
Abstract
Introduction
Innovation
This study provides novel data with regard to a mechanism for the regulation of the microRNA, miR-21, by reactive oxygen species (ROS) in prostate cancer. We demonstrate that NADPH oxidase activity and miR-21 expression are high in prostate cancer, where they serve as an important regulator of cell invasiveness. Inhibition of ROS by inhibition of NADPH oxidase or reducing miR-21 expression decreased the metastatic phenotype of prostate cancer cells. Overall, the data presented provide a novel mechanism by which ROS can influence the metastatic potential of prostate cancer cells and identify discrete targets for new drug development.
MicroRNAs miRs are 18∼22 nucleotides long non-coding RNAs that are endogenously expressed in the cells (19). They regulate expression of the proteins by suppressing translation or causing degradation of mRNAs by binding to their 3′- untranslated region (UTR) (30). In cancer, several miRNAs are deregulated, resulting in both tumor suppression and tumor promotion (56). One of these miRs, miR-21, an onco-miR (38) is up-regulated in a number of cancers, including prostate cancer, suggesting that this miR could play a pivotal role in oncogenesis (49). In breast cancer, miR-21 promoted the growth of breast cancer cells by down-regulating tumor suppressor genes, such as programmed cell death 4 (PDCD4), maspin, and tropomyosin 1 (TPM1) (41, 59). PDCD4 was also shown to be involved in the miR-21-regulated colon cancer intravasation and metastasis (4). In hepatocellular and ovarian cancer cells, miR-21 increased proliferation and metastasis by decreasing phosphatase and tensin homolog deleted on chromosome 10 (PTEN) (27, 31); while in glioma cells, it increased invasion by regulating the matrix metalloproteinase regulators (16).
Prostate cancer cells exhibit high levels of reactive oxygen species (ROS), which augment cell proliferation and metastasis (43). Increased levels of ROS are observed in androgen-dependent (LNCaP) and -independent (PC3 and DU145) prostate cancer cell lines that are exposed to prostate-specific antigen and testosterone in cultures (45). NADPH oxidase appears to be the major source of ROS in these cells, whose activity is induced by extracellular signal-regulated kinase 1/2 (ERK1/2), a member of the mitogen-activated protein kinase (MAPK) family (17, 18). In DU145 prostate cancer cells, ROS derived from NADPH oxidase provided trophic support and reduced apoptosis of these cells (7). However, the mechanism(s) underlying ROS-dependent proliferation and metastasis of prostate cancer cells still remain unclear.
In the present study, we show that ROS produced by NADPH oxidase plays a critical role in the induction of miR-21 in prostate cancer cells. The data suggest that miR-21 could serve as a mediator of ROS-dependent progression of prostate cancer to an invasive and metastatic phenotype, and thus provide an ideal target for the development of anti-cancer therapies.
Results
miR-21 regulates the invasiveness of PC-3M-MM2 prostate cancer cells
Previous studies have shown that miR-21 is over-expressed in prostate cancer (23, 34, 49). The androgen-dependent LNCaP cells are less invasive than the androgen-independent DU145 and PC3 cells (22). Furthermore, PC-3M-MM2 cells (derived from bone metastatic cultures) are the highly invasive variant of PC3 cells (11). To determine whether miR-21 expression correlates with invasiveness of prostate cancer cells, we examined its expression in LNCaP, DU145, and PC-3M-MM2 cells. The levels of miR-21 were 1.0±0.09, 6.18±0.31, and 8.03±0.58 arbitrary units for LNCaP, DU145, and PC-3M-MM2 cells, respectively (see Supplementary Fig. S1A; Supplementary Data are available online at
To determine the consequence of miR-21 over-expression, we examined the effect of its down-regulation by anti-miR-21 oligonucleotides on the invasiveness of PC-3M-MM2 cells. First, the effectiveness of the anti-miR-21 in reducing the expression of miR-21 was determined by quantitative real-time polymerase chain reaction (qRT-PCR) studies after transfecting cells with anti-miR-21 for 24 h. Figure 1A shows dose-dependent reductions in the miR-21 expression by anti-miR-21. The levels of miR-21 in the presence of 12.5, 25, and 50 nM anti-miR-21 were reduced to 0.42±0.04, 0.30±0.04, and 0.08±0.02–fold (mean±standard error), respectively. Anti-miR-21 also increased levels of PDCD4 and maspin, which are negatively regulated by miR-21 (59). PDCD4 levels were increased to 180%±28% and 163%±13%, while the maspin levels were increased to 155%±15% and 149%±10%, at 25 and 50 nM anti-miR-21 concentration, respectively (Fig. 1B, C). Control cells were transfected with 5 nM scrambled anti-miR (anti-miR-scr) and were assumed to be 100%. Increasing the concentrations of anti-miR-scr to 50 nM did not affect the levels of miR-21 expression or the levels of PDCD4 and maspin (data not shown). We also observed high basal levels of both PDCD4 and maspin in immortalized prostate epithelial cells (RWPE-1), as compared with the PC-3M-MM2 (Fig. 1B, C). Since miR-21 binds to the 3′-UTRs of PDCD4 and maspin to down-regulate their expression, we determined the transcriptional regulation of both these proteins in the presence of anti-miR-21 by luciferase assays (59). Cells exposed to anti-miR-21 showed increased PDCD4 and maspin luciferase activities, which are indicative of reductions in the inhibitory effects of miR-21 by anti-miR-21. The activities of PDCD4 and maspin 3′-UTRs were 132%±7% and 124%±7% of the control, respectively, in the anti-miR-21 group (Fig. 1D). These data confirm previous observations that miR-21 serves as a negative regulator of the expression of both PDCD4 and maspin (23, 59).
