
Abstract
Introduction
Innovation
Recently, we identified six S-bacillithiolated proteins in Bacillus subtilis in response to NaOCl stress (7). The methionine synthase MetE was inactivated by S-bacillithiolation causing methionine auxotrophy. Here, we have studied the conserved S-bacillithiolome among four different Bacillus species and Staphylococcus carnosus using bacillithiol (BSH)-specific immunoblots, shotgun liquid chromatography–tandem mass spectrometry analysis, and redox proteomics. We identified 54 S-bacillithiolated proteins, including eight conserved proteins (MetE, AroA, GuaB, TufA, PpaC, SerA, YphP, and YumC) and 29 unique proteins involved in amino acid and cofactor biosynthesis, nucleotide metabolism, translation, protein quality control, redox, and antioxidant functions. S-bacillithiolation is accompanied by an increased oxidized bacillithiol disulfide (BSSB) level and a decreased BSH/BSSB ratio. Together, our data support a major role of the BSH redox buffer in redox control and protection of conserved and essential proteins against irreversible oxidation by S-bacillithiolations in Firmicutes bacteria.
Gram-positive bacteria do not produce GSH. Among the actinomycetes, mycothiol is the predominant LMW thiol that serves analogous roles to GSH (20, 39). In low-GC Gram-positive bacteria, such as Bacillus and Staphylococcus species, bacillithiol (BSH) serves as a redox buffer that was structurally characterized as Cys-GlcN-malate (12, 16, 40). Mutants with defects in BSH biosynthesis displayed strong sensitivities to the epoxide antibiotic fosfomycin and to NaOCl stress (7, 12). In Bacillus subtilis, the redox-sensing MarR-type repressor OhrR forms mixed disulfides with BSH (S-bacillithiolations) in response to organic hydroperoxides and NaOCl stress (7, 26). S-bacillithiolation of OhrR alleviates repression of transcription of the ohrA peroxiredoxin gene as a protection mechanism against NaOCl toxicity (7). Besides OhrR, we have recently identified by liquid chromatography (LC)–tandem mass spectrometry (MS/MS) analysis five cytoplasmic proteins with S-bacillithiolations after NaOCl treatment (7). Hypochloric acid shows very fast reaction rates with Cys residues (k 2=3×107 M −1 s −1) and rapidly leads to irreversible oxidation products, such as sulfinic and sulfonic acids (9, 15, 42, 50). Thus, S-bacillithiolation serves to protect active-site Cys residues of key metabolic enzymes against overoxidation. S-bacillithiolation of the methionine synthase MetE at the active site Cys730 caused a methionine starvation phenotype in NaOCl-treated cells, presumably to slow down translation while the OhrA peroxiredoxin removes the toxic oxidant (7).
In this work, we have identified proteome-wide conserved S-bacillithiolations in various industrially important Bacillus and Staphylococcus species. We used BSH-specific immunoblot analyses, redox proteomic approaches, and shotgun-LC-MS/MS analyses to identify essential and conserved proteins in the S-bacillithiolome that have been previously identified in the S-glutathionylome in eukaryotes. The protein bacillithiolations were accompanied by an increased level of oxidized bacillithiol disulfide (BSSB) and a decreased BSH/BSSB redox ratio in B. subtilis.
Results
Defining the conditions of NaOCl stress to induce S-bacillithiolations in the proteome of B. amyloliquefaciens, B. pumilus, B. megaterium, and S. carnosus
To describe more comprehensively the S-bacillithiolome, we focused on four industrially important Bacillus species: B. subtilis 168, B. pumilus SBUG1799, B. amyloliquefaciens FZB42 (5), B. megaterium SBUG1152 (36), and S. carnosus TM300 (43). The B. subtilis, B. pumilus, and B. megaterium strains were able to grow in the Belitsky minimal medium (BMM), and low NaOCl concentrations could be applied to reduce the growth rate: 75 μM NaOCl for B. subtilis (7), 125 μM for B. pumilus, and 80 μM for B. megaterium (Supplementary Fig. S1B, C; Supplementary Data are available online at
MetE is the major target for S-bacillithiolation in Bacillus species as revealed by nonreducing BSH-specific immunoblot analysis
To monitor the extent of S-bacillithiolation after NaOCl stress, BSH-specific antibodies were used to test protein extracts from NaOCl-treated B. subtilis wild-type and bshA-mutant cells in nonreducing immunoblot analyses. The BSH immunoblots showed a very strong 90-kDa band after NaOCl stress, corresponding to MetE-SSB (Fig. 1A). The identity of MetE was confirmed by LC-MS/MS analysis of the specific band from nonreducing sodium dodecyl sulfate (SDS) gels (Supplementary Table S2). S-bacillithiolated MetE, modified at Cys719 and Cys730, was previously identified as the most strongly oxidized protein in the redox proteome of B. subtilis (7). We further confirmed S-bacillithiolation of MetE by comparison of extracts from NaOCl-treated B. subtilis cells grown in the BMM and LB media. In a rich LB medium, MetE is repressed, and thus the MetE-SSB modification is not visible in the BSH immunoblots (Fig. 1B). The MetE-SSB band was also not visible in NaOCl-exposed bshA-mutant cells or in reducing BSH immunoblots (Supplementary Fig. S2). Besides MetE, there were a few weaker bands in the 45–55-kDa range, which represent other protein BSH-mixed disulfides (protein-SSB), since these are absent in the bshA mutant cells. These BSH immunoblot results show that MetE is the most abundant S-bacillithiolated protein in NaOCl-treated B. subtilis cells.
