whereas the mouse immunized with native msLDL (Fig. 1B) or PBS (Fig. 1C) had no IgG antibodies to carbamyl-LDL. We also tested plasma IgG levels to native LDL, oxidized LDL, and malondialdehyde-modified LDL (MDA-LDL) because LDL was used as a carrier molecule in the immunizations. Carbamyl-msLDL-immunized mouse had moderate levels of plasma IgG binding to MDA-LDL but not to oxidized LDL or native LDL (Fig. 1A). Mouse immunized with native msLDL (Fig. 1B) or PBS (Fig. 1C) did not have IgG antibodies binding to native LDL, oxidized LDL, or MDA-LDL.

To study the immune response in all mice (n=31) throughout the experiment, the antibody binding was tested in samples taken at different times of the study. Mice immunized with carbamyl-msLDL had increased plasma IgG levels to carbamyl-LDL (Fig. 2A), compared to mice immunized with native msLDL or PBS in samples taken after immunizations and after high-fat diet (HFD). There were no statistically significant differences in plasma IgG levels to native LDL, oxidized LDL, or MDA-LDL between the three immunization groups (Fig. 2A). To investigate whether carbamyl-msLDL immunization resulted in autoantibody response directed to other carbamylated proteins than LDL, the IgG binding in mouse plasma samples was also tested with carbamylated bovine serum albumin (carbamyl-BSA). Mice immunized with carbamyl-msLDL had increased plasma IgG levels also to carbamyl-BSA at the end of the study (Fig. 2B). There were no IgG antibodies binding to carbamyl-BSA in mice immunized with native LDL or PBS at any point of the study (Fig. 2B).
Immunoglobulin M immune response in mice immunized with carbamyl-msLDL
Plasma immunoglobulin M (IgM) dilution curves for one mouse from each immunization group are shown in Figure ID-F. IgM dilution curves for carbamyl-LDL, native LDL, oxidized LDL, and MDA-LDL were similar in a mouse immunized with carbamyl-msLDL (Fig. 1D), msLDL (Fig. 1E), or PBS (Fig. 1F). The highest plasma IgM levels in all three mice were against MDA-LDL.
Plasma IgM levels to carbamyl-LDL, native LDL, oxidized LDL, and MDA-LDL in all mice are shown in Figure 2A. IgM levels to all antigens increased during the study in all mice regardless of the immunizing agent (Fig. 2A). Plasma IgM levels to MDA-LDL were higher than IgM levels to other antigens in all mice. IgM levels were especially high at the end of the study, after the HFD, even in PBS-immunized mice, which suggests that IgM response did not result from immunizations but from other factors, for example, age, or HFD. Plasma IgM response to carbamyl-BSA was also tested. The IgM antibodies binding to carbamyl-BSA increased during HFD in all mice, similarly to the IgM antibodies to LDL-antigens (Fig. 2B). The highest increase was observed in mice immunized with carbamyl-msLDL. However, there was no IgM binding to carbamyl-BSA before introduction of the HFD in any of the mice (Fig. 2B).
IgG immune response to carbamylated LDL is specific in carbamyl-msLDL-immunized mice
The specificity of plasma IgG antibody binding to carbamyl-LDL in mice immunized with carbamyl-msLDL was tested in liquid-phase competitive immunoassay in the presence of increasing amounts (0–200 μg/ml) of competitors (Fig. 3A). Mouse plasma IgG binding to immobilized carbamyl-LDL was competed out with carbamyl-LDL (50%–75% reduction, mean 64%, Fig. 3A), suggesting that the IgG binding to carbamyl-LDL was specific. We also tested the specificity of plasma IgG binding to native LDL, oxidized LDL, and MDA-LDL. Figure 3A shows that native LDL and oxidized LDL did not compete for binding in the liquid phase, whereas MDA-LDL demonstrated competition for IgG binding to immobilized carbamyl-LDL (6%–47% reduction, mean 29%).
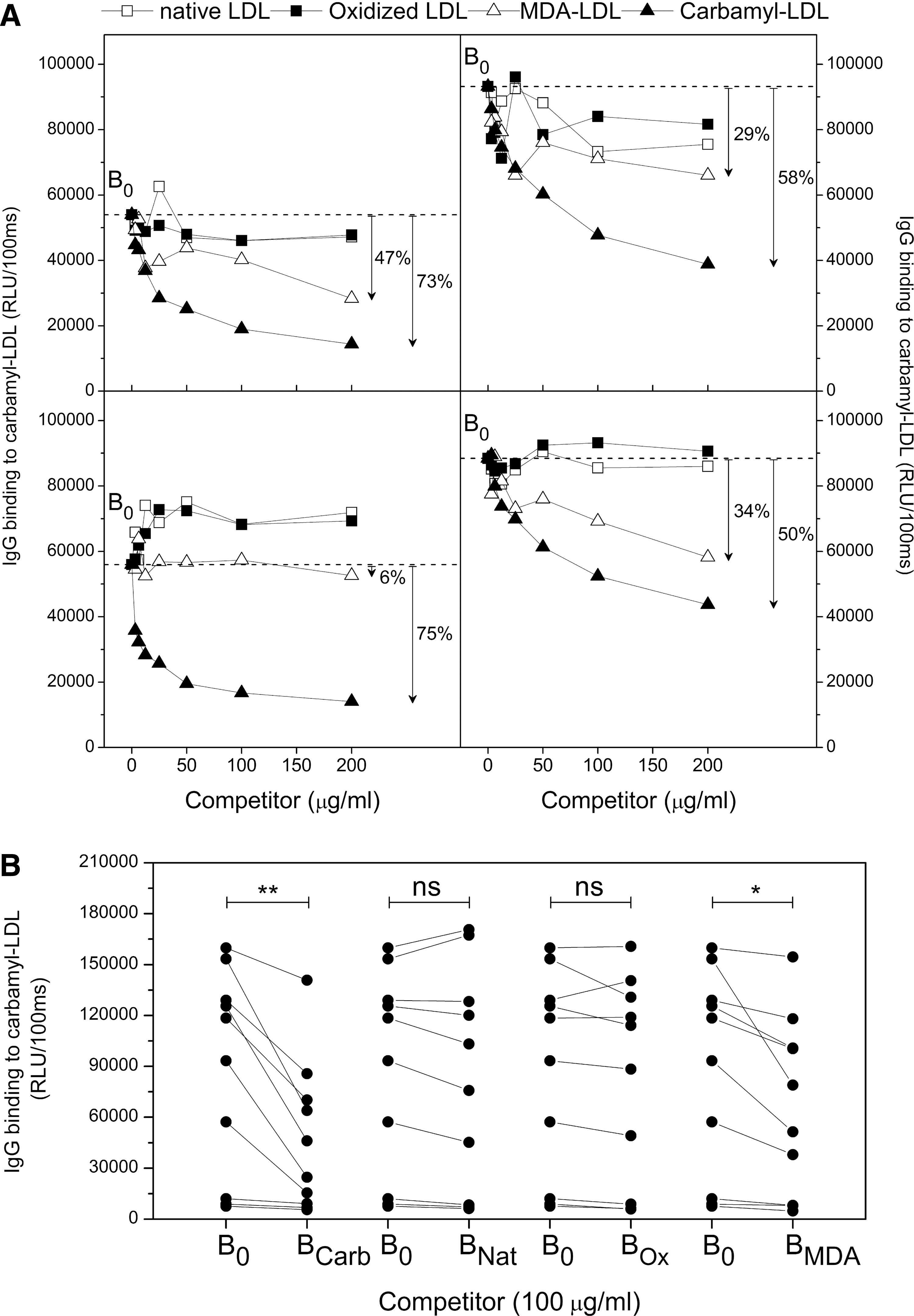
The IgG binding specificity to immobilized carbamyl-LDL was further tested in all carbamyl-msLDL-immunized mice (Fig. 3B). In this assay a fixed amount of carbamyl-LDL, native LDL, oxidized LDL, or MDA-LDL (100 μg/ml) was added to each plasma sample and IgG binding to immobilized carbamyl-LDL was measured. Carbamyl-LDL competed out the plasma IgG binding to immobilized carbamyl-LDL by 43% (mean, p=0.004) and MDA-LDL by 25% (mean, p=0.021) when compared to plasma IgG binding to immobilized carbamyl-LDL without competitor (Fig. 3B). Oxidized LDL or native LDL did not compete for IgG binding to immobilized carbamyl-LDL in mice immunized with carbamyl-msLDL. Mice immunized with native msLDL or PBS did not have IgG antibody responses to carbamyl-LDL or oxidized forms of LDL; therefore, the IgG specificities were not tested.