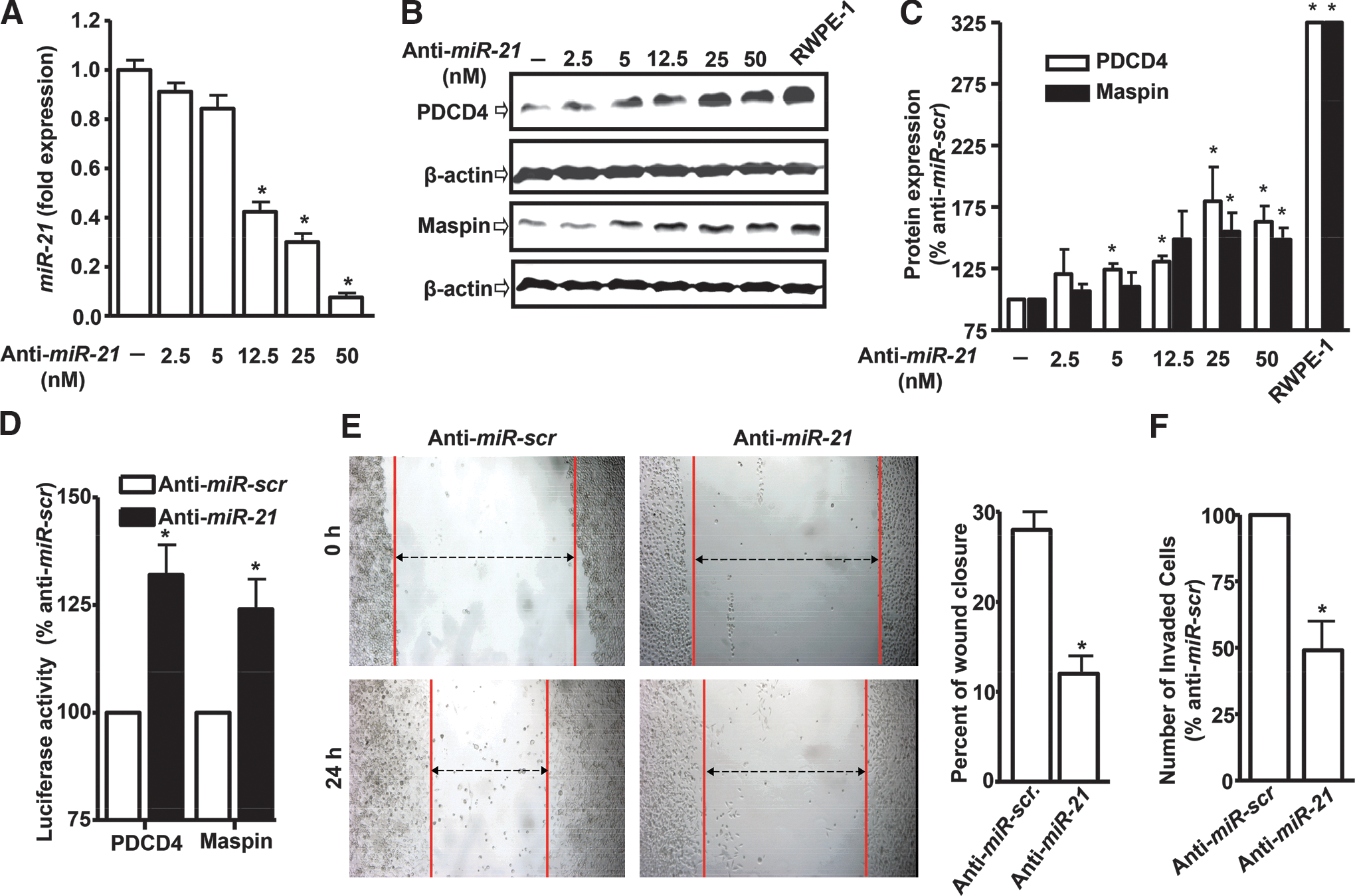
Since miR-21 regulates the invasiveness of various cancer cells (4, 23, 27, 59), we next determined whether it has similar functions in PC-3M-MM2 prostate cancer cell lines. Cell invasion was assessed by a modified wound-healing and matrigel invasion assays. Cells were first transfected with 50 nM anti-miR-scr or anti-miR-21 for 24 h and then plated for these assays. As shown in Figure 1E, anti-miR-21 reduced migration in wound-healing assays, as compared with anti-miR-scr cells. The percent of wound closure was 28%±2% in anti-miR-scr-transfected cells, while it was 12%±2% in anti-miR-21-transfected cells (Fig. 1E). Similarly, anti-miR-21 reduced invasion by ∼50%, compared with anti-miR-scr control group in matrigel assays (Fig. 1F). Thus, similar to an earlier study in DU145 and PC3 prostate cancer cells (23), miR-21 regulates invasiveness of PC-3M-MM2 prostate cancer cells.
