
Abstract
Introduction
Biomarker is a characteristic that is objectively measured and evaluated as an indicator of normal biological processes, pathogenic processes, or responses to a therapeutic intervention. A few biomarkers of oxidative stress/damage are validated (16, 18, 23, 43). Proteins are major targets of oxidants and can undergo reversible redox reactions, which can also be part of their normal function, or irreversible oxidative damage at various amino acids, which are generally associated with a (permanent) loss of function (14 –18, 23). Whenever ascertained, oxidative modifications may allow to establish a putative relationship between structural/functional protein alterations and pathological conditions.
Research toward a better understanding of the molecular mechanisms of a disease together with identification of specific targets of oxidative stress/damage is urgently necessary. Although tissue samples represent the best choice for understanding direct target-organ effects, there is an overwhelming, impelling need to validate more specific oxidative damage biomarkers that can be easily measured in human biological fluids obtained from noninvasive approaches, to overcome limitations of invasive monitoring (7, 43). The most accessible biological fluids for potential biomarker discovery are intravascular fluids (blood and plasma) and extravascular fluids, such as cerebrospinal fluid (CSF), synovial fluid (SF), and bronchoalveolar lavage fluid (BALF). In particular, blood plasma is a rich source of information, because it contains a large majority of all human proteins, including albumin. Since alterations in some blood parameters can reflect alterations even in hardly achievable compartments of the body, the redox state of albumin has been analyzed in some human diseases related to oxidative stress.
Metabolism and Distribution of Human Albumin
In healthy adults, albumin synthesis occurs almost exclusively in hepatocytes (∼0.2 g/kg body weight per day) and accounts for 10% of total liver protein synthesis. Whether human serum albumin (HSA) found in milk is produced by the mammary gland or derived from the circulation has not yet been resolved (19, 56). Recently, convincing evidence of albumin synthesis in human microglial cells has been provided (19). There is no storage of newly synthesized albumin, only a small amount (<2%) in transit during its 30-min trip through hepatocytes. Under physiological conditions, only 20%–30% of hepatocytes produce HSA and its synthesis can therefore be increased on demand by a factor of 200%–300% (56). The colloid osmotic pressure and the osmolality of the interstitial liquid around the hepatocytes is the most important regulator of HSA biosynthesis, which is also regulated by the nutritional status and hormones, including insulin, glucagon, cortisol, and thyroid hormones (19, 56). Under physiological conditions, a steady state exists between albumin synthesis and metabolism. The amount of albumin metabolized daily is proportional to the plasma concentration: a fixed percentage of ∼10% of HSA content is metabolized per day. The catabolism of HSA takes place in several organs, primarily in muscle, skin, and the liver, after ∼20 days from its synthesis at a rate of about 14 g per day in a 70 kg healthy adult, or 4% of whole-body protein turnover (19, 56).
Albumin distribution is adequately described by a two-compartment model where about 40% is taken up by the intravascular and 60% by the extravascular space (Fig. 1). Intravascular (blood plasma) HSA is the most abundant protein (mean concentration ∼43 mg/ml, range 35–52 mg/ml, i.e., ∼0.6 mM), accounting for 50%–60% of total plasma proteins in healthy humans (56). Measurement of albumin concentration in the extravascular fluids has been performed in healthy humans (Supplementary Table S1; Supplementary Data are available online at
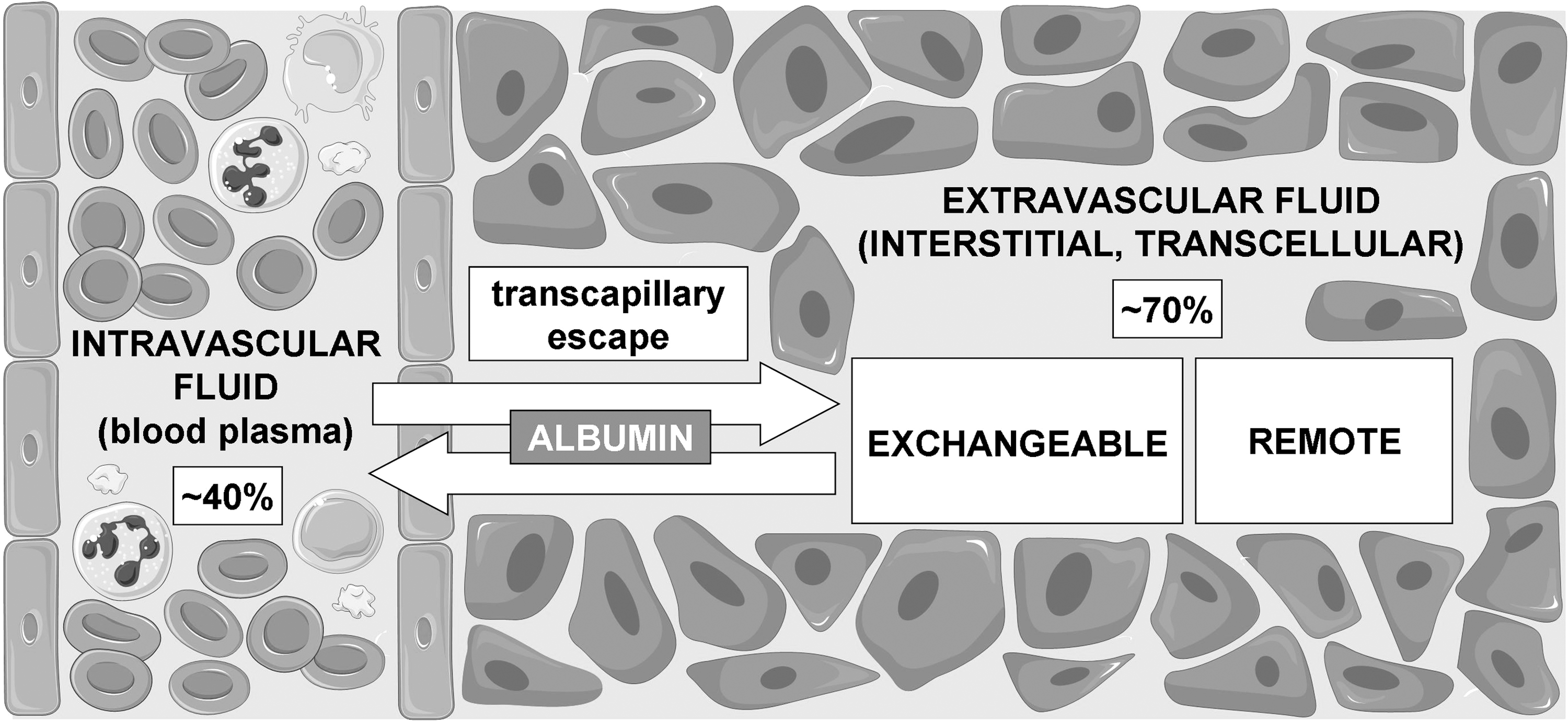
Structure and Functions of Human Albumin