The specificity of IgM immune response in mice immunized with carbamyl-msLDL is similar to mice immunized with msLDL or PBS
The specificity of plasma IgM antibody response to carbamyl-LDL was also tested in liquid-phase competitive immunoassay (Fig. 4). The competition curves for plasma samples from PBS (Fig. 4A), msLDL (Fig. 4B), and carbamyl-msLDL (Fig. 4C) -immunized mice were determined. Soluble carbamyl-LDL showed minimal competition for IgM binding to immobilized carbamyl-LDL in all immunized mice regardless of the immunogen.
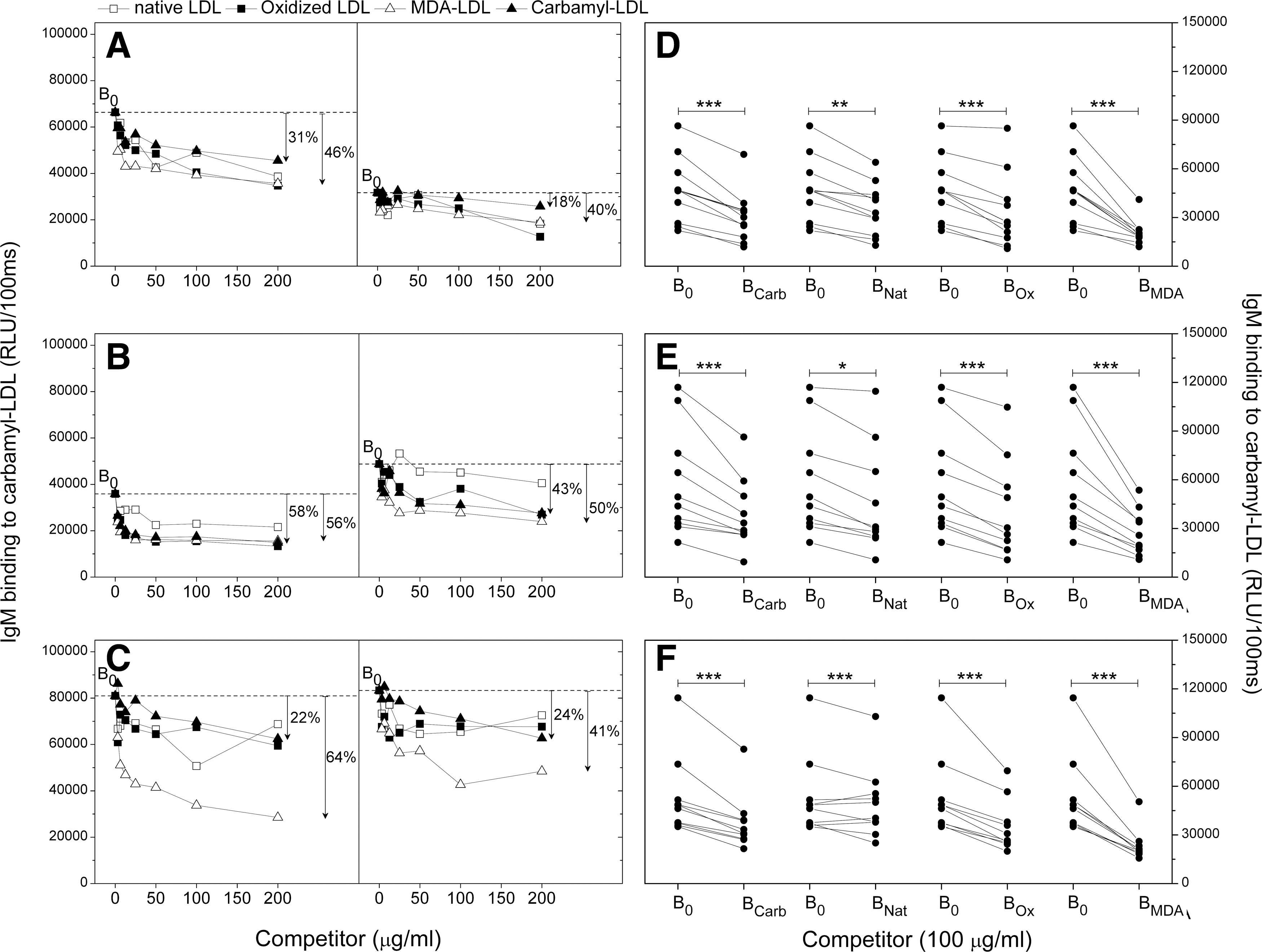
The IgM binding specificity to immobilized carbamyl-LDL was further tested in all mice (Fig. 4D–F). In this assay a fixed amount of carbamyl-LDL, native LDL, oxidized LDL, or MDA-LDL (100 μg/ml) was added to each plasma sample, and IgM binding to immobilized carbamyl-LDL was measured. Carbamyl-LDL, native LDL, oxidized LDL, and MDA-LDL all competed out a statistically significant amount of the plasma IgM binding to immobilized carbamyl-LDL when compared to plasma IgM binding to immobilized carbamyl-LDL without competitor regardless of the immunogen (Fig. 4D–F), suggesting again that IgM response did not result from the immunizations but from other factors, for example, HFD.
Total plasma immunoglobulin levels in mice
An increase in plasma total immunoglobulin levels, both IgG and IgM, was observed in all mice regardless of the immunization agent (Fig. 5A, B). The plasma IgG specificity (see Fig. 3B) showed that on average 43% of the binding was accounted by specific IgG binding to carbamyl-LDL, suggesting that more than half was nonspecific. Therefore, we wanted to normalize the IgG binding to carbamyl-LDL, native LDL, oxidized LDL, or MDA-LDL by plasma total IgG levels (Fig. 5C–F). After normalization, IgG binding to carbamyl-LDL was significantly higher in the carbamyl-msLDL-immunized mice compared to the mice immunized with msLDL or PBS (Fig. 5C). There were no differences in IgG binding to native LDL (Fig. 5D), oxidized LDL (Fig. 5E), or MDA-LDL (Fig. 5F) between mice immunized with carbamyl-msLDL, msLDL, or PBS.