miR-21 regulates the in vivo metastasis of prostate cancer cells
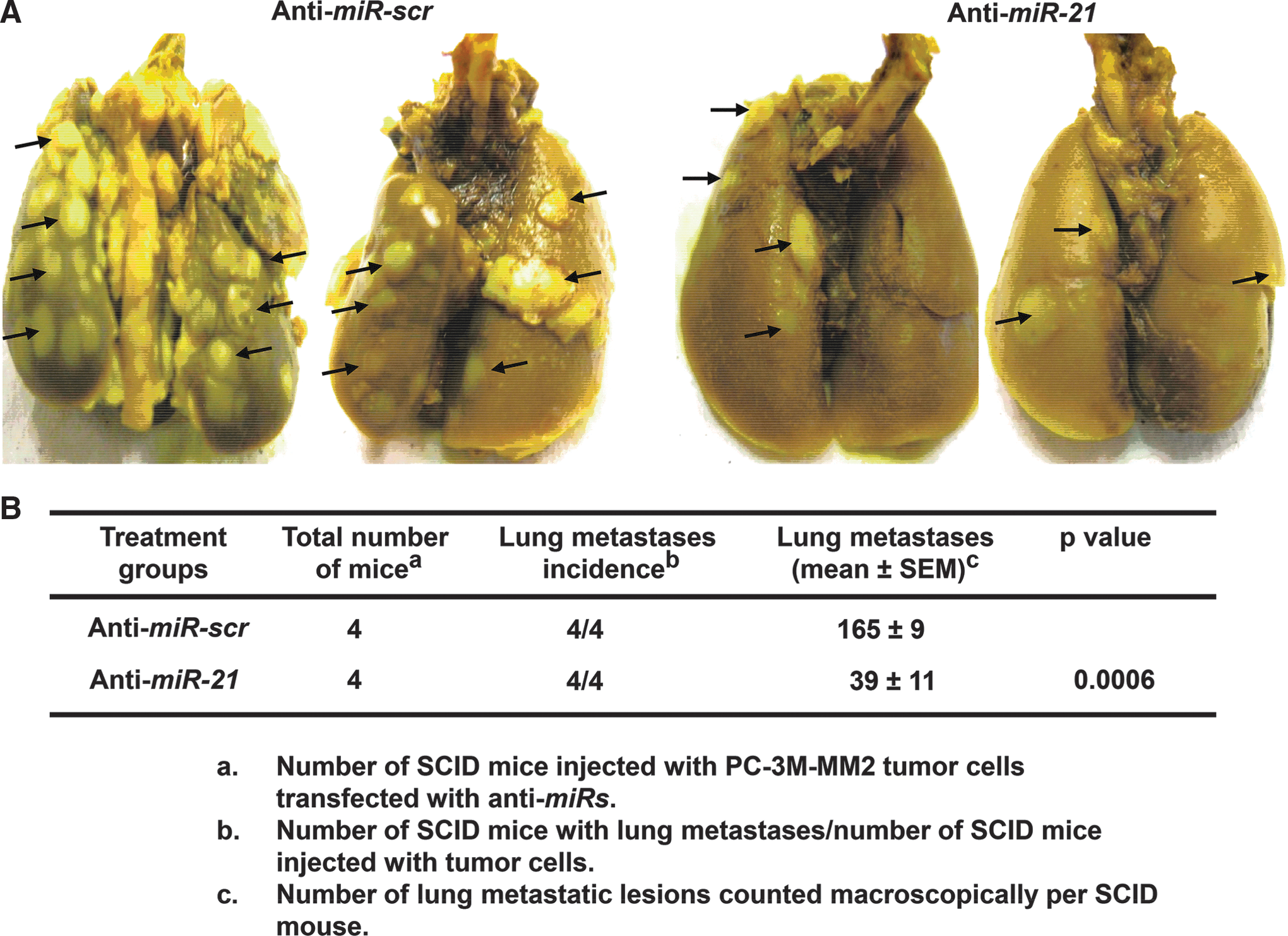
To determine whether miR-21 promotes in vivo tumor metastases, lung metastases were induced by injecting PC-3M-MM2 cells via dorsal tail vein into severe combined immunodeficient (SCID) mice. Twenty-four hours before injections, these cells were transiently transfected with either 50 nM anti-miR-scr or anti-miR-21. The anti-mir-21 group exhibited significantly less lung metastatic lesions, compared with mice injected with anti-miR-scr cells (Fig. 2A). The numbers of lung metastatic lesions were 165±9 and 39±11 in animals injected with the anti-miR-scr and anti-miR-21 cells, respectively (Fig. 2B). These findings indicate that miR-21 regulates in vivo metastasis of prostate cancer cells.
miR-21 expression in prostate cancer cells is dependent on high ROS generation