HSA is a single polypeptide of 585 amino acids (66.5-kDa), including 35 cysteine residues forming 17 intramolecular disulfide bonds, with the only free sulfhydryl group located at Cys34 (56). Reduced form of HSA (Alb-SH) is also known as human mercaptalbumin (HMA, with Cys34 containing a free thiol). Within the tertiary protein structure of HSA, Cys34 resides in a unique microenvironment close to three ionizable residues, Asp38, His39, and Tyr84 (69), which affect the ionization state of Cys34, thus modulating its reactivity (19, 55). As a result, Cys34 has an unusually low pK a (<6.7 vs. ∼8.0–8.5 for thiols in most other proteins) and exists primarily in the highly nucleophilic thiolate form (72). However, HSA presents pH-dependent structural transitions that impact on thiol reactivity, and consequently, the Cys34 pK a value remains controversial (77). Moreover, the reactivity of the Cys34 thiol is affected by the presence of ligands. For example, fatty acids, which bind to sites relatively distant from Cys34, induce conformational changes at the thiol site and increase the thiol reactivity toward disulfides (77). Indeed, the three-dimensional (3D) structure of lipidated albumin shows increased exposure of Cys34 compared to fatty acid-free albumin (19). The enhanced reactivity of the albumin Cys34 thiol and its sulfenic derivative at neutral pH upon fatty acid binding can be explained by considering that the corresponding conformational changes that increase thiol exposure increase both the availability of the thiolate due to a lower apparent pK a and also loosen steric constraints for reactions (77). Considering that situations that tend to increase the amount of fatty acids bound to HSA (such as diabetes mellitus) are associated with oxidative stress, a better antioxidant ability of HSA could constitute a protective adaptation. Indeed, oxidized forms of HSA are found increased under these situations (Table 1). The high reactivity of HSA toward electrophilic reactive carbonyl species is due to several accessible nucleophilic residues, mainly Cys34, followed by His146 and Lys199 (3).
280/340 nm.
Hemodialysis patients with diabetes mellitus.
Hemodialysis patients without diabetes mellitus.
of the thiol-mBrB conjugate.
Alb-Cys, S-cysteinylated albumin (S-thiolated albumin, also called human nonmercaptalbumin1 [HNA-1]); Alb-SH, mercaptalbumin; Alb-SO2H, albumin sulfinic acid (also called human nonmercaptalbumin2 [HNA-2]); Alb-SO3H, albumin sulfonic acid (also called human nonmercaptalbumin2 [HNA-2]); Alb-SOH, albumin sulfenic acid; Alb-SSR, albumin-mixed disulfide (S-thiolated albumin, also called human nonmercaptalbumin1 [HNA-1]); LC-ESI-MS/MS, liquid chromatography–electrospray ionization tandem mass spectrometry; HPLC, high performance liquid chromatography.
Albumin can also bind metal ions such as Cu(II), Pt(II), Ag(I), Hg(II), and Co(II), in up to four sites with different specificities (56), which makes it susceptible to carbonylation via metal-catalyzed oxidation reactions.
The secondary structure of HSA is dominated by α-helices (68%), without any β-sheet. HSA is arranged in a globular heart-shaped conformation, organized in three homologous domains (I, II, and III), each consisting of two subdomains, A and B (12, 19, 73).
Albumin is responsible for about 80% of intravascular colloid oncotic pressure and predominantly contributes to the interstitial colloid osmotic pressure; it also modulates the capillary permeability, neutrophil adhesion, and activation (19, 52). HSA excellently binds and transports both endogenous and exogenous molecules, including long-chain fatty acids, cholesterol, bile pigments, hormones, proteins and peptides (e.g., bilirubin, ceruloplasmin, and apoliporoteins), nitric oxide, many drugs (e.g., ibuprofen and warfarin), ion metals (19, 20, 52), as well as reactive carbonyl species (3, 11).
Antioxidant activity has been recognized as a significant function of albumin in the plasma, a body compartment exposed to continuous oxidative stress where only a limited amount of antioxidant enzymes are available (59). The antioxidant activity of HSA results from the redox properties of the Cys34 residue and from its metal-binding properties. Due to the high plasma concentration of HSA, its Cys34 sulfhydryl group represents the largest fraction (∼80%, corresponding to ∼500 μM) of all free thiols in plasma, thus conferring a major role in the plasma antioxidant capacity to HSA (52, 63). In addition, as albumin is the predominant protein within BALF (which is representative of components in alveolar space), CSF, and SF (37, 48, 64), it may be the major source of free thiols in BALF, CSF, and SF as well. Therefore, Alb-SH may be an important endogenous antioxidant within the body.
HSA also displays an indirect antioxidant activity by binding several compounds, including oxysterols, bilirubin, and homocysteine (52, 63).