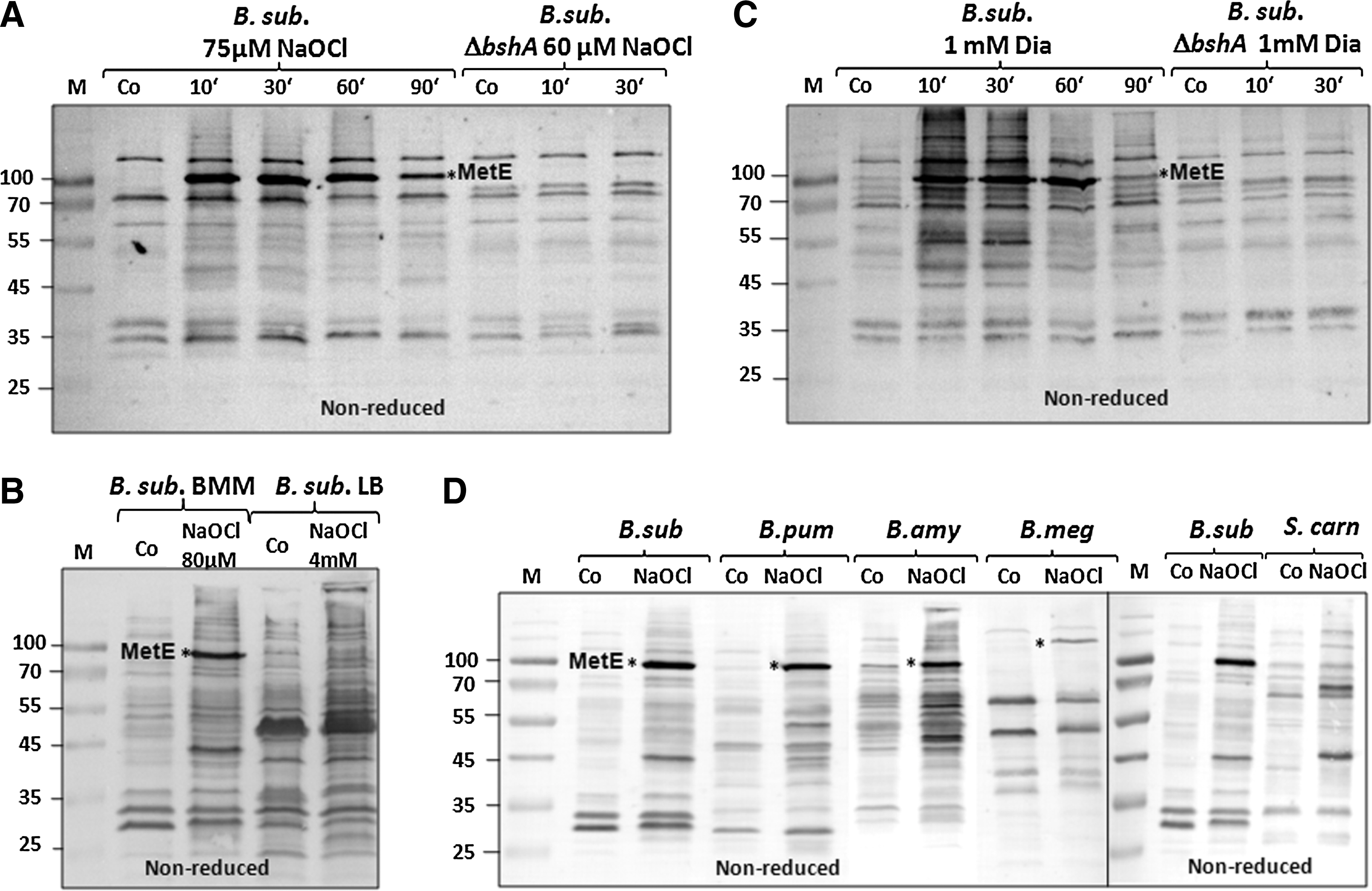
Furthermore, BSH immunoblots indicate that MetE is also the major BSH-modified protein in diamide-treated cells (Fig. 1C). In addition, many other BSH-modified proteins are visible after diamide stress, indicating that S-bacillithiolation is also a significant thiol modification in diamide-treated cells.
Next, we analyzed the S-bacillithiolome of NaOCl-treated cells from B. amyloliquefaciens, B. pumilus, B. megaterium, and S. carnosus (Fig. 1D). The protein extracts of all NaOCl-treated Bacillus strains showed MetE as the most abundant protein-SSB, indicating that S-bacillithiolation of MetE is a common protection mechanism against oxidative stress among Bacillus species. In B. megaterium, MetE-SSB was observed at a higher molecular weight and confirmed by LC-MS/MS analysis (Supplementary Table S2). In S. carnosus grown in LB medium, various protein-SSBs were visible, indicating that NaOCl also causes S-bacillithiolations in Staphylococcus. Because MetE synthesis is strongly repressed in LB medium, MetE-SSB was not detected in S. carnosus using BSH-specific immunoblots.
Identification of proteins with S-bacillithiolations in NaOCl-treated cells of B. subtilis, B. amyloliquefaciens, B. pumilus, B. megaterium, and S. carnosus using redox proteomics and shotgun-LC-MS/MS analyses
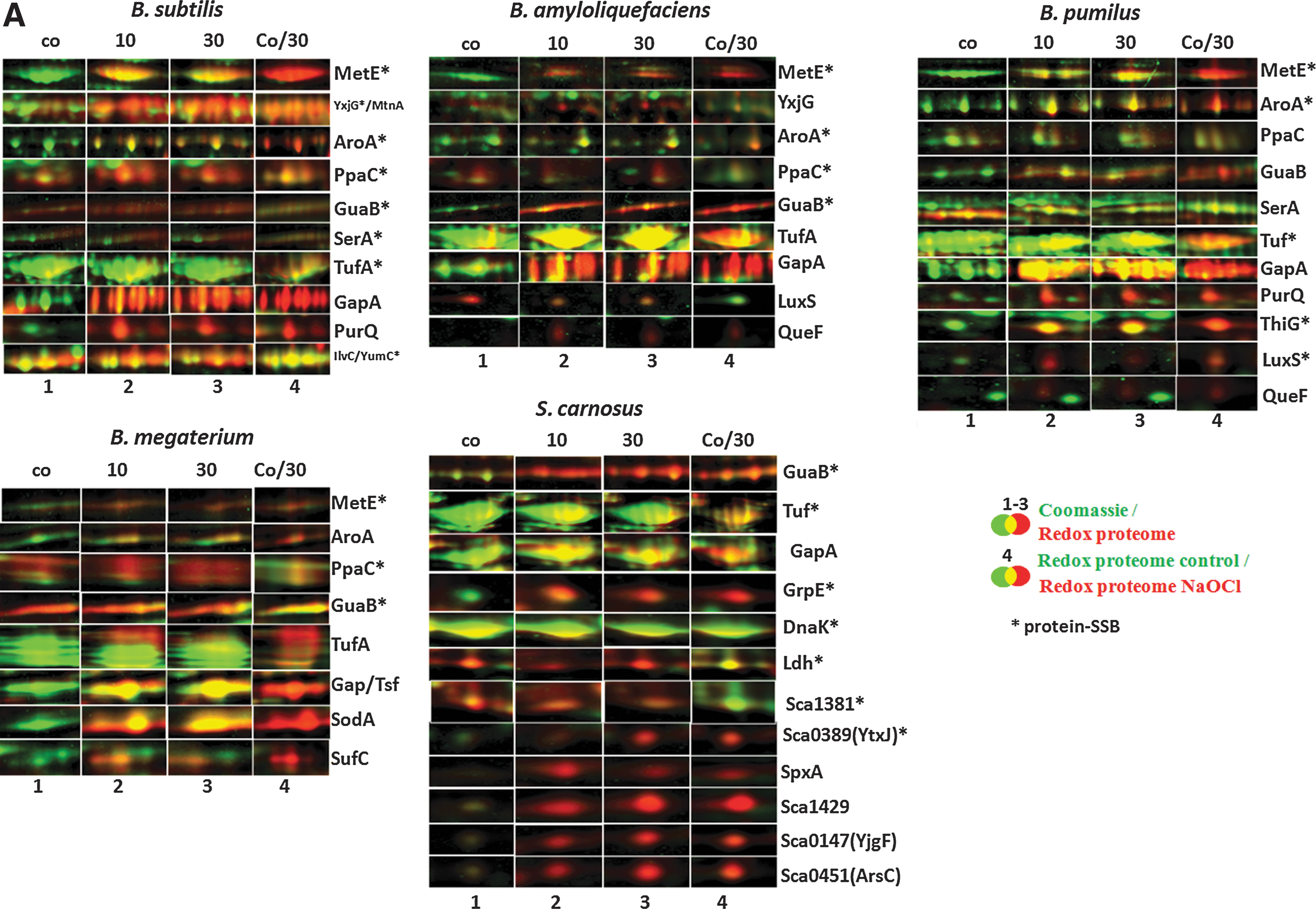
Previously, we have used redox proteomics based on the reduction of disulfides and labeling of released thiols with the fluorescence dye BODIPY FL C1-IA to identify proteins oxidized by NaOCl stress in B. subtilis (7). We applied our redox proteome approach for comparison of B. subtilis, B. amyloliquefaciens, B. megaterium, B. pumilus, and S. carnosus to identify proteins with disulfides after NaOCl stress, including those with potential SSB sites. The BODIPY-fluorescence image was overlayed with the Coomassie-stained protein amount image, and the oxidation ratios were calculated versus protein quantities (Fig. 2A, B and Supplementary Figs. S3–S7 and Table 1). The reversibly oxidized proteins were identified using MALDI-TOF/TOF mass spectrometry analysis (Supplementary Table S1), and the oxidation ratios were used for hierarchical cluster analyses (Supplementary Figs. S3–S7).