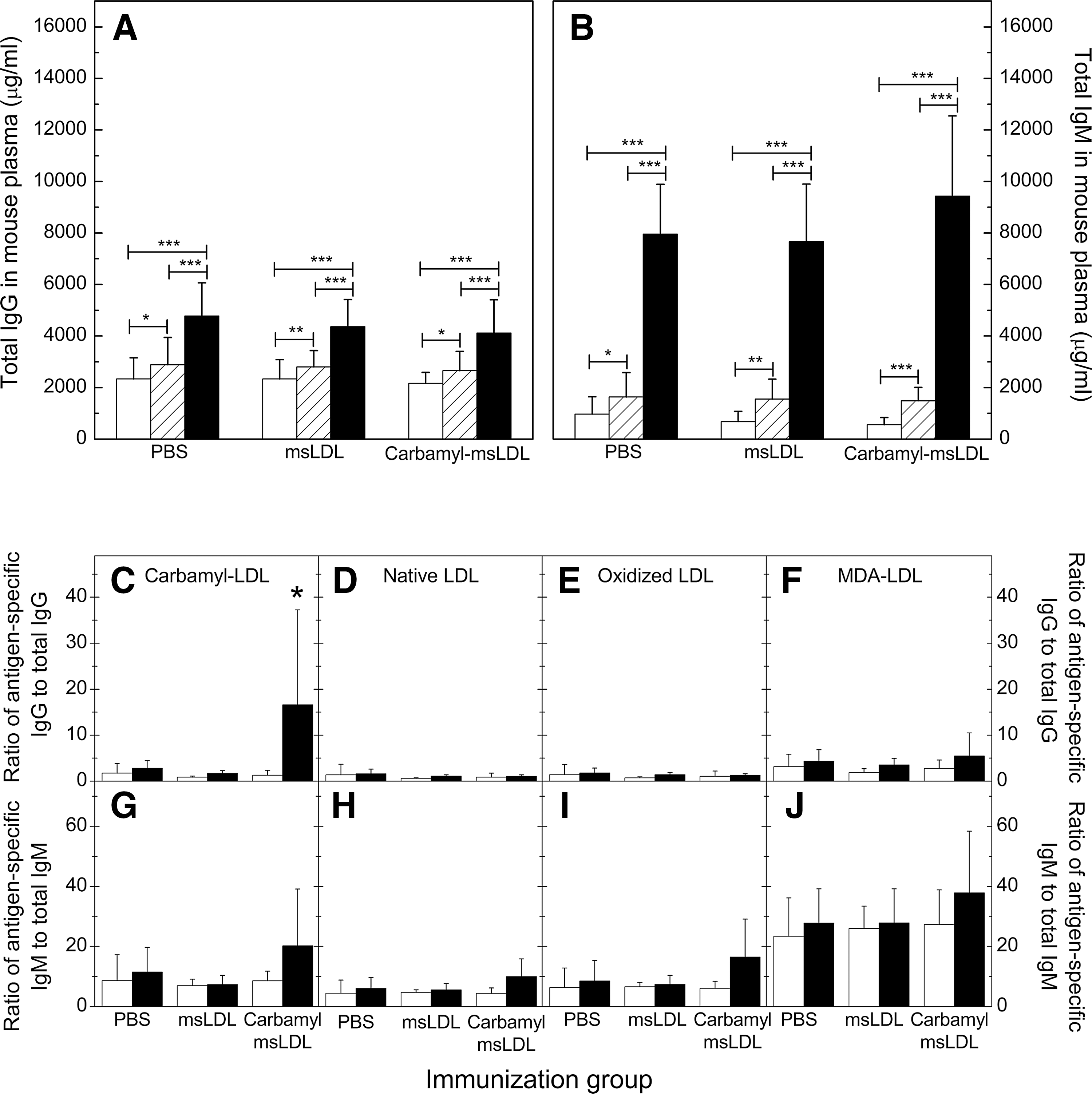
IgM levels to carbamyl-LDL, native LDL, oxidized LDL, or MDA-LDL were normalized (15) similarly by the total plasma IgM levels (Fig. 5G–J). After normalization, there were no differences in IgM binding to any antigen between samples taken at different times, or between the three immunization groups (Fig. 5G–J).
Plasma lipid and urea levels increased in all mice during the study
Total cholesterol (Fig. 6A), high-density lipoprotein (HDL) cholesterol (Fig. 6B), triglyceride (Fig. 6C), and urea (Fig. 6D) levels increased during the HFD, but there were no statistically significant differences between the three immunization groups.

Carbamyl- and MDA-epitopes in mouse plasma
All immunized LDLR
−/− mice had high plasma urea levels after HFD (Fig. 6D); therefore, the amount of carbamylated proteins also in nonimmunized LDLR
−/− mice was investigated. Figure 7A shows that the amount of carbamyl-epitopes on plasma apolipoprotein B100 (apoB100)-containing particles was higher in LDLR
−/− mice fed HFD (n=17, HFD) compared to genetic background control C57BL/6 mice (n=26) (p<0.001, Fig. 7A). In contrast, LDLR
−/− mice fed with normal chow diet (n=21 chow diet) had less carbamyl-epitopes than LDLR
−/− mice fed with HFD (p<0.001, Fig. 7A). Similar results were obtained for carbamyl epitopes in plasma proteins (Fig. 7A). The amount of carbamyl-epitopes in plasma apoB100-containing particles and in plasma proteins measured from LDLR
−/− mice immunized with carbamyl-msLDL, native msLDL, and PBS are shown in Figure 7B. Mice immunized with carbamyl-msLDL had the highest level of carbamylated epitopes after HFD (Fig. 7B). To investigate further the cause for increased amount of carbamyl-LDL in mice immunized with carbmyl-LDL (Fig. 7B), the plasma MPO activity was measured and macrophage uptake assay was performed with rabbit anti-homocitrulline-IgG as described in Supplementary Methods (Supplementary Data are available online at