Thiol Oxidation in Human Albumin
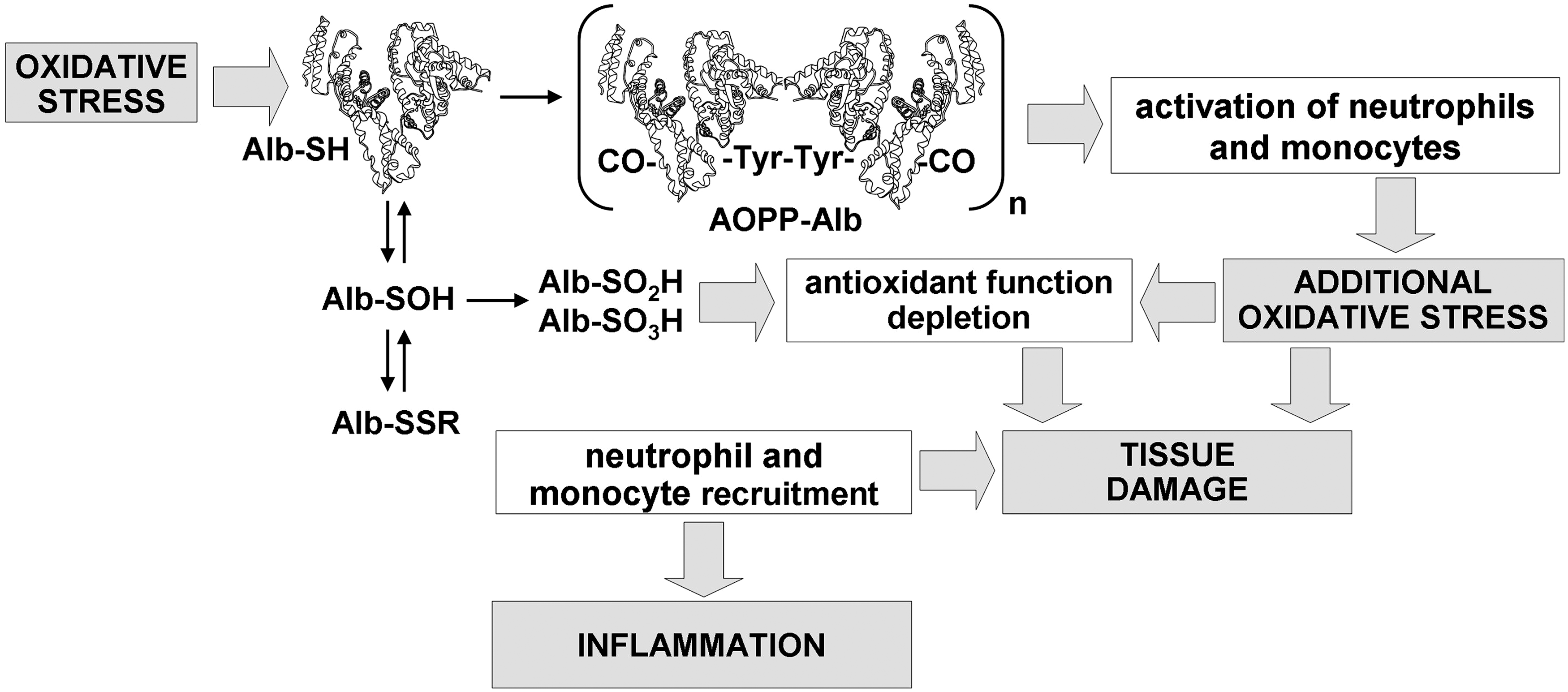
The albumin Cys34 thiol exists in both reduced and oxidized forms (Fig. 2). In plasma of healthy young adults, 70%–80% of total HSA contains the free sulfhydryl group of Cys34. About 20%–30% of the Cys34 forms a reversible mixed disulfide with low-molecular-weight thiols, like cysteine, homocysteine, cysteinylglycine, or glutathione, generating S-thiolated albumin (Alb-SSR, also called human nonmercaptalbumin1 [HNA-1]), the predominant modification being S-cysteinylated albumin (Alb-Cys) (5, 27, 31, 65). A small fraction of the Cys34 (2%–5%) is more highly oxidized to the sulfinic (Alb-SO2H) or sulfonic acid (Alb-SO3H) form (79, 80), which are also called human nonmercaptalbumin 2 (HNA-2) and can cause a loss of protein function. The sulfenic acid form (Alb-SOH) is a central intermediate in the formation of oxidized albumin species and increases under pathological conditions (10, 79, 80). Alb-SOH is often included within the HNA-2 fraction. The Cys34 thiol can also become S-nitrosated (78).
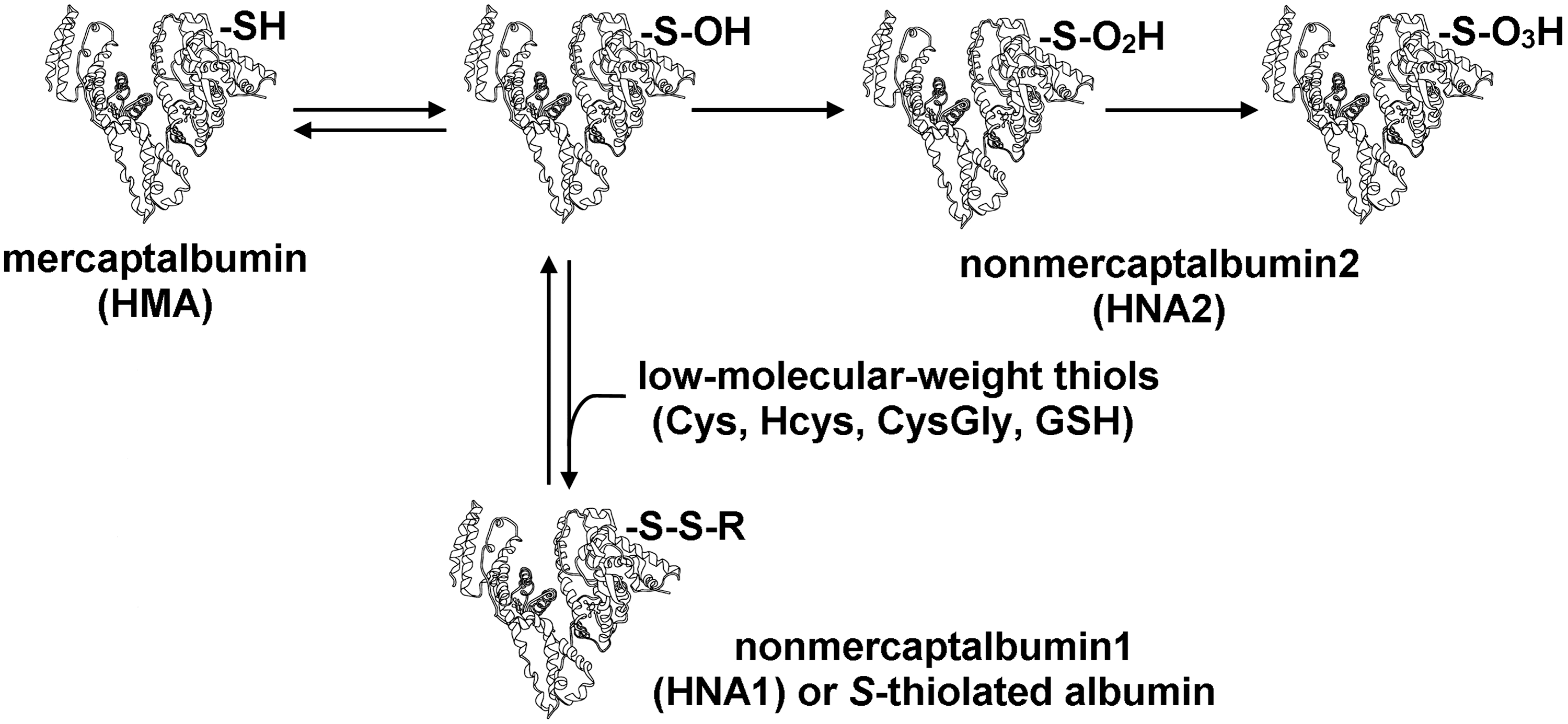
While in vitro studies have long supported the antioxidant role of HSA in plasma, evidence that albumin oxidation takes place also in vivo has been reported only recently. In particular, the advent of potent mass spectrometric techniques applied to albumin, together with chromatographic analytical approaches, has made detection of albumin redox modifications in different pathophysiological conditions possible, where decreases in HMA correlate with increases in oxidized HSA forms (mixed disulfides, HNA-1, and higher oxidation states, HNA-2) (Table 1). These oxidized species are not present when albumin is secreted from hepatocytes, and thus they may constitute potential biomarkers of the involvement of the oxidative stress/damage in human disease and of the antioxidant activity of the albumin Cys34.