The proteins were cut from Coomassie-stained 2D-gels, tryptic in-gel-digested, and identified using MALDI-TOF-TOF MS/MS. The table lists the oxidation ratios/protein quantities of all identified reversibly oxidized proteins at control (co) and 10 and 30 min after NaOCl stress as average values of two biological replicate experiments (Exp1 and Exp2), their protein names and functions, Uniprot-accession numbers, and information about conserved Cys residues as revealed by the Conserved Domain Database CDD (33) (
BSH, bacillithiol; Cys, cysteine; MS/MS, tandem mass spectrometry.
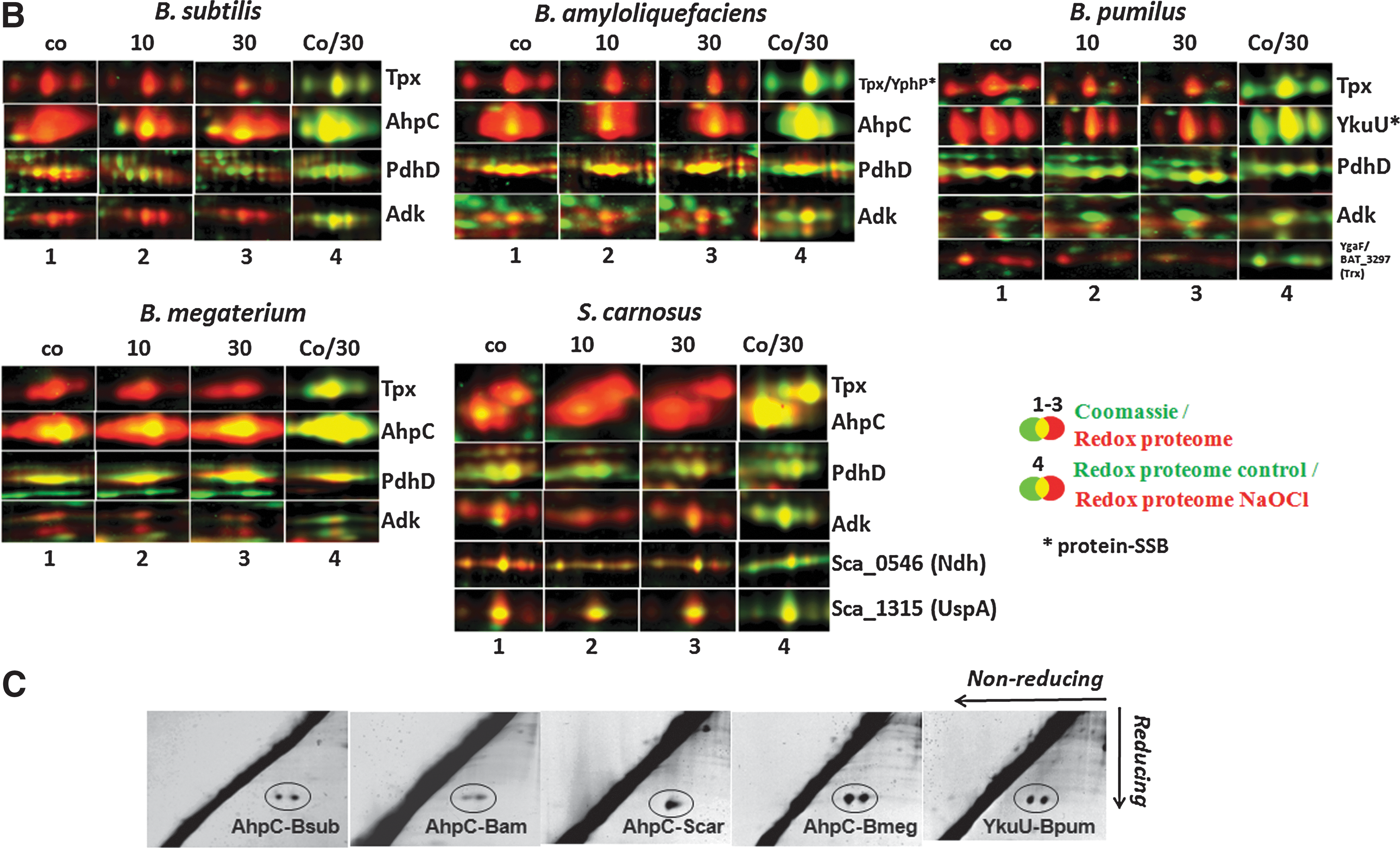
In total, 71 different proteins were identified in all five strains that are constitutively oxidized at either control or NaOCl stress conditions. Several conserved redox-sensitive proteins that coordinate metal ions (Zn, Fe-S-cluster) or form disulfide bonds for catalysis were identified in untreated cells, including the adenylate kinase Adk, the alkyl hydroperoxide reductase small-subunit AhpC, the 2-Cys peroxiredoxins YkuU and YgaF (in B. pumilus), the thiol peroxidase Tpx, the dihydrolipoamide dehydrogenase E3 subunit of the pyruvate dehydrogenase complex PdhD, and the sulfite reductase flavoprotein alpha-subunit CysJ (Fig. 2B). The intramolecular disulfide bonds were confirmed previously for PdhD, Adk, and CysJ in B. subtilis (7). In B. pumilus, which does not possess the AhpC-AhpF peroxiredoxin/reductase system (14), the YkuU 2-Cys peroxiredoxin is strongly oxidized similar to AhpC in other Bacillus strains, suggesting that YkuU might replace AhpC in peroxide detoxification. The intermolecular disulfides of AhpC homologs and YkuU were confirmed for all Bacillus strains and S. carnosus using diagonal nonreducing/reducing SDS–polyacrylamide gel electrophoresis (PAGE) analysis (Fig. 2C). Notably, in S. carnosus, the NADH dehydrogenase Sca_0546 and the universal stress protein UspA are among the most abundant proteins with strong oxidation ratios under control conditions (Supplementary Fig. S7).