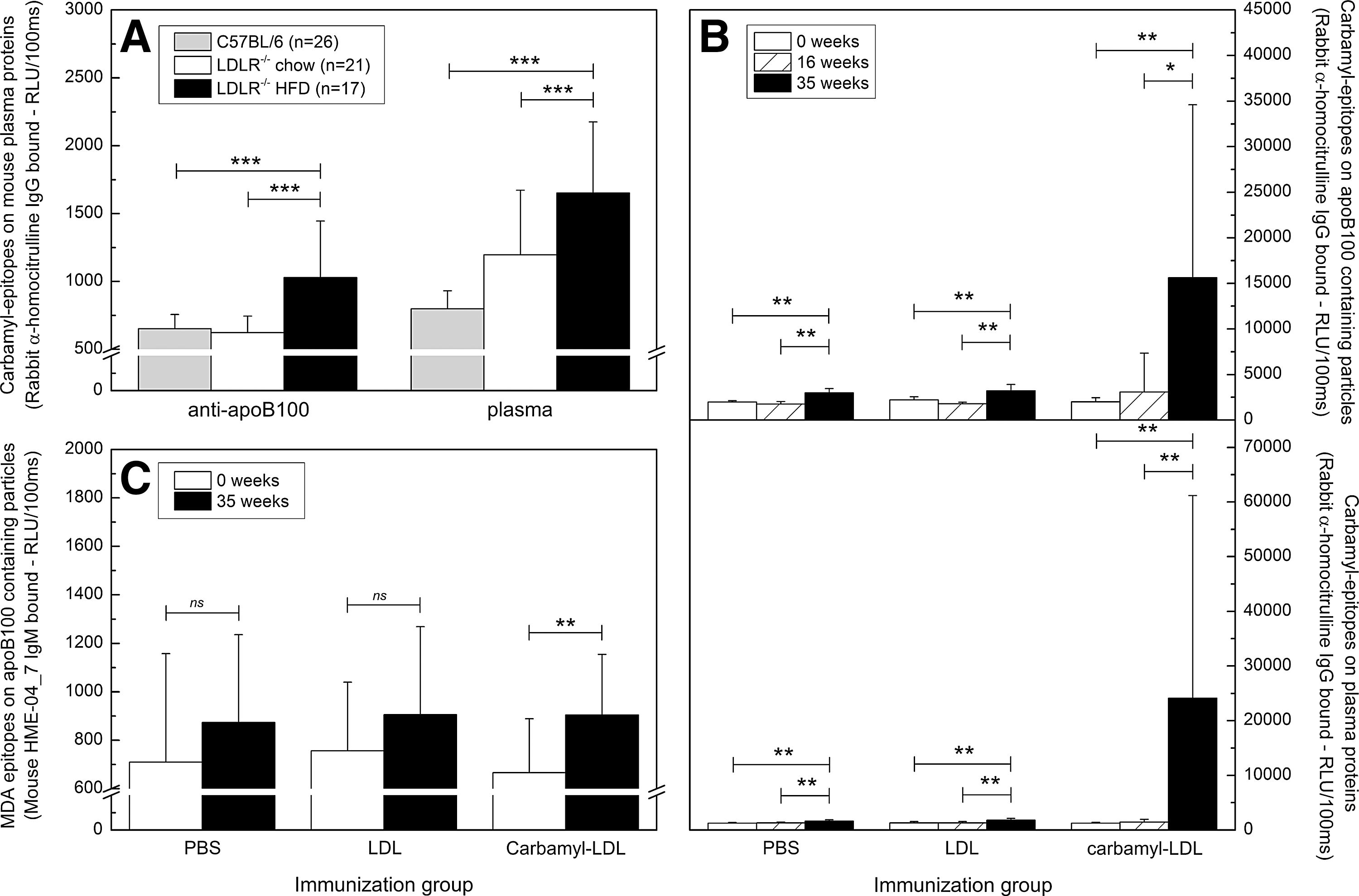
Plasma total cholesterol, HDL cholesterol, and triglyceride levels increased in all mice during the study, as shown in Figure 6A–C. Therefore, the amount of oxidized LDL, measured as MDA-epitopes on plasma apoB-containing particles, was detected in mouse plasma samples. Figure 7C demonstrates that the amount of MDA-epitopes in mice immunized with carbamyl-msLDL was significantly higher after HFD than in the beginning of the study. The MDA-epitopes increased also in mice immunized with LDL and with PBS, but the differences were not statistically significant.
Carbamyl-msLDL immunization does not modulate atherosclerotic lesion size in LDLR−/− mice
The total plaque areas in the cross sections of the aortic origin (Fig. 8A) and in the en face mouse aortas (Fig. 8B) were similar in all groups, although in mice immunized with carbamyl-msLDL, the plaque density was decreased (p=0.036) compared to mice immunized with PBS (0.31±0.11 plaques/mm2 and 0.54±0.35 plaques/mm2, respectively) (data not shown). An aorta from each immunization group is shown in Figure 8C–E.
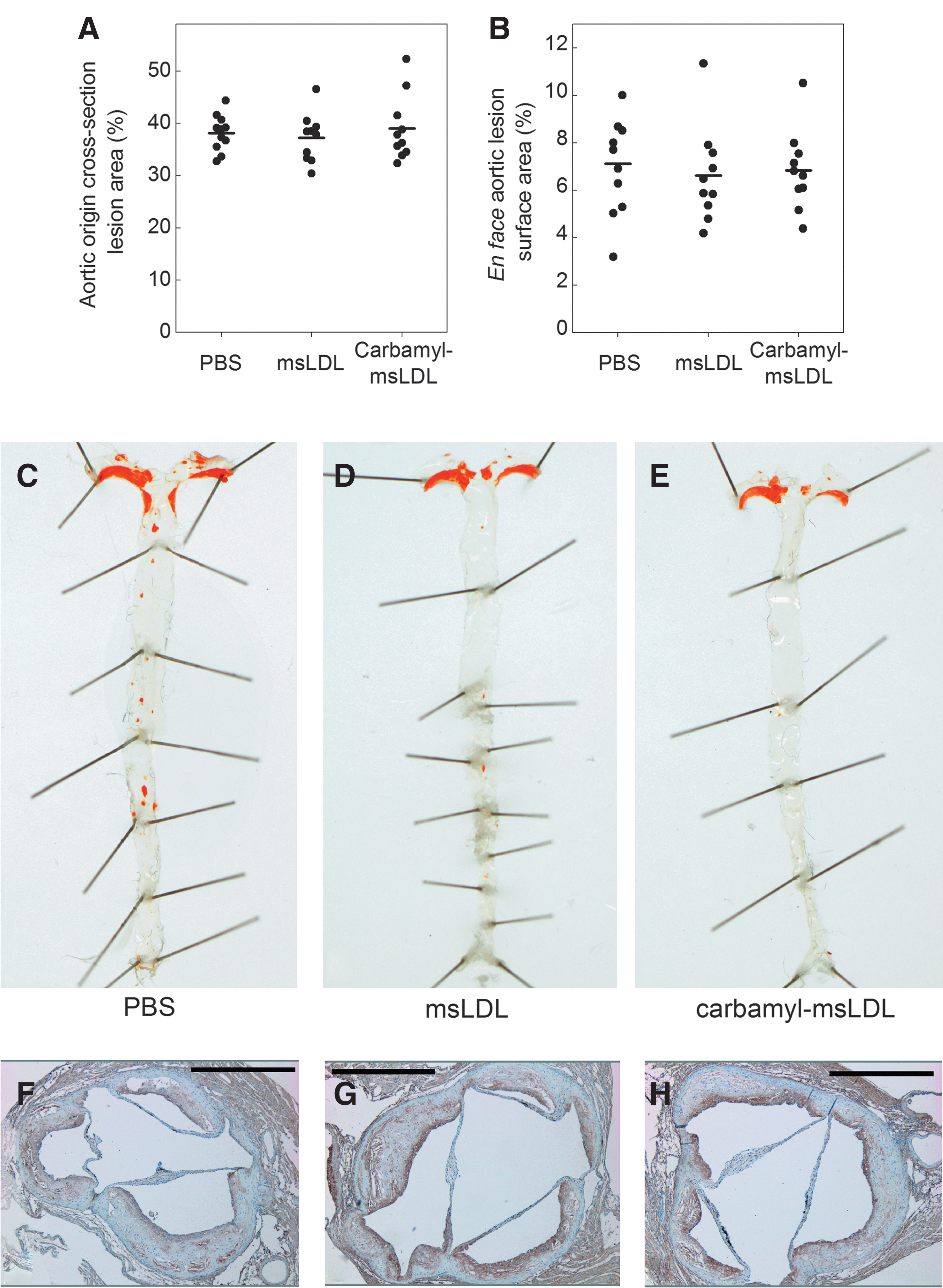
Carbamyl-epitopes are present in atherosclerotic plaques of LDLR−/− mice
Carbamyl-epitopes were present in atherosclerotic lesions when detected with rabbit anti-homocitrulline antibody (43) on aortic origin cross sections of LDLR −/− mice immunized with carbamyl-msLDL, native msLDL, or PBS. Representative cross sections of one mouse in each group are shown in Figure 8F–H. There were no differences in the proportional areas of anti-homocitrulline staining in the atherosclerotic plaques between the immunization groups as detected with MCID Core 7.0 image analysis software (InterFocus Imaging Ltd., Cambridge, United Kingdom).