Studies on plasma HSA performed by means of liquid chromatography–electrospray ionization tandem mass spectrometry (LC-ESI-MS/MS) demonstrated for the first time in vivo massive and stable oxidation of Alb-SH into Alb-SO3H in nephrotic syndrome and focal segmental glomerulosclerosis (8, 44, 45), a renal disease characterized by sclerosis of the glomerular tuft, for which an increased oxidative stress has been hypothesized. This oxidation produces relevant changes to the protein structure and is thought to play a role in the pathogenesis of the proteinuria associated with focal segmental glomerulosclerosis (44, 45). Thiol oxidation to -SO3H increases the net negative charge of albumin as demonstrated by using the electrophoretic titration curve, a 2D electrophoretic technique where proteins move perpendicularly to a pH gradient generated by a stack of carrier ampholytes, generating sigmoidal pH/mobility curves proportional to their chemical ionization (8, 44).
Oxidative modifications of HSA Cys34 have been reported in advanced liver disease. Marked increases in the percentages of HNA-1 and/or HNA-2, and a corresponding decrease in the Alb-SH percentage, were measured by high performance liquid chromatography (HPLC) with UV detection in both acute-on-chronic liver failure (50, 51) and liver cirrhosis (21, 50, 51, 81), these differences being even more pronounced in the former (50, 51). The redox state of HSA was also used as a marker to investigate the current status of oxidative stress in patients with chronic renal failure. The fraction of oxidized HSA forms (HNA-1 and HNA-2) is elevated in correlation with the degree of renal dysfunction, even before dialysis (75, 76). In addition, albumin oxidation is a powerful independent risk factor for a serious cardiovascular disease incidence and mortality among patients on hemodialysis (75). These reports suggest that the redox state of HSA is a good marker to investigate the current status of oxidative stress in patients with chronic renal failure, which is enhanced in correlation with the level of renal dysfunction.
S-thiolated albumin increases during aging and in different kinds of diseases characterized by oxidative stress, as measured by HPLC with fluorescence detection (Table 1). The percentage of Alb-SSR significantly increases with age in healthy people (22, 49, 65), in particular that of Alb-Cys (22, 65). This indicates that the antioxidant efficiency of the protein thiol pool in plasma decreases with age, causing an increased risk of irreversible oxidation of protein sulfhydryl groups and, therefore, a depletion and/or impairment of the antioxidant capacity of plasma.
Plasma Alb-SSR is increased in patients with mild cognitive impairment and Alzheimer's disease (24). Although plasma Alb-SSR is not a specific marker for neurodegenerative disease, this biomarker of oxidative stress (together with carbonylated proteins and malondialdehyde) was proposed to be a more useful marker for neurodegenerative disorders than homocysteine, vitamin B12, or folate (24). Increased levels of Alb-SSR as a systemic marker of oxidative stress have also been measured in the plasma of hemodialysis patients (38, 70), of patients with diabetes mellitus and, to an higher extent, of those with proliferating diabetic retinopathy, which also showed marked increases in the irreversibly oxidized HSA forms (53). In addition, levels of Alb-SSR and HNA-2 were found markedly increased in the aqueous humor of patients with senile cataract with no complications (30). However, further study is required to elucidate the eventual relationship between the redox state of the aqueous humor albumin and cataract formation.
Elevated levels of Alb-Cys were detected by HPLC coupled with ESI-time-of-flight (TOF)-MS in maternal plasma of pregnancies with intrauterine growth restriction (4) and by HPLC coupled with nanoESI-quadrupole-TOF-MS in amniotic fluid (where albumin is the most abundant protein) of pregnancies associated with gestational diabetes mellitus (6).
Albumin Carbonylation
Many studies have shown the presence of elevated levels of carbonylated albumin (Alb-CO) in various human diseases and pathophysiological conditions by means of spectrophotometric and immunochemical methods (enzyme-linked immunosorbent assay [ELISA], or sodium dodecyl sulfate–polyacrylamide gel electrophoresis [SDS-PAGE] combined with Western blot (OxyBlot) as well as redox proteomics (2D polyacrylamide gel electrophoresis [2-DE], combined with Western blot [2D-OxyBlot], eventually coupled to the MS analysis) (Table 2). Spectrophotometric and most immunochemical assays rely on the use of 2,4-dinitrophenylhydrazine (DNPH) to derivatize protein carbonyl groups, which forms a stable 2,4-dinitrophenylhydrazone (DNP) adduct that is then detected, respectively, by either absorbance at 366–370 nm or by using an anti-DNP antibody (17).
Standard agarose gel electrophoresis, after which proteins in the gel were submitted to reaction with DNPH and resolved in the second stage by SDS-PAGE.
BALF, bronchoalveolar lavage fluid; MALDI-TOF, matrix-assisted laser desorption/ionization time of flight; DNPH, 2,4-dinitrophenylhydrazine; ELISA, enzyme-linked immunosorbent assay; SDS-PAGE, sodium dodecyl sulfate–polyacrylamide gel electrophoresis.
Albumin is the main plasma carbonylated protein in chronic kidney disease and hemodialysis patients, as evidenced by using for the first time a Western blotting technique to detect differential susceptibility of plasma proteins to carbonyl formation in vivo in human disease (26). Carbonylation, also associated to dityrosine formation (34), alters the structural and functional properties of uremic HSA (34, 38), suggesting that oxidation of plasma HSA might adversely affect its vasculoprotective effects in hemodialysis patients. A further study also demonstrated increased albumin modification by advanced glycation end products (AGEs), and that the amount of both Alb-CO and AGE-modified HSA was the highest in hemodialysis patients with diabetes mellitus (39), a pathology where increased levels of serum Alb-CO were further confirmed (62). Increasing albumin carbonylation parallels the severity of chronic kidney disease and is even higher in the plasma of hemodialysis patients, as detected by the OxyBlot assay, indicating that oxidative stress in such patients gradually increases during the development of the disease (40). Alb-CO was also detected by the OxyBlot assay in urine of patients with chronic kidney disease (1) as well as in the plasma and urine in diabetic nephropathy (2).