Exposure of Bacillus strains and S. carnosus to sublethal NaOCl concentrations caused increased oxidation ratios of 57 different NaOCl-sensitive proteins (Table 1). The information about the conservations of Cys residues was obtained from the CDD database (
Cytoplasmic protein extracts of B. subtilis, B. amyloliquefaciens, B. pumilus, B. megaterium, and S. carnosus were prepared in an urea-IAM-buffer, tryptic in-gel-digested, and analyzed in a LTQ Orbitrap-Velos™ mass spectrometer as described in the Materials and Methods section. Peptides with S-bacillithiolations were identified by the additional mass of 396 Da at Cys residues and the diagnostic neutral malate loss fragment ions that appeared as abundant ions in the fragment ion MS/MS spectra. Peptides with S-cysteinylation were identified with a mass difference of 119 Da at Cys residues. The table includes the Uniprot accession numbers and protein functions as derived from the UniprotKB database (
IAM, iodoacetamide; LTQ, linear-trap quadrupole; MW, molecular weight.
We were interested to identify proteins with NaOCl-sensitive thiols that form SSB sites in Bacillus spp. and S. carnosus. Tryptic digests of protein extracts from control and NaOCl-treated cells were analyzed for S-bacillithiolations using Orbitrap-Velos LC-MS/MS analysis. The CID MS/MS spectra of the BSH-modified peptides show characteristic and abundant malate loss precursor ions (precursor-134 Da) that were used as diagnostic fragment ions to search for BSH-modified peptides and labeled in the MS/MS spectra (Table 2, Supplementary Tables S2 and Supplementary Figs. S8 and S9) (7). In our previous study, we found S-bacillithiolation sites for the thioredoxin-like protein YphP in B. subtilis under control conditions (7), but no further SSB sites were identified in Bacillus species and S. carnosus. In response to NaOCl stress, we could identify in total 54 proteins with characteristic SSB sites, including 29 unique proteins and eight conserved proteins with identical SSB sites in two or more Bacillus strains and S. carnosus. The eight proteins with conserved SSB sites include the methionine synthase (MetE), the inorganic pyrophosphatase (PpaC), the 3-D-phosphoglycerate dehydrogenase (SerA), the bifunctional 3-deoxy-7-phosphoheptulonate synthase/chorismate mutase (AroA), the translation elongation factor EF-Tu (TufA), the inosine 5′-monophosphate (IMP) dehydrogenase (GuaB), the thioredoxin-like protein (YphP), and the ferredoxin–NADP+ oxidoreductase (YumC) (Fig. 2A, E and Supplementary Figs. S8 and S9).
The methionine synthase MetE was oxidized most strongly and specifically by NaOCl stress in the redox proteome of all Bacillus strains (Fig. 2A and Supplementary Figs. S3–S7), and the conserved Cys730-SSB and Cys719-SSB sites were mapped by LC-MS/MS analyses (Supplementary Fig. S8). The YxjG methionine synthase paralog was only oxidized by NaOCl stress in B. subtilis and B. amyloliquefaciens (Supplementary Figs. S3 and S4). Further, NaOCl-sensitive enzymes with SSB sites at nonconserved Cys369 and Cys41, respectively, include the cystathionine-gamma synthase (MetI) and S-ribosylhomocysteine lyase (LuxS) of B. pumilus involved in homocysteine biosynthesis (Supplementary Figs. S8 and S9). The conserved proteins SerA with Cys410-SSB and PpaC with Cys158-SSB modifications (7) are constitutively oxidized in the redox proteomes of all strains, suggesting that basal S-thiolation is observed for selected proteins during aerobic growth (Fig. 2A and Supplementary Figs. S3–S7). The AroA protein was strongly oxidized in all redox proteomes by NaOCl, and the conserved Cys126-SSB site was identified by LC-MS/MS analysis (Fig. 2A and Supplementary Figs. S3–S7 and Table 2). AroA functions in the first step of chorismate synthesis catalyzing the production of 3-deoxy-