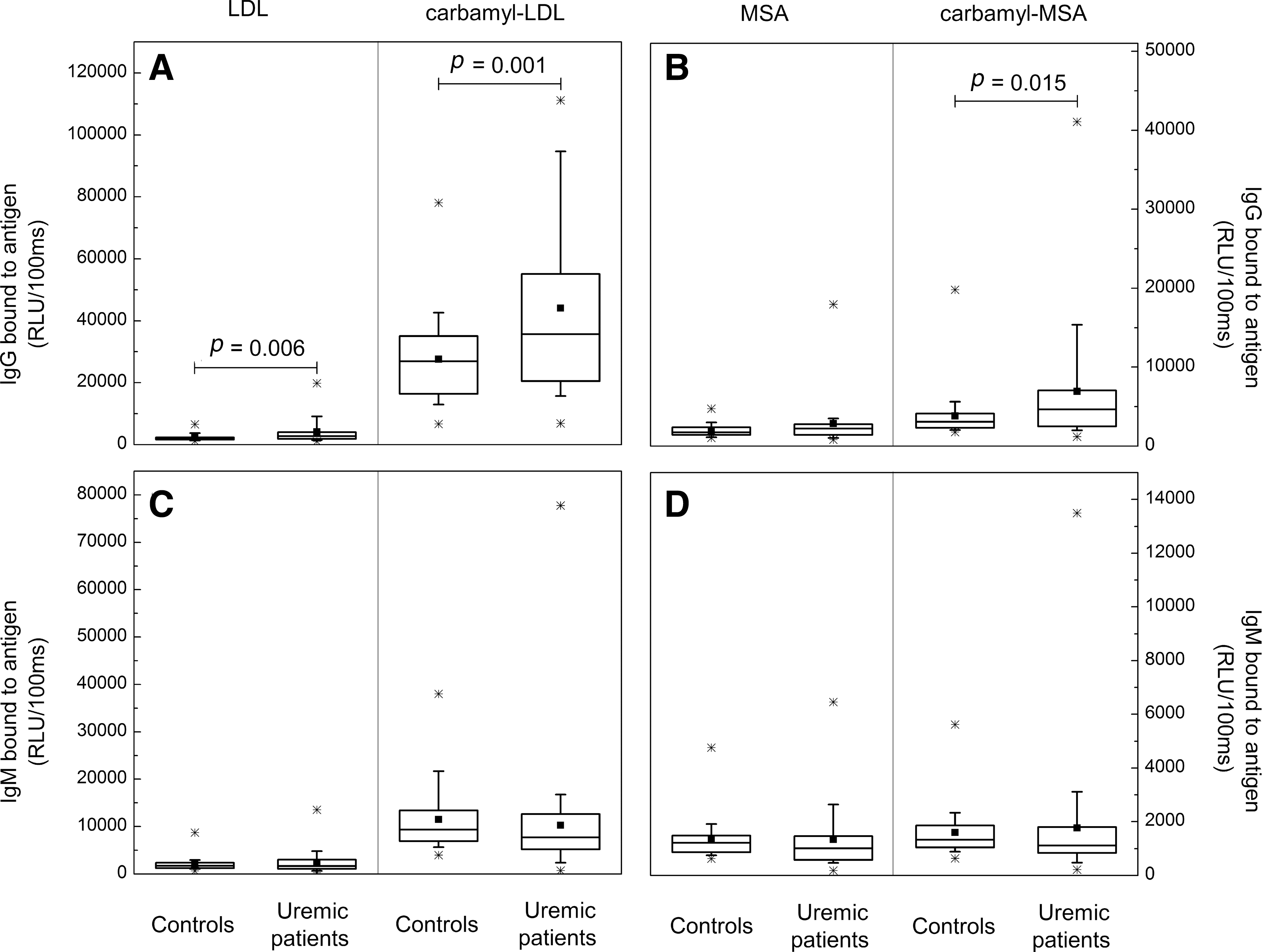
Human plasma contains autoantibodies binding to carbamyl-LDL and carbamyl-mouse serum albumin
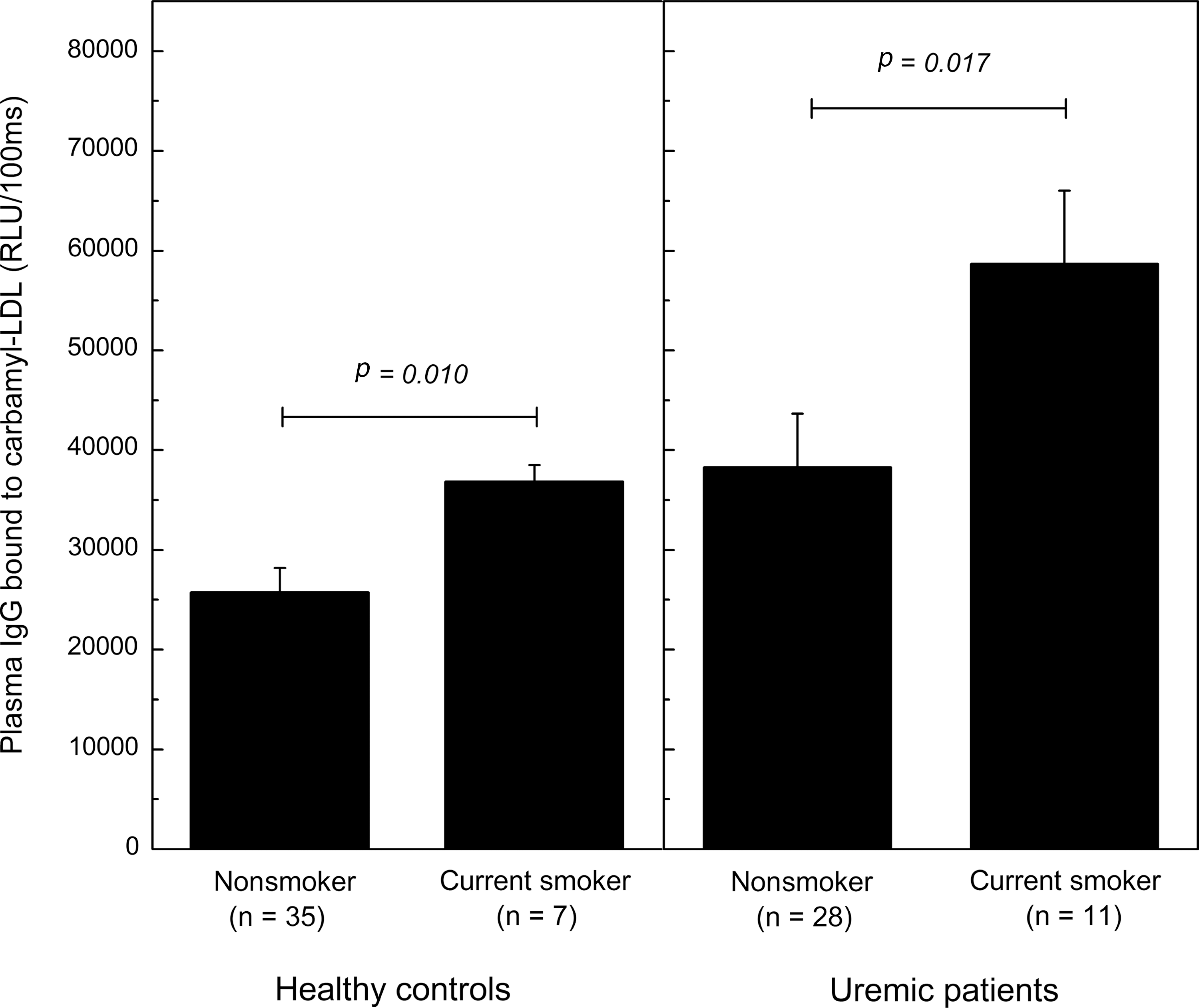
Uremic patients, as well as smoking subjects, are known to have enhanced carbamylation of plasma proteins. Therefore, plasma IgG and IgM antibodies binding to carbamyl-LDL and carbamyl-mouse serum albumin (MSA) were measured from 39 uremic hemodialysis patients and 42 controls with normal plasma urea levels. Eighteen of the study subjects were current smokers. The IgG antibody levels to carbamyl-LDL (Fig. 9A) and to carbamyl-MSA (Fig. 9B) in uremic patients were significantly higher compared to those of the controls (p=0.001 for carbamyl-LDL and p=0.015 for carbamyl-MSA). There were no differences in plasma IgM antibody levels between uremic patients and control subjects (Fig. 9C, D). The IgG antibody levels to carbamyl-LDL were significantly higher in the healthy smoking controls compared to the nonsmokers (Fig. 10A) (p=0.010). In addition, the IgG antibody levels to carbamyl-LDL were significantly higher in the smoking uremic patients compared to the nonsmoking ones (Fig. 10B) (p=0.017). Among all study subjects (n=81) the plasma urea levels associated with plasma IgG to carbamyl-LDL (Spearman's ρ=0.27, p=0.014) and IgG to carbamyl-MSA (Spearman's ρ=0.264, p=0.017).