There is evidence that oxidative damage occurs in rheumatoid diseases. OxyBlot showed that levels of serum Alb-CO were significantly higher in patients with rheumatoid arthritis than in controls (32). Furthermore, by means of two-stage electrophoresis, by which serum proteins were separated by standard agarose gel electrophoresis, submitted to reaction with DNPH, and resolved in the second stage by SDS-PAGE, followed by OxyBlot, albumin and γ-globulins were found to be the most carbonylated proteins in plasma of juvenile idiopathic arthritis (84).
Progressive increase of plasma Alb-CO levels, associated with increased oxidation of Cys34 (Table 1), was found by ELISA and OxyBlot assays, with increasing severity of liver cirrhosis and acute-on-chronic liver failure (51). The increase in total serum protein carbonyl levels in hepatocellular carcinoma patients (61) as well as in lung cancer patients (60) was largely due to an increase in Alb-CO levels.
In humans, a major cause of chronic oxidative stress is cigarette smoking. By the OxyBlot analysis, the major carbonylated protein in the BALF of older smokers with long-term smoking histories was found to be albumin, even in the absence of lung diseases (46, 74). Parenchymal tissue from chronic obstructive pulmonary disease (COPD) patients, who were current smokers, contained lower levels of total albumin, but had proportionally greater levels of Alb-CO, compared to patients with the normal lung function, as determined by ELISA and OxyBlot (25). Lung tissue from current smokers was also found to contain lower levels of albumin, which was highly carbonylated compared to lung tissue from ex-smokers and nonsmokers. Decreased levels of albumin in current smokers with COPD could contribute to the excessive accumulation of oxidants, which would ultimately lead to oxidative tissue injury. Lung tissue from ex-smokers had the same mean concentration of Alb-CO as nonsmokers (25), suggesting that smoking cessation could prevent albumin carbonylation, contributing to the restoration of the oxidant/antioxidant balance within the lung.
Oxidative stress is also involved in asthma. Carbonylation of sputum proteins in adult asthmatic patients and healthy controls was assessed by using OxyBlot (47). Albumin and α1-antitrypsin were specifically carbonylated in asthmatics, and Alb-CO was significantly correlated with sputum eosinophilia.
Recently, albumin carbonylation in different human diseases has been clearly documented by redox proteomics. Redox proteomics analysis, in combination with MS protein identification, of serum samples from patients afflicted with the autoimmune disease systemic lupus erythematosus showed albumin and immunoglobulins to be particularly carbonylated (42). In patients with stable disease activity, higher long-term protein carbonylation correlated with higher long-term disease activity, whereas patients with variable disease activity exhibited varying correlations between protein carbonylation and disease activity markers (42). These results confirm and extend previous findings (68) and further support a role for oxidative stress in this disease pathogenesis.