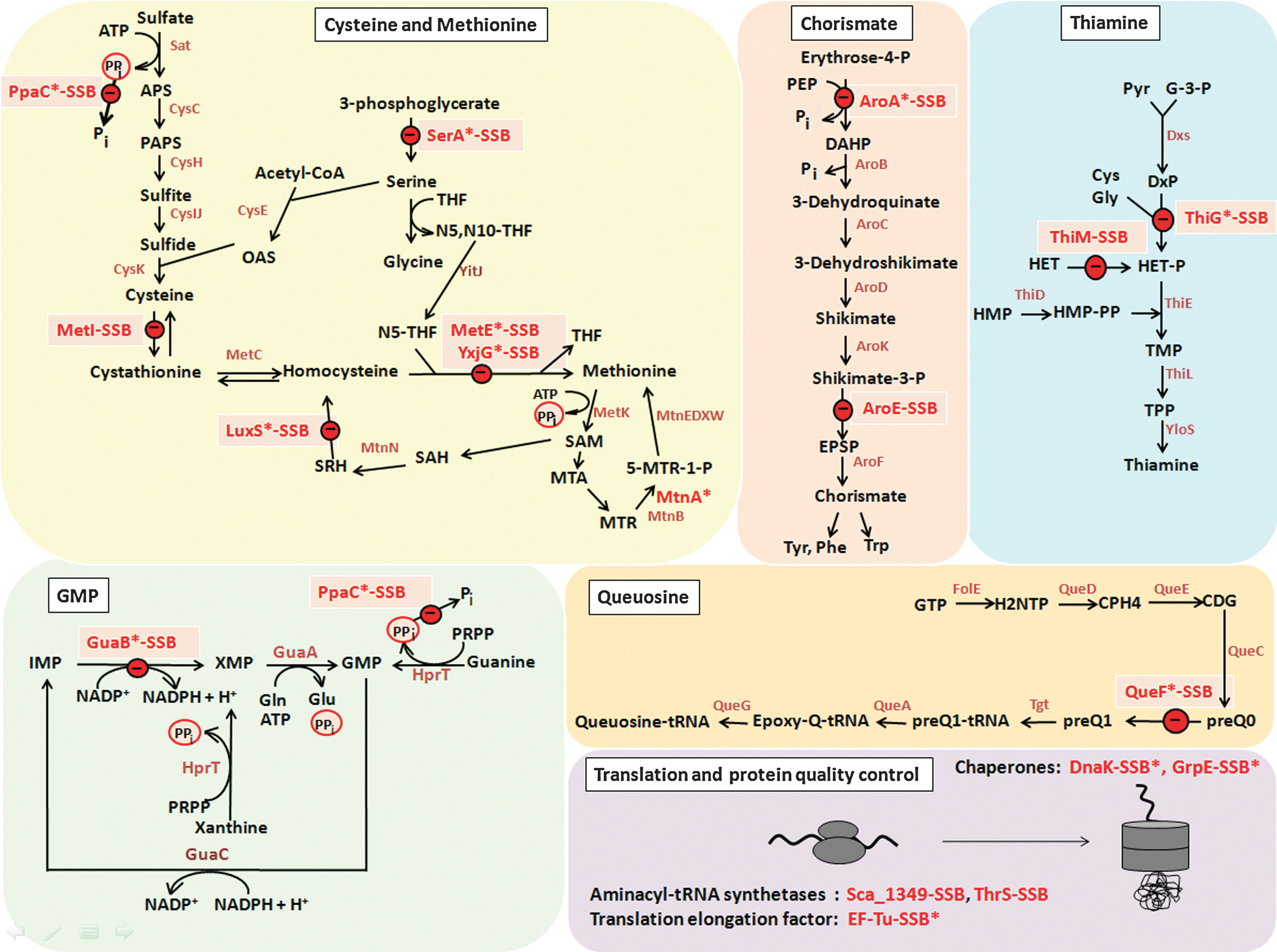
Among the redox and antioxidant proteins, the thioredoxin-like proteins YphP and YtxJ (Sca_0389) were identified with SSB sites at their active-site Cys53 and Cys30, respectively, and oxidized in the redox proteome of S. carnosus by NaOCl stress (Fig. 2A and Supplementary Figs. S8 and S9). The ferredoxin–NADP+ oxidoreductase YumC (23) was identified with an SSB modification at the active-site Cys85.
In S. carnosus, the lactate dehydrogenase Ldh and the putative transaldolase Sca_1381 are constitutively oxidized in the redox proteome, and the Cys72-SSB and Cys200-SSB sites, respectively, were identified by LC-MS/MS (Fig. 2A; Table 2). The aldehyde dehydrogenase AldA and the arginine deiminase ArcA are NaOCl-sensitive proteins in the redox proteome of S. carnosus that also possess active-site Cys residues.
NaOCl stress caused a decreased BSH/BSSB redox ratio in B. subtilis as shown by thiol metabolomics
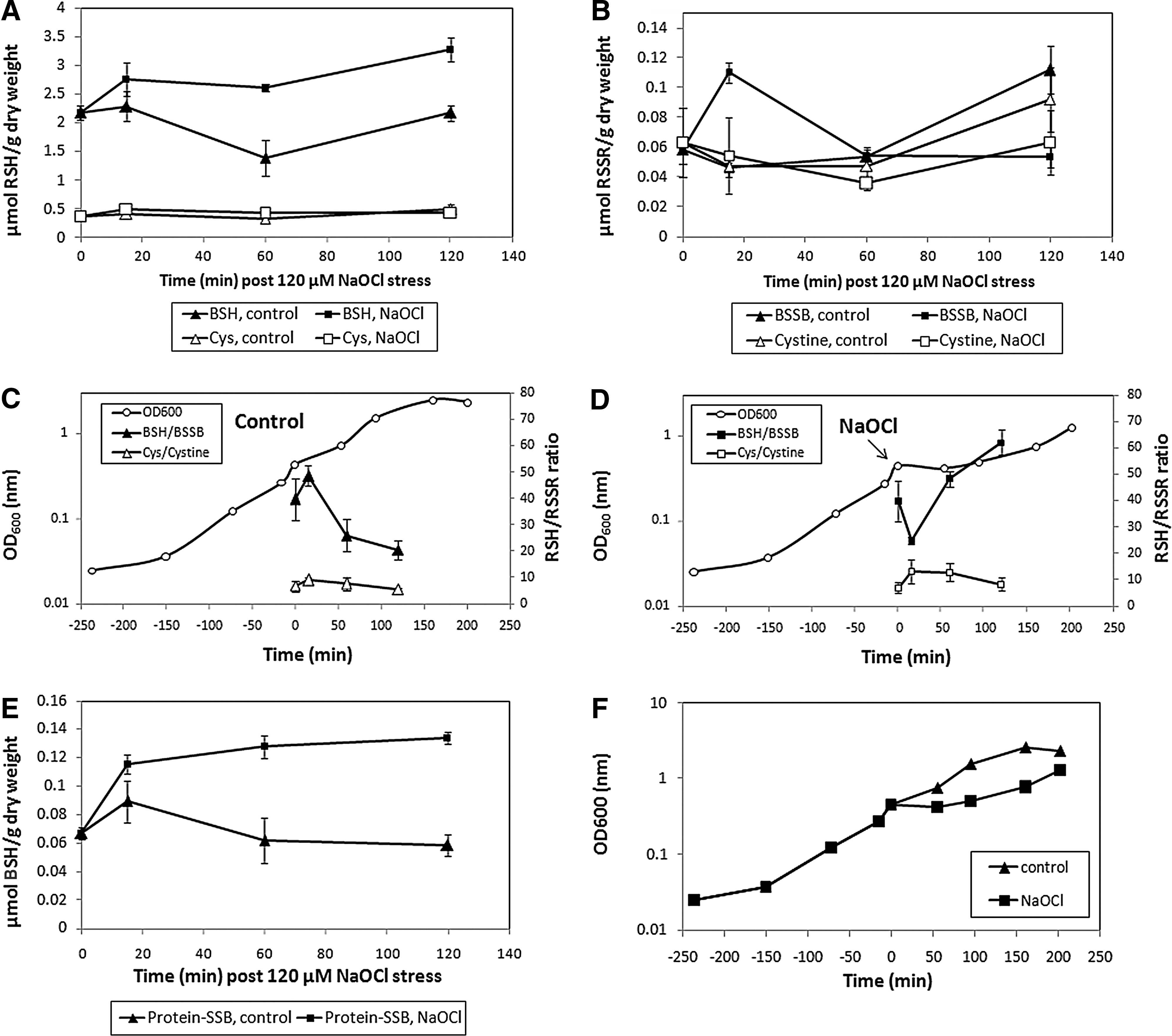
In B. subtilis, grown in an LB medium, similar amounts of BSH (0.6–1 μmol/g dry weight [DW]) and Cys (0.6 μmol/g DW) have been quantified previously (12, 40). The redox ratios were calculated previously as ∼400:1 for BSH/BSSB and ∼120:1 for Cys/Cystine in LB-grown B. subtilis cells (40). We quantified the change of the BSH/BSSB and Cys/Cystine redox ratios in B. subtilis cells grown in the BMM after NaOCl stress (Fig. 4 and Supplementary Fig. S10). The BSH level was 2.2 μmol/g DW in growing cells (OD500=0.4) and increased to 2.7 μmol/g DW directly after NaOCl stress and to 3.2 μmol/g DW during postexponential growth (Fig. 4A). BSH synthesis is about two-fold increased by NaOCl stress compared to control cells. The BSSB level was 0.06 μmol/g DW in growing cells, and NaOCl exposure resulted in a rapid two-fold increase to 0.11 μmol/g DW after 15 min (Fig. 4B). The BSSB level decreased within 60 min after NaOCl stress to control levels. The BSH/BSSB redox ratio was calculated as 40–50:1 during exponential growth, dropped two-fold shortly after NaOCl exposure, and recovered to 50–60:1 within 2 h (Fig. 4C, D). Approximately 3% (0.06 μmol/g DW) of the total cellular BSH content was bound as protein-SSB during growth and increased two-fold after exposure to 120 μM NaOCl (Fig. 4E). The BSH level is four- to eight-fold higher than Cys, and the levels of Cys (0.4 μmol/g DW), Cystine (0.05 μmol/g DW), and the Cys/Cystine redox ratio (∼10:1) were not affected by NaOCl stress. These data confirm that BSH is used as major redox buffer and oxidized to BSSB by NaOCl stress, causing a decreased BSH/BSSB redox ratio and S-bacillithiolation in B. subtilis cells.
Discussion
Previously, we have shown that BSH forms mixed protein disulfides (S-bacillithiolations) with essential catalytic Cys residues of enzymes involved in the methionine biosynthesis pathway and with the redox-sensing OhrR repressor after NaOCl stress in B. subtilis (7). Thus, S-bacillithiolation acts as a redox-switch under hypochlorite stress to induce methionine starvation and up-regulation of the OhrA peroxiredoxin for detoxification. In this work, S-bacillithiolation sites in 54 proteins have been identified among four different industrially important Bacillus species and S. carnosus after exposure to NaOCl. We applied our previous LC-MS/MS analysis to directly identify S-bacillithiolation sites and proved the MS/MS spectra by their characteristic malate loss precursor ions. The number of S-bacillithiolated proteins is lower than that of glutathionylated proteins identified in