Discussion
Immune responses to modified LDLs are suggested to be involved in atherogenesis (2, 9, 26). Carbamylation (homocitrullination) is a post-translational modification of importance in the context of uremia, smoking, inflammation, and atherogenesis. The present study investigated the potential role of immunization with carbamyl-modified LDL to elicit humoral immune response and adaptive IgG antibody generation in vivo, and addressed the question if this influenced the course of atherogenesis in LDLR −/− mice during HFD. To further investigate these findings in humans, we found increased plasma IgG immunoglobulin levels to carbamyl-modified proteins in uremic patients and smoking subjects, both known to have increased load of protein carbamylation in vivo.
Previously, vaccination approaches to influence atherogenesis in animal models have been widely studied, and various different chemical modifications of LDL are known to ameliorate the disease process (3, 18, 31, 35, 49). Here, immunization with carbamyl-modified LDL without adjuvant triggered adaptive humoral immune response in LDLR −/− mice, as previously shown in guinea pigs (40), and our starting hypothesis was that this would further, for example, influence the uptake of modified proteins by macrophage scavenger receptors, and subsequently play a role in atherogenesis during HFD. There are no previous reports assessing the functional role of carbamyl-msLDL immunization in atherosclerosis using animal models. We postulated that the humoral immune system may be involved via carbamyl-epitope itself, or via other modifications of LDL generated when the antigen is taken up by dendritic cells, such as those generated from oxidation of polyunsaturated fatty acid-containing lipids in LDL, for example, oxidation-specific epitopes of LDL. We found that immunization of LDLR −/− mice with carbamyl-LDL did not influence the development of total atherosclerotic lipid deposits in en face aortas or atherosclerotic lesion areas in mouse aortic origin cross sections.
To investigate the functional role of humoral immune response more in detail, we demonstrated that carbamyl-LDL immunization did not induce IgG immune response to native LDL or to oxidized LDL, but instead a minimal and specific IgG antibody response to MDA-LDL. Proteins haptenated with MDA are immunogenic and vaccination with MDA-LDL ameliorates atherosclerosis in LDLR −/− mice (18). Serum MDA (11, 41) and MDA-modified LDL (MDA-LDL) (45) levels are increased in humans with enhanced atherosclerosis. In a competitive immunoassay, soluble MDA-LDL was able to compete out approximately one fourth of the plasma IgG binding to immobilized carbamyl-LDL, suggesting some degree of cross-reactive epitopes between carbamyl-LDL and MDA-LDL being created after the carbamyl-LDL immunization. No comparable cross-reactivity was observed between IgG binding to carbamyl-LDL and oxidized LDL, or native LDL. One possible explanation is that when in tissue (after immunization) carbamyl-LDL is internalized by the antigen presenting cells, and LDL lipids further undergo oxidation to create MDA-epitopes. This was supported by the increased amount of MDA-LDL measured in carbamyl-LDL-immunized mice, but not PBS- or LDL-immunized mice. Furthermore, we did not observe IgG response to MDA-LDL in mice immunized with PBS or native LDL.
A clear IgM immune response to MDA-LDL, carbamyl-LDL, oxidized LDL, and native LDL was observed in all study mice at the end of the study. This IgM response was observed also in mice immunized with PBS and native LDL, suggesting that it was caused by, for example, HFD in this LDLR −/− mouse strain. These data are well in line with the previously published results for total IgM in mice immunized with oxidized LDL (37). In mice, most natural antibodies are of the IgM isotype and oxidation-specific epitopes are proposed to be one of the principal targets (15). The total IgM immunoglobulin levels were increased during the study in all mice, and there were no differences between the three immunization groups, demonstrating that this was unlikely to influence the atherosclerosis results.
Majority of the immunization studies assessing the functional role of modified LDL in atherogenesis in animal models have used Freund's adjuvant to stimulate the immune reactions (18, 35, 49). Freund's adjuvant has been documented to ameliorate atherosclerosis in apoliprotein E-deficient (apoE −/−) mice (24), and one of the mechanisms proposed included increased plasma IgM levels to MDA-LDL. In New Zealand White (NZW) rabbits, however, Freund's adjuvant has been shown to enhance the lesion development (47). We omitted Freund's adjuvant in the present immunizations of LDLR −/− mice, and observed as previously in guinea pigs (40), that modified self-lipoproteins are particularly immunogenic, and can induce adaptive humoral IgG immune response, even in the absence of any adjuvant. The potential confounding effects of the use of Freund's adjuvant on atherogenesis in animal models have often been overlooked; therefore, it is challenging to compare our results with the previous reports using other modifications of LDL.