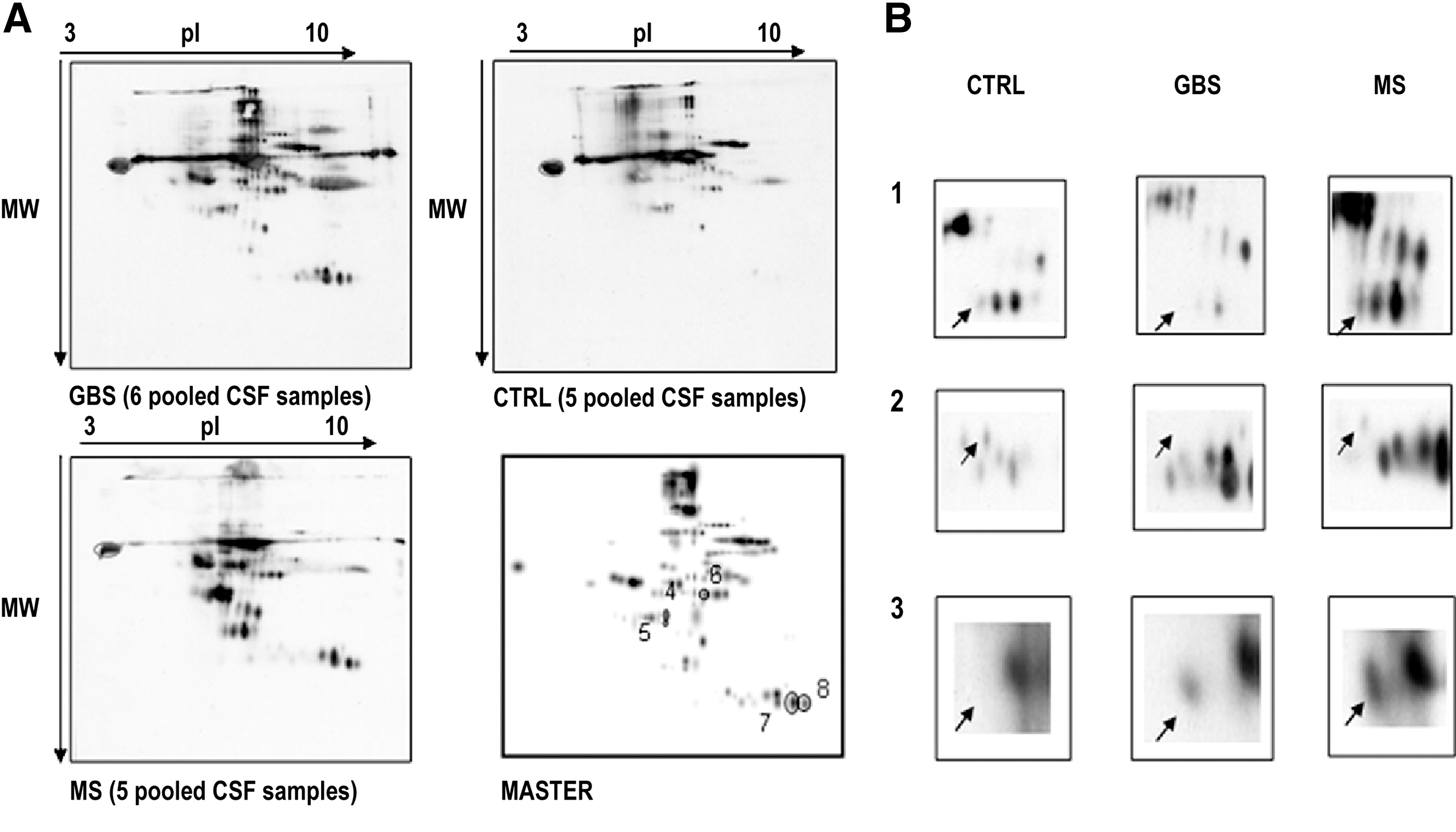
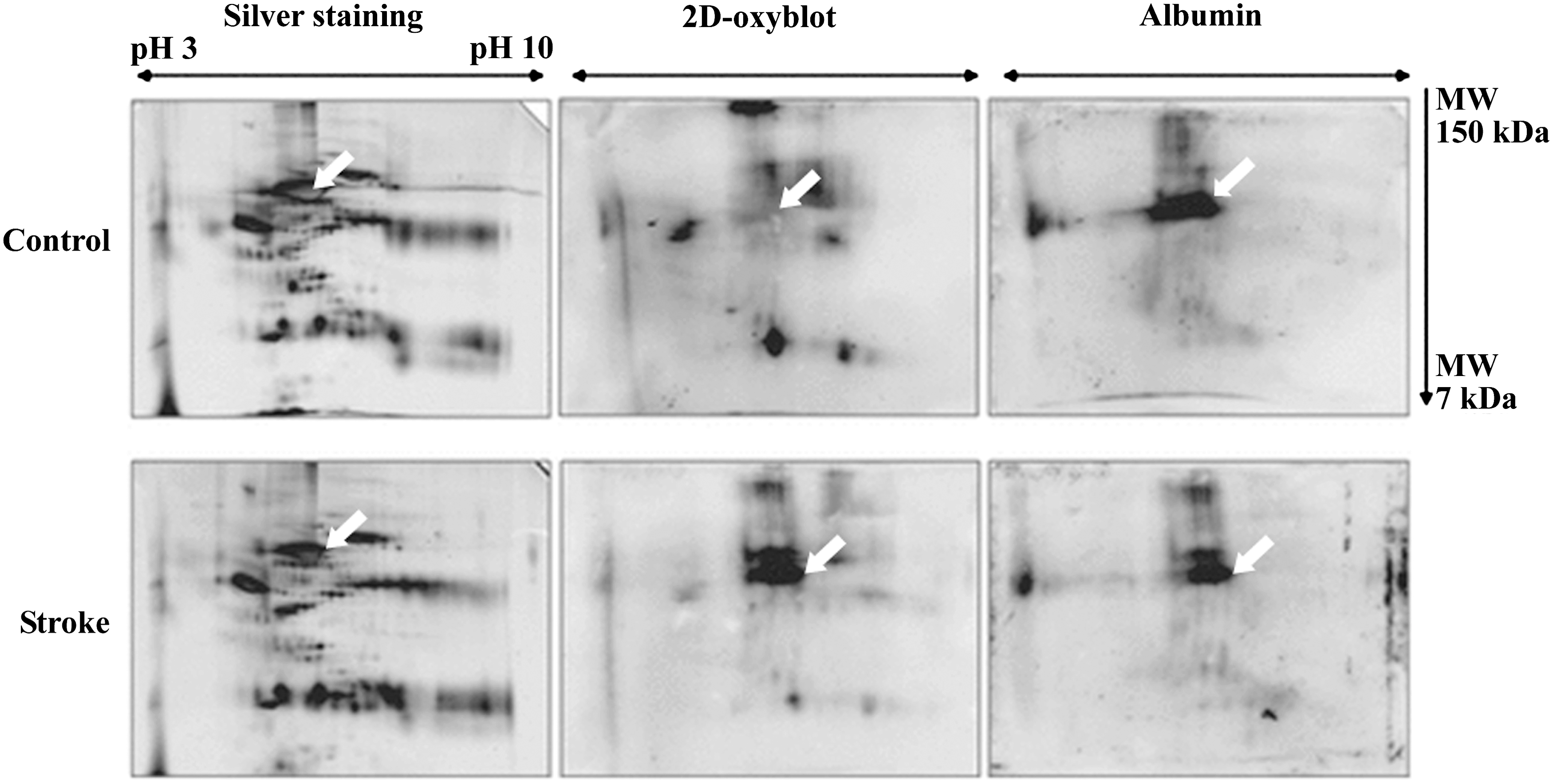
CSF is produced in the brain mainly by cells in the choroid plexus at a rate of 500 ml/day and contains ∼0.3% plasma proteins, mainly albumin and immunoglobulins. It is the only body fluid in direct contact with the brain. Thus, any changes in the CSF composition will reflect pathological processes and make CSF a potential source of biomarkers for different diseases. A recent study used redox proteomics to analyze the extent of carbonylation and, thus, of oxidative damage of CSF proteome in patients with Guillain–Barrè syndrome, an acute and immune-mediated inflammatory disease affecting the peripheral nervous system (13), evidencing a major sensitivity to carbonylation for albumin, α-glycoprotein, and Fab fragments from an IgM globulin (Fig. 3). Brain biomarkers of oxidative stress need to be identified to better understand and treat brain damage in human stroke patients. Redox proteomics was recently used to identify carbonylated proteins in the CSF of stroke patients with acute ischemic brain injury (41). Albumin was the only protein that was markedly carbonylated in the CSF from stroke patients as compared with controls (Fig. 4). Carbonylation of albumin normalized to the total albumin from each relevant group was significantly increased in CSF from stroke patients compared with the control group, and Alb-CO was significantly correlated to clinical outcomes. This finding in humans agrees with the experimental results obtained using an animal model of ischemic stroke. These results suggest that Alb-CO in CSF can be utilized as a biomarker of oxidative stress in stroke patients and animal models, thereof (41).
OxyBlot coupled to matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF)/MS mass fingerprinting identification of carbonylated proteins recently confirmed previous observations that albumin is the major plasma protein target of oxidative/carbonyl stress in chronic hemodialysis patients (54). This redox proteomic study also first showed that albumin carbonylation in uremic blood mainly results from protein modification by carboxymethyl-lysine, in agreement with the occurrence of carbonyl stress in uremia and hemodialysis patients. A further study demonstrated, by OxyBlot coupled to MALDI-TOF/MS mass fingerprinting and nano-LC-MS/MS analysis, that postdialysis plasma carbonylated-protein levels are significantly increased compared to predialysis levels, and that cabonylation targets almost a dozen of plasma proteins, among which albumin is the most susceptible to carbonyl formation (55). Furthermore, the in vitro finding that even low concentrations of Alb-CO may have proatherogenic properties indicates a potential role for protein carbonylation in the early atherogenic events of chronic uremia.