eukaryotes. This discrepancy could be due to the labile nature of the Cys-GlcN-Mal moiety that limits its detection, and also because the amount of BSH is much lower (2–3 μmol/g DW) compared to GSH in E. coli (19 μmol/g DW) or mycothiol (MSH) in Mycobacterium smegmatis (40 μmol/g DW) (40). Thus, selective thiol-trapping and enrichment tools need to be developed to quantify S-bacillithiolated proteins more comprehensively. To identify S-glutathionylated proteins in the proteome of eukaryotic organisms, redox proteomic approaches have been developed that use glutaredoxin (Grx) for specific de-glutathionylation of cell extracts, followed by biotin-N-ethylmaleimide (NEM) alkylation and affinity purification (e.g., the biotin-switch assay) (21, 28, 30, 31). By analogy, large-scale identification and quantification of BSH-mixed protein disulfides would require selective reduction by bacilliredoxins (Brxs), followed by alkylation and purification protocols. We identified possible candidates for putative Brxs (YphP and YtxJ) that were S-bacillithiolated at their active-site Cys residues. Current attempts are directed to develop Brx-based redox proteomics methods to further characterize and quantify the S-bacillithiolome, and to analyze the substrate specificities of these putative Brxs.
The fluorescence-based redox proteomics approach was used to quantify all oxidized proteins, and many conserved S-bacillithiolated proteins show increased oxidation ratios upon NaOCl exposure (e.g., MetE, YxjG, AroA, and ThiG). However, some conserved S-bacillithiolated proteins like TufA, GuaB, PpaC, and SerA did not show increased oxidation ratios by NaOCl stress in the redox proteomes of most Bacillus spp., indicating that S-bacillithiolation also occurs under nonstress aerobic growth conditions. This confirms results of S-glutathionylations in malaria parasites, human, and yeast cells, where the basal level S-thiolation occurs even under nonstress conditions as a result of aerobic growth (21, 24, 30). In Plasmodium falciparum, more than 400 S-glutathionylated proteins were identified in nonstressed cells (21). The authors further demonstrated in vitro that the activities of only a few identified metabolic enzymes were inhibited by S-glutathionylation. The physiological role and verification of the predicted basal level S-bacillithiolation of essential proteins in our study require further detailed investigations, since we could not identify S-bacillithiolations of these proteins in untreated cells using LC-MS/MS analysis. Overall, our combined redox proteomics and LC-MS/MS analyses showed a similar composition of the S-bacillithiolome compared to the S-glutathionylomes described for eukaryotes in oxidatively stressed cells (8, 31). The S-bacillithiolome contains mainly biosynthetic enzymes for amino acids, cofactors, nucleotides, as well as translation factors, chaperones, and redox and antioxidant proteins. Most proteins are enzymes involved in amino acid biosynthesis pathways, such as methionine, Cys (MetE, YxjG, MetI, LuxS, and PpaC), serine (SerA), aromatic amino acids (AroA and AroE), branched chain amino acids (YwaA), GTP synthesis (GuaB), queousine biosynthesis (QueF), and thiamine biosynthesis (ThiG, ThiI, and ThiM). We further identified S-bacillithiolated proteins involved in translation, such as elongation factor EF-Tu and aminoacyl-tRNA-synthetases; the heat-specific chaperones (DnaK and GrpE); and redox and antioxidant proteins, including thioredoxin-like proteins (YphP and YtxJ), the ferredoxin-NADP+ oxidoreductase (YumC), and peroxiredoxins (YkuU), which could function in the de-bacillithiolation process or require BSH for their regeneration.