Carbamylated LDL possesses various atherogenic properties, such as the capacity to increase vascular smooth muscle cell proliferation and monocyte adhesion via overexpression of intercellular adhesion molecule-1 (ICAM-1) and vascular cell adhesion molecule-1 (VCAM-1) (8). These are likely to contribute to the enhanced atherosclerosis in renal patients. Carbamylated LDL has also been suggested to increase atherosclerotic disease in patients with CKD by inducing oxidative stress and accelerating senescence in human endothelial progenitor cells (13). Carbamyl-LDL binds to lectin-type oxidized LDL receptor 1 (LOX-1) and CD36 scavenger receptors, whereas the transcytosis involves SR-A1, SREC-1, but not LOX-1 receptors (7). Carbamylation was also shown to be the major post-translational modification of HDL influencing its antiatherogenic properties (20). A recent study suggests that cyanate, rather than carbamylated lipoproteins, induces ICAM-1 expression in human coronary artery endothelial cells, and when administered orally it induces endothelial ICAM-1 expression in mouse aorta (17). We used IgG from NZW rabbits immunized with homocitrulline-containing synthetic peptide (CEKAHDGGRYHomocitA) (43) to assess the amount of carbamylated lysine-epitopes in apoB-containing lipoproteins in LDLR −/− mice. In the present study, LDLR −/− mice on regular chow diet had similar amounts of carbamylated LDL compared to C57BL/6 mice. However, during HFD the amount of carbamyl-LDL in LDLR −/− increased significantly, possibly due to increase in serum urea levels. The plasma levels of carbamyl-LDL in LDLR −/− mice immunized with carbamyl-LDL were significantly higher compared to PBS and native LDL-immunized mice. In addition, plasma MPO activity was significantly increased, which might explain the increased amount of carbamyl-LDL in these mice. The levels of plasma carbamyl-LDL did not associate with the plasma IgG antibody titers to carbamyl-epitopes, suggesting that immune complexes are not likely to play a mechanistic role. Carbamyl adducts have been identified previously in human atherosclerotic lesion (46). In the present study, all mice had atherosclerosis and elevated plasma urea levels at the end of the study regardless of the immunizing agent. It was no surprise that they all had carbamyl-epitopes present in the atherosclerotic lesions shown by immunohistochemical staining, but there were no significant differences in proportional stained areas between the mice immunized with carbamyl-msLDL, native ms-LDL, or PBS.
One work, so far, has demonstrated autoantibodies to carbamylated (homocitrulline-containing) proteins in humans, and the study suggested that IgG anti-carbamyl antibodies are predictive for a more severe course of rheumatoid arthritis (38). We demonstrated increased IgG, but not IgM, antibody levels to carbamylated LDL and carbamylated albumin in uremic subjects, as well as in smokers. The highest antibody levels to carbamyl-LDL were among smokers with elevated plasma urea levels, and the lowest antibody levels were found in healthy nonsmokers. The absence of IgM antibodies indicates that the immune response is likely to be secondary, not primary, in nature. To our knowledge, there are no data suggesting that the IgG immunoglobulin levels in these patients would be increased simply by kidney disease-specific factors or medications. Quite the contrary, the patients may have lower levels of total immunoglobulin, and humoral immune response to certain pathogenic antigens has been shown to be lower/diminished (14, 16, 30). Diminished antibody production in patients with end-stage renal disease is due to impairment of T-cell activation, reduction in number of B lymphocytes, and dendritic cells (the major antigen presenting cells) (1, 34). Protein carbamylation may be used as a marker to predict increased coronary artery disease risk, and the amount of systemic ɛ-carbamyllysine is suggested to be used as a quantitative index of cardiovascular risk (46). The measurement of homocitrulline levels from, for example, plasma samples, however, is laborious and requires stable–isotope-dilution high-performance liquid chromatography (HPLC) with online tandem mass spectrometry (46). Our data suggest that the IgG type plasma antibodies to carbamyl adducts may indicate increased homocitrulline levels, and potentially should be further investigated also as markers of cardiovascular disease in subjects with increased carbamylative burden.
In conclusion, we showed that immunization of mice with carbamyl–haptenated msLDL resulted in specific IgG response to carbamyl-LDL and MDA-LDL. The immunization did not influence the development of atherosclerosis in LDLR −/− mice during high-fat feeding. We demonstrated that humans had specific IgG autoantibodies binding to carbamylated proteins, and these antibody levels were increased in conditions known to have enhanced carbamylation, for example, uremia and smoking. The existence of human autoantibodies binding to carbamyl-LDL in vivo gives new insights into mechanisms leading to recognition of post-translationally modified structures, which are suggested to play a role in conditions leading to enhanced carbamylation, such as in uremia and smoking.
Materials and Methods
LDL isolation and modifications
LDL was isolated by sequential ultracentrifugation from pooled plasma of LDLR −/− mice (msLDL for immunizations) and from healthy human donors (antigen for immunoassays). To exclude the possibility of lipid oxidation, antioxidants (20 μM butylated hydroxytoluene [BHT] and 0.27 mM ethylenediaminetetraacetic acid [EDTA]) were added to the freshly isolated LDL preparation before the modifications. LDL was in vitro carbamylated with potassium cyanate (KCNO) as previously described with slight modifications (23). In brief, 2 mg of LDL was diluted with 0.3 M Na2B4O7, pH 8.0 to 1.5 times the original volume, and 20 mg of KCNO was added per 1 mg of LDL protein. LDL was carbamylated at +37°C for 6 h. Albumin was carbamylated similarly for 24 h (antigen for immunoassays). This in vitro carbamylation method results in heavily carbamylated antigens that may not be physiological in vivo. After carbamylation, the excess cyanate was removed by dialyzing against 0.27 mM EDTA–PBS. To verify the presence of homocitrulline residues, the native and carbamylated protein preparations (LDL and albumin) were hydrolyzed in 6 M HCl at 110°C for 16 h, freeze-dried, and analyzed with HPLC (43) (Supplementary Fig. S2). To further analyze the amount of homocitrulline in carbamyl-BSA and carbamyl-LDL, the amino acid analysis was performed as described in Supplementary Methods and the data are shown in Supplementary Figure S3. Carbamyl-LDL preparations were also tested for the absence of thiobarbituric acid reactive substances (TBARS) (48). Copper oxidized LDL (oxidized LDL) and MDA-LDL were prepared as described previously (36).
Mice and immunizations
All animal studies were conducted by protocols approved by the National Animal Experiment Board, Finland (STH1164A, 21.11.2007). Thirty-one female LDLR −/− mice (C57BL/6J background B6.129S7-Ldlr tm1Her ) were immunized subcutaneously (s.c.) with 50 μg carbamylated msLDL (carbamyl-msLDL, n=10), native msLDL (native-msLDL, n=10) or phosphate-buffered saline (PBS; n=11). Booster immunizations containing 25 μg of the same antigens were given intraperitoneally (i.p.) at weeks 3, 6, 12, and 16. During the first 16 weeks of the study mice were fed regular chow (4.4% fat and 0.01% cholesterol). Mice were fed Western type HFD (Harlan Teklad TD.88137; containing 21.2% fat and 0.2% cholesterol) from week 17 onwards. Three booster immunizations (25 μg) were given (i.p.) once a month during the HFD (weeks 20, 24 and 29). Freund's adjuvant has been documented to ameliorate atherosclerosis in animal models (24), and therefore, all immunizations were carried out without any adjuvant. Blood samples were taken at the beginning of the study (week 0), after immunizations, before HFD (week 16) and after HFD (week 35). The mice were sacrificed between weeks 36 and 37 and the blood was collected from the posterior vena cava (week 37).