Significantly higher levels of protein carbonylation than in healthy controls were determined in BALF of children with interstitial lung disease, gastroesophageal reflux disease, and pulmonary alveolar proteinosis, although with significant interindividual variations. Surfactant protein A, α1-antitrypsin, and, mainly, albumin were the proteins most sensitive to carbonylation, whose extent was proportional to the count of neutrophils in BALF (71).
Albumin Nitration
Most data show that protein tyrosine nitration is a selective process where only a few proteins become nitrated, and only one or a few Tyr residues are modified within each protein. While HSA contains 18 Tyr residues, an in vitro study showed that only Tyr138 and Tyr411 are particularly susceptible to nitration (28).
Using a sensitive, quantitative ELISA, nitrated albumin (nitroalbumin) levels were assayed in plasma samples collected during the first hour, at day 1 and at day 4 of life from term newborns suffering from perinatal asphyxia (83). Nitroalbumin levels at day 1 were significantly increased in infants who developed moderate or severe encephalopathy compared to those who had a normal neurological evolution or developed mild encephalopathy. In contrast, the nitroalbumin concentration at day 1 was not associated with systemic complications (83). Plasma nitroalbumin at day 1 was thus specifically associated with perinatal hypoxic–ischemic cerebral injury. The nitroalbumin concentration increased with the severity of neonatal encephalopathy, adding support to the hypothesis that reactive nitrogen species (RNS) toxicity plays a role in hypoxic–ischemic brain injury. Therefore, plasma nitroalbumin seems to be a specific marker of neurological injury after perinatal asphyxia and is an easily measurable, noninvasive marker of nitrative stress usable as a secondary end-point in neuroprotective clinical trials. Increased plasma nitroalbumin is also associated with early transient hypoglycemia in preterm newborns, as determined by ELISA (82).
Advanced Oxidation Protein Products
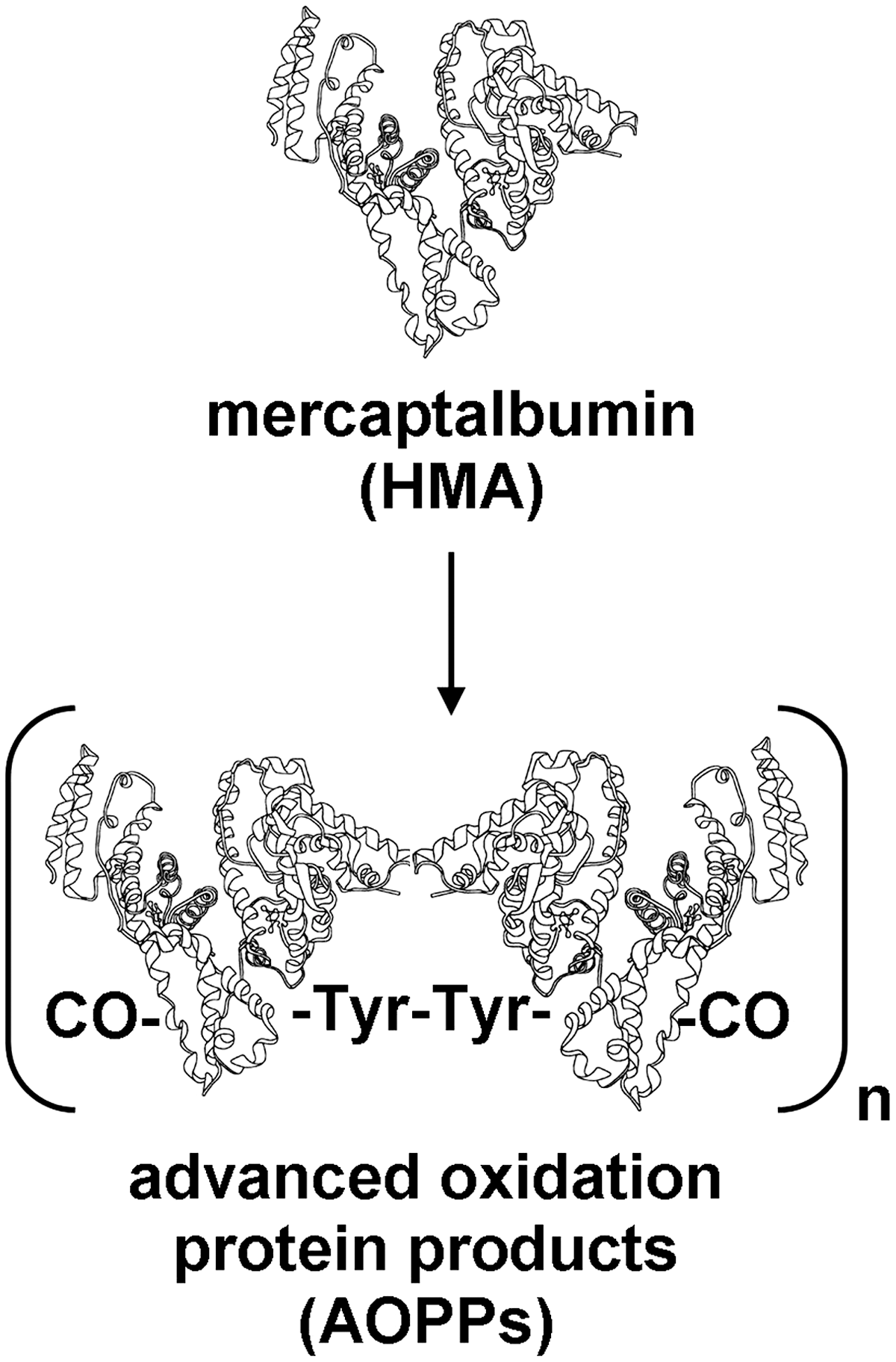
Elevated levels of advanced oxidation protein products (AOPPs), which arise from the reaction between chlorinated oxidants and plasma proteins, have been described in several human diseases (Supplementary Table S2) (67). AOPPs are dityrosine-containing and cross-linking protein products formed mainly during oxidative burst of activated neutrophils via the myeloperoxidase reaction. Plasma levels of AOPPs closely correlate with levels of dityrosine and pentosidine, an AGE product closely related to oxidative stress; indeed, AOPPs contain dityrosine, protein carbonyls, and pentosidine in an aggregate-forming or monomeric form (9) (Fig. 5). Utilizing indirect techniques, albumin was identified as the main AOPP product in plasma of chronic uremic patients (9). Notwithstanding, a lack of chemical structural completeness did not allow any conclusion. In addition, recent data appear to indicate that oxidized fibrinogen is the key molecule responsible for AOPP formation in the human plasma (66, 67). However, a role of AOPPs in atherosclerotic cardiovascular events in patients with chronic renal failure and, more in general, in the inflammatory process that is associated with uremia and hemodialysis is hypothesized (36, 67). In particular, in vivo and in vitro evidence that AOPPs are proinflammatory mediators has been provided (36), and pro-oxidant–proinflammatory properties of AOPPs were evidenced in animal models of chronic kidney failure (33, 35, 69), suggesting a vicious cycle with amplification of oxidative stress and inflammation (Fig. 6). In the adults with chronic renal insufficiency, AOPPs accumulate proportionally with the decline in renal function and are also proposed as predictors of survival in these patients (67).