The identified S-bacillithiolated proteins include eight conserved proteins identified in B. subtilis (MetE, GuaB, PpaC, SerA, AroA, Tuf, YphP, and YumC), which could be confirmed as S-bacillithiolated proteins in other species (Fig. 2D, E). Most conserved S-bacillithiolated proteins are abundant proteins oxidized in the redox proteomes of Bacillus and Staphylococcus species (e.g., MetE, Tuf, GuaB, PpaC, SerA, and AroA) and have conserved SSB peptides (Fig. 2A and Table 2). In addition, we detected 29 unique proteins that were S-bacillithiolated only in one Bacillus species or S. carnosus. The most unique proteins with SSB sites were identified in S. carnosus (12 SSB sites) and B. pumilus (8 SSB sites). These proteins are less abundant in the gel-based proteome and were only detected by LC-MS/MS analysis. For example, ThiM and ThiG proteins were found as specific targets for S-bacillithiolation in S. carnosus and B. pumilus, respectively. Novel S-bacillithiolated proteins were mapped in S. carnosus as the putative aldehyde dehydrogenase (Sca_1625), the transaldolase (Sca_1381), and the lactate dehydrogenases (Ldh), as well as the SceB precursor. These proteins are not conserved in Bacillus species, but homologs are present in the pathogen S. aureus. Interestingly, the chaperones GrpE and DnaK were S-bacillithiolated in S. carnosus, but Cys residues are absent in the GrpE and DnaK homologs of Bacillus species. The detection of some unique SSB-peptides could be also due to better peptide ionization properties of the tryptic peptides of less-conserved proteins.
Using BSH-specific immunoblots, we could demonstrate that MetE is the most abundantly S-bacillithiolated protein in all Bacillus species. MetE is most strongly oxidized by NaOCl stress in the redox proteome of B. subtilis wild-type and bshA mutant cells and is S-cysteinylated in the bshA mutant (7). Similar MetE oxidation ratios were calculated after 30 and 60 min post NaOCl stress in the wild-type and bshA mutant cells, but reduction of MetE took significant longer in the bshA mutant (Supplementary Fig. S11). Similarly, AroA was S-bacillithiolated in the wild-type cells and displays similar oxidation ratios in the wild-type and bshA mutant cells within the first hour, but is more slowly reduced in the bshA mutant. This may suggest that S-cysteinylation can fully compensate for S-bacillithiolation in the absence of BSH, but the reduction of these mixed disulfides occurs with different efficiencies.
The physiological importance of MetE inhibition by S-thiolation has previously been demonstrated. Methionine auxotrophy is one common phenotype in bacteria resulting from S-thiolation of MetE in B. subtilis and E. coli in response to oxidative stress (7, 19). In all Bacillus species, the conserved, active-site Cys730 and the nonconserved Cys719 were identified as SSB sites for MetE. The observed S-bacillithiolation of LuxS and MetI could be required to inhibit homocysteine biosynthesis after NaOCl stress, since homocysteine is toxic and accumulates when Met biosynthesis is inhibited, due to MetE-SSB modification (Fig. 3).
Among the conserved S-bacillithiolated proteins, the chorismate synthase AroA was identified, which catalyzes the synthesis of DAHP. Notably, the DAHP synthase Aro4p of yeast cells was found as a target for the Grx2 and showed increased thiol redox ratios at Cys76 and Cys244 in the grx2 mutant (35). The essential IMP dehydrogenase GuaB and the pyrophosphatase PpaC have been previously found to be S-glutathionylated in human T-lymphocytes and endothelial cells under diamide stress conditions (11, 30).
An important finding of this study is the identification of S-bacillithiolated proteins involved in translation and protein quality control, such as elongation factor EF-Tu and the DnaK and GrpE chaperones. The heat-specific Hsp70 and Hsp90 chaperones are well represented in the S-glutathionylomes of human and yeast cells (11, 45). The elongation factor TufA promotes GTP-dependent binding of aminoacyl-tRNAs to the A-sites of the ribosome, but also has chaperone functions (4, 47, 49). The TufA protein was oxidized at Cys256 by NaOCl stress in E. coli, and a tufA mutant was highly sensitive to NaOCl (27). In the Bacillus spp. and S. carnosus TufA proteins, we identified the conserved Cys82-SSB modification in the GTP-binding site. Since TufA is essential in B. subtilis, its S-thiolation in oxidatively stressed cells could cause inhibition of protein synthesis during oxidative stress as previously shown in yeast cells (45). Thus, the fact that amino acid biosynthetic enzymes and translation proteins dominate in the conserved S-bacillithiolome suggests that S-bacillithiolation could control protein synthesis during oxidative stress. The strong repression of the stringent controlled RelA regulon, including genes for ribosomal proteins, by NaOCl stress in our previous microarray analysis further indicates a decreased protein synthesis that could be caused by S-bacillithiolation (7). The activation of the stringent response by ROS and RNS has also been shown in Salmonella (2, 17). Specifically, the stringent response regulatory DnaK suppressor protein (DksA) was shown to be play a role in controlling GSH biosynthesis and other metabolic pathways associated with the generation of reducing power under oxidative stress conditions (17).
The next question is how the reversibility of this redox mechanism (protein de-bacillithiolation) might be mediated. Redoxins catalyzing the reversible S-thiolation/reduction of S-glutathionylated proteins include Grxs, thioredoxins, glutathione S-transferases, and GSH disulfide reductases that were identified in large-scale redox proteomics studies (21). The mechanisms of protein de-glutathionylation by mono- and dithiol Grx have been studied in plants and mammalian systems (13, 52). In both mechanisms, the nucleophilic active-site Cys first attacks the GSH-mixed protein disulfide, resulting in substrate de-glutathionylation and formation of a Grx-SSG intermediate (52). In plant dithiol Grx, this Grx-SSB intermediate is then attacked by a second reactive Cys not present in the CXXC motif that leads to Grx intramolecular disulfide formation. This Grx disulfide is finally resolved by an NADPH-dependent ferredoxin:thioredoxin reductase. In the monothiol mechanism, the Grx-SSG intermediate is regenerated by GSH that is oxidized to GSSG and requires the GSH reductase to restore the reduced GSH pool (52). The thioredoxin-like proteins YphP and YtxJ could possibly function as putative Brxs. YphP is a DUF1094 family protein whose previously solved crystal structure adopts a thioredoxin-fold, has a conserved C53XC55 motif (with a reduction potential of −130 mV), and was demonstrated to display some weak disulfide isomerase activity (10). Under NaOCl stress, YphP is S-bacillithiolated at its more solvent-exposed active-site Cys53. The DUF2847 family Brx protein YtxJ is another thioredoxin-like protein with a conserved TCPIS motif reminiscent of the same redox-active motif found in many monothiol Grxs (18). YtxJ was identified in S. carnosus, S-bacillithiolated in its TCPIS motif. The detection of S-bacillithiolated YphP and YtxJ during NaOCl stress could represent the trapping of transient intermediates of Brx-mediated de-bacillithiolation pathways. The distribution of YphP and YtxJ homologs among bacteria encoding BSH biosynthesis genes (12) further suggests their roles as Brx candidates. Our current experiments are directed to further investigate the functions of these putative Brx proteins in the de-bacillithiolation pathway.