Human study subjects
Blood samples were collected from 39 (12 females and 27 males) uremic patients with end-stage chronic renal failure (CKD stage 5D). Patients were regularly visiting the hemodialysis unit of the Department of Internal Medicine at Oulu University Hospital. The mean age of the uremic patients was 59±18 (mean±SD) years and the mean plasma urea (predialysis) was 19.0±4.5 mM. Control samples were collected from 42 healthy subjects (26 females and 16 males), aged 36±9 years, with normal plasma urea levels and renal function (plasma urea 5.2±1.2 mM). Clinical characteristics of the hemodialysis patients and the control subjects are represented in Supplementary Table S1. The causes for renal failure and the patients' medications are listed in Supplementary Table S2. The uremic patients were on average considerably older than the control subjects. In addition, the gender distribution was different and the proportion of smokers was larger among the uremic patients than among the control subjects. The mean plasma total cholesterol and LDL cholesterol and also plasma albumin were somewhat lower in uremic patients than in controls. Fasting blood glucose level was higher among uremic patients than among control subjects. In addition to decreased renal function, age and smoking may also influence carbamylation. All study subjects gave informed written consent for the investigations. The study was approved by the Ethical Committee of Oulu University Hospital, Finland (21/2006, 20.3.2006), and followed the Declaration of Helsinki.
Chemiluminescent immunoassays
Plasma antibody levels were determined using chemiluminescent immunoassay method as described (44). For determination of antibody levels in mouse and human plasma samples, antigens or capture antibodies were immobilized on microtiter plate (Nunc ImmunoFluor2; Thermo Fisher Scientific, Inc.) 5 μg/ml in PBS −0.27 mM EDTA overnight at +4°C. Mouse plasma samples were diluted 1:50–1:3200 for plasma dilution curves, 1:1000 for determination of antigen-specific antibody levels, and 1:50000 for determination of plasma total IgG and IgM levels. Human plasma samples were diluted 1:500. The plasma samples were incubated on antigen, or antibody, coated plate for 1 h at room temperature and the amount of bound antibodies was measured with alkaline phosphatase labeled secondary antibodies (anti-mouse-IgG, anti-mouse-IgM, anti-human-IgG and anti-human-IgM) (Sigma-Aldrich). LumiPhos 530 (Lumigen, Inc.) was used as substrate and the chemiluminescence was measured with Wallac Victor3 multilabel counter (Perkin Elmer). The results are expressed as relative light units measured in 100 ms (RLU/100 ms). The specificity of mouse antibody binding to carbamylated LDL was verified using liquid-phase competition immunoassay (32). Plasma samples (1:1000) were incubated overnight at +4°C in the presence and absence of competitors (0–200 μg/ml). The immunocomplexes were pelleted by centrifugation for 30 min 16,000 g at +4°C and antibodies remaining in the liquid phase were analyzed using chemiluminescent immunoassay described above. Mouse IgG and IgM antibody binding to carbamyl-LDL was tested with carbamylated human LDL and carbamylated msLDL. The results were similar, and carbamylated human LDL was used as immobilized antigen and soluble competitor in immunoassays.
Determination of carbamyl- and MDA-epitopes in mouse plasma samples
The carbamyl-epitopes in mouse plasma samples were determined with chemiluminescent immunoassay described above using rabbit anti-homocitrulline IgG purified with Protein G column (Pierce/Thermo Fisher Scientific) from sera of immunized rabbit. The immunization was performed with homocitrulline-containing synthetic peptide (CEKAHDGGRYHomocitA) as described earlier (43). In this assay, anti-human apoB100/48 (Meridian Life Sciences) was used 5 μg/ml to capture LDL particles from mouse plasma samples (1:200), or proteins in plasma samples were immobilized directly onto 96-well plates. Captured LDL particles were detected with biotinylated rabbit anti-homocitrulline-IgG 3 μg/ml and alkaline phosphatase-labeled NeutrAvidin (Thermo Fisher Scientific) and Lumiphos 530 substrate. MDA epitopes on captured apoB-containing particles were determined similarly, except mouse plasma samples were diluted 1:1000 and biotinylated mouse IgM specific for MDA-LDL (clone HME-04_7 from naïve apoE −/− mouse fed HFD) (44) was used 1 μg/ml as primary antibody.
Determination of mouse atherosclerosis
After sacrifice, the mice were perfused through the left ventricle with PBS containing 1 μg/ml BHT for 10 min and then perfusion-fixed with formalin-sucrose solution pH 7.4 (10% formalin −5% sucrose–3 μM EDTA–20 μM BHT) for 10 min (36). The en face preparation of the entire aorta and cross sections through the aortic origin were carried out as previously described (44). Mouse aortas were cleaned, stained with Sudan IV (Sigma Aldrich), opened longitudinally, and pinned on paraffin plates. Mouse hearts were embedded in paraffin and 5 μm cross sections were made through the aortic origin. Hematoxylin-eosin stained cross sections and Sudan IV stained aortas were analyzed using QImaging MicroPublisher 5.0 RTV camera attached to the Leitz Aristoplan microscope and MCID Core 7.0 analysis software (InterFocus Imaging Ltd.).
Immunostaining of mouse heart aortic origin cross sections
Aortic origin cross sections from LDLR −/− mice immunized with carbamyl-msLDL, native msLDL, and PBS were stained with rabbit anti-homocitrulline-IgG to localize carbamyl-epitopes in atherosclerotic lesions. Antigen retrieval was performed by a 10-min treatment with 10 mM sodium citrate pH 6.0 near boiling point. The cross sections were stained with EnVision+ System-horseradish peroxidase (HRP) (DAB) for use with rabbit primary antibodies (Dako North America) according to kit protocol. In short, endogenous peroxidase activity was quenched with Peroxidase Block. Rabbit anti-homocitrulline-IgG primary antibody 2 μg/ml or nonimmunized rabbit IgG (data not shown) were applied onto separate tissue sections followed by HRP-labeled polymer conjugated with secondary antibody. Staining was completed with 3,3′-diaminobenzidine (DAB+) substrate chromogen, which results brown-colored precipitate at antigen site. Sections were further counterstained with Mayer's hematoxylin. Images were acquired with Leica DM 3000 using Leica Application Suite V4.1 (Leica Microsystems). For determining the proportional areas of anti-homocitrulline staining, the aortic origin cross sections were lined following the adventitia-media border and within that region, the anti-homocitrulline IgG stained areas were automatically detected with MCID Core 7.0 analysis software (InterFocus Imaging Ltd.).
Determination of mouse plasma lipid and urea levels
Mouse plasma total cholesterol, HDL cholesterol, and total triglycerides were measured by enzymatic methods (Roche Diagnostics) according to the manufacturer's instructions. Mouse plasma urea concentrations were measured using QuantiChrom Urea Assay Kit (BioAssay Systems) according to the kit protocol.
Clinical measurements of human blood samples
Human plasma total cholesterol, HDL cholesterol, LDL cholesterol, triglycerides, creatinine, albumin, urea, glucose, and urate were determined in the laboratory of Oulu University Hospital using standard laboratory methods. Blood samples from uremic patients with end-stage chronic renal failure were collected before dialysis treatment.
Statistical analysis
Data analyses were performed with the software package SPSS Statistics (19.0). The differences between the groups were analyzed by t-test or Mann–Whitney U-test. Different samples from a same mouse were compared using paired samples t-test. p-value <0.05 was regarded statistically significant.
Footnotes
Acknowledgments
The authors thank Ms. Sirpa Rannikko for her excellent technical assistance. These studies were supported by the Academy of Finland, the Finnish Foundation for Cardiovascular Research, the Sigrid Juselius Foundation, the Sohlberg Foundation, the Paulo Foundation, and the Aarne Koskelo Foundation.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.