Concluding Remarks
In vivo albumin oxidation in relation to various human diseases has been proven, by direct techniques, only recently. Demonstration of oxidized HSA in biological fluids has stimulated studies aimed at defining its levels in relation to various human diseases potentially linked to oxidative/nitrative stress. High oxidized HSA levels have been assayed in vivo in pathologies where reactive oxygen species/RNS seem to play a pathogenic role, thus demonstrating the HSA antioxidant role also in vivo. Moreover, HSA purified from hemodialysis patients had decreased the free radical-scavenging ability and reduced drug-binding properties, thus confirming that oxidation-induced structural changes affect HSA- binding properties and antioxidant activity (31, 38, 52). Disturbed binding/transport properties of oxidized albumin may impair intercellular/interorgan traffic of endogenous compounds, but also delivery of drugs that are crucial in the treatment of human disease, and this may at least in part explain the altered behavior of drugs in some pathological situations.
Indeed, although the albumin Cys34 thiol does not react particularly fast with oxidants, it can still be considered an important intravascular scavenger due to its very high concentration in human plasma, particularly considering that plasma is scarce in antioxidant defenses and low molecular weight thiols. In this regard, the increase of oxidized HSA forms in pathological conditions can be considered as a proof of concept of the scavenger activity of albumin.
Approximately 10 g/h of HSA travels in and out of blood vessels (29); this means that within an hour, approximately half of intravascular HSA is replaced with extravascular HSA. As albumin is quickly distributed between blood and the extravascular compartment, it may serve as a global marker for the redox state in the body. Interestingly, nitroalbumin has been reported to be transported across the endothelial barrier two to four times faster than native albumin (58).
Current research progress on the detailed measurement of oxidative damage to proteins by different redox proteomics and MS technologies suggests that many more biomarkers are likely to be developed and clearly validated in the near future. As a whole, most data here presented demonstrate that massive albumin oxidation occurs in vivo in different biological fluids, and, to some extent, that this process is correlated to organ dysfunction in human pathology. We hope that these findings put the basis for developing/applying specific analytical techniques for the characterization of oxidized albumin forms in screening studies, also considering that in a number of studies, albumin is usually ignored, since it is generally removed from plasma/serum, CSF, or other biological tissue fluids to detect lower-abundance proteins. However, the potential use of albumin as a biomarker is presently not fully clear: is albumin oxidation only a marker of oxidative stress, or could it be reasonably considered a specific disease marker? In that light, the finding that nitroalbumin concentration is directly correlated with the severity of neurological morbidity after perinatal asphyxia indicates that nitroalbumin seems to be a reliable, early, and easily obtainable marker of long-term effects on the cognitive and behavioral consequences for the survivors of moderate-to-severe neonatal encephalopathy, beside a marker of RNS generation and nitrative stress occurrence (83). Analogously, the finding that Alb-CO was the unique carbonylated proteins found in CSF of patients with acute ischemic stroke, whose increased levels were positively correlated with neurologic indicators (41), suggests that HSA carbonylation in CSF after stroke may be able to act not only as a biomarker providing information of oxidative stress status but also as an important clue that will allow interpretation of the diagnostic outcomes of stroke.
A clear benefit of using oxidized albumin as a biomarker is that the highly abundant HSA concentration in most biological fluids allows the investigation of oxidative/nitrative stress in clinical practice. On the other hand, its high concentration in these fluids may also represent a limitation of using oxidized albumin as a biomarker, because one may wonder whether or not it is an intentional free radical scavenger or if it is simply targeted because of its abundance and high concentration. In this respect, the fact that, surprisingly, nitrated albumin is not a frequent finding in human pathology, despite HSA is the most abundant protein in most biological fluids on a molar basis and contains 18 Tyr residues, may suggest specificity/selectivity and intentionality. Furthermore, the finding that the relative content of 3-nitrotyrosine on the albumin molecule was very low when compared with that of other much less concentrated plasma proteins, such as fibrinogen and complement factor D (57), may corroborate this hypothesis.
In conclusion, it is clear from the most recent publications that redox albuminomics is increasingly used to analyze oxidative albumin modifications occurring in vivo. This approach has a clear applicability to diseases where oxidative stress is involved in their pathophysiology. Occurrence of oxidized albumin in human body fluids easily obtainable without invasive procedures makes oxidized albumin forms potentially useful as diagnostic biomarkers. In addition, because the albumin half-life in plasma is ∼20 days, increases in its oxidized forms may be indicative of relatively recent oxidative events in human body, and oxidized albumin forms might serve as early biomarkers for human diseases associated to oxidative stress. The challenge in the near future will be to analyze how these oxidative albumin modifications are related to real impact to the body. It will have to be clarified if albumin oxidation can be disease specific or if it just reveals a disease nonspecific increase in oxidative stress as well as if the extent of albumin oxidation can be a possible discriminant to recognize different diseases or, more likely, different stages of a disease. In this sense, the finding that HNA-1 and HNA-2 levels increase in correlation with the degree of renal dysfunction (75, 76), which in turn is paralleled by enhanced oxidative stress, suggests that the redox state of HSA could be a good marker to investigate the progression of, at least some, oxidative stress-related human diseases. Anyway, this is still an open question.
Footnotes
Acknowledgments
Owing to the strict space limitations, we could not cite all the works relevant to the presented topic. We sincerely apologize to those authors whose work we could not include. ![]() was prepared using and combining medical cliparts and illustrations available within the Servier Medical Art section, by courtesy of Servier International.
was prepared using and combining medical cliparts and illustrations available within the Servier Medical Art section, by courtesy of Servier International.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.