We further analyzed whether the BSH/BSSB redox ratio is affected by NaOCl stress that could contribute to S-bacillithiolation of proteins. In E. coli cells, a four-fold decrease in the GSH/GSSG redox ratio from 370:1 to 87:1 was observed in menadione-treated cells (46). The two-fold increase of BSSB post NaOCl stress is comparable to the increase of GSSG in E. coli cells exposed to menadione (46). The increased GSSG levels result in a decreased GSH/GSSG ratio that could contribute to protein S-glutathionylation spontaneously via thiol–disulfide exchange between GSSG and protein thiols (13). However, this thiol–disulfide exchange is kinetically too slow and very unlikely in vivo, since it would require a drop in the GSH:GSSG ratio from 100:1 to 1:1 for 50% conversion of protein thiols to protein GSH-mixed disulfides (13, 52). In our studies, the BSH:BSSB redox ratio dropped two-fold, confirming that BSH is oxidized to BSSB by NaOCl. This BSH:BSSB ratio of 25:1 is unlikely to be sufficient for spontaneous formation of protein-SSB via a direct thiol–disulfide exchange reaction between protein-SH and BSSB. Instead, S-bacillithiolation first requires oxidation of reactive Cys residues to activated thiol derivatives, such as sulfenic acids (9, 15). NaOCl leads to chlorination of Cys residues to unstable sulfenylchloride intermediates that rapidly react further to form disulfides (50). Thus, we suggest that sulfenylchloride formation at reactive Cys residues by NaOCl and the decreased BSH/BSSB redox ratio contribute to the mechanism of S-bacillithiolation in Firmicutes bacteria.
Finally, the question arises if hypochlorite is a physiologically relevant oxidant that mediates protein S-bacillithiolation in Bacillus and Staphylococcus species. Hypochlorite is encountered by soil bacteria and pathogenic Bacillus and Staphylococcus species, since it is present in household bleach and other disinfectants, and it is also produced by activated macrophages during the infection process by the enzyme myeloperoxidase. The observed S-bacillithiolations in soil-dwelling Bacillus species after NaOCl stress might mimic the protective response to toxic ROS, such as hydroxyl radicals, which are generated during respiration, explaining the conservation of some of the protein targets across different species.
Materials and Methods
Bacterial strains and growth conditions
The bacterial strains used were B. subtilis wild-type strains 168 (trpC2) and CU1065 (trpC2 pheA1) and bshA-mutant strain HB11002 (CU1065 trpC2 and bshA::mlsr) (12), B. amyloliquefaciens FZB42 (5), B. pumilus SBUG1799, B. megaterium SBUG1152 (36), and S. carnosus TM300 (43). Cultivation was performed as detailed in the Supplementary Materials and Methods section.
Thiol redox proteome analysis and protein identification using MALDI-TOF-TOF MS/MS
The thiol redox proteome analysis was performed as described (7). Cells were harvested at control and NaOCl stress conditions, sonicated, alkylated in 8 M urea/1% chaps/100 mM iodoacetamide (IAM), and acetone-precipitated. The pellet was resolved in an urea/chaps buffer without IAM, and 200 μg of the protein extract was reduced with 10 mM Tris-(2-carboxyethyl)-phosphine and labeled with BODIPY FL C1-IA [N-(4,4-difluoro-5,7-dimethyl-4-bora-3a,4a-diaza-s-indacene-3-yl)-methyl)-iodoacetamide] (Invitrogen, Eugene, OR). The fluorescence-labeled protein extract was separated using 2D PAGE, scanned for BODIPY fluorescence, and stained with Coomassie as described (7). Quantitative image analysis was performed with DECODON Delta 2D software (
Linear-trap quadrupole–Orbitrap Velos mass spectrometry
Bacterial cells were sonicated in an urea/chaps alkylation buffer with 100 mM IAM and separated by 15% nonreducing SDS-PAGE. In-gel tryptic digests and Orbitrap Velos LC-MS/MS analysis were performed as described in the Supplementary Materials and Methods section. S-bacillithiolated peptides were identified by searching all MS/MS spectra in .dta format against B. subtilis 168, B. megaterium, B. amyloliquefaciens FZB42, B. pumilus, and S. carnosus TM300 target-decoy protein sequence databases extracted from UniprotKB release 12.7 using Sorcerer™-SEQUEST® according to the parameters and SEQUEST filter criteria as described in the Supplementary Materials and Methods section.
Preparation of keyhole limpet hemocyanin-BSH for generation of BSH-antibodies
Two milligrams of synthetic BSH (44) was conjugated to 10 mg keyhole limpet hemocyanin (KLH) using the Maleimide-Activated KLH Conjugation Kit (Sigma) according to the details described in the Supplementary Materials and Methods section.
Nonreducing BSH-immunoblot analysis and nonreducing/reducing diagonal SDS-PAGE
Polyclonal BSH antisera were used at 1:500 dilution, and 25 μg IAM-alkylated protein extracts was used for nonreducing 15% SDS-PAGE and immunoblot analysis as described (6). Nonreducing/reducing diagonal SDS-PAGE analysis was performed as previously described (6).
Quantification of reduced thiol redox buffer (RSH), oxidized thiol redox buffer (RSSR), and protein BSH
B. subtilis CU1065 was grown in BMM to an OD600 of 0.44 and harvested before and after NaOCl stress for subsequent RSH, RSSR, and protein RSH quantification as described in the Supplementary Materials and Methods section.
Footnotes
Acknowledgments
We thank the Decodon Company for support with Decodon Delta 2D software, Frieder Schauer for providing strains B. pumilus SBUG1799 and B. megaterium SBUG1152, and Dana Clausen for excellent technical assistance. We are very grateful to Ahmed Gaballa and John D. Helmann for discussion of the results before publication and for providing the B. subtilis bshA mutant. This work was supported by grants from the Deutsche Forschungsgemeinschaft (AN746/2-1 and AN746/3-1) to H.A., by project TRIG A from the University of Science, Hanoi National University to T.T.T.H., and by a grant from the Biotechnology and Biological Science Research Council (BB/H013504/1) to C.J.H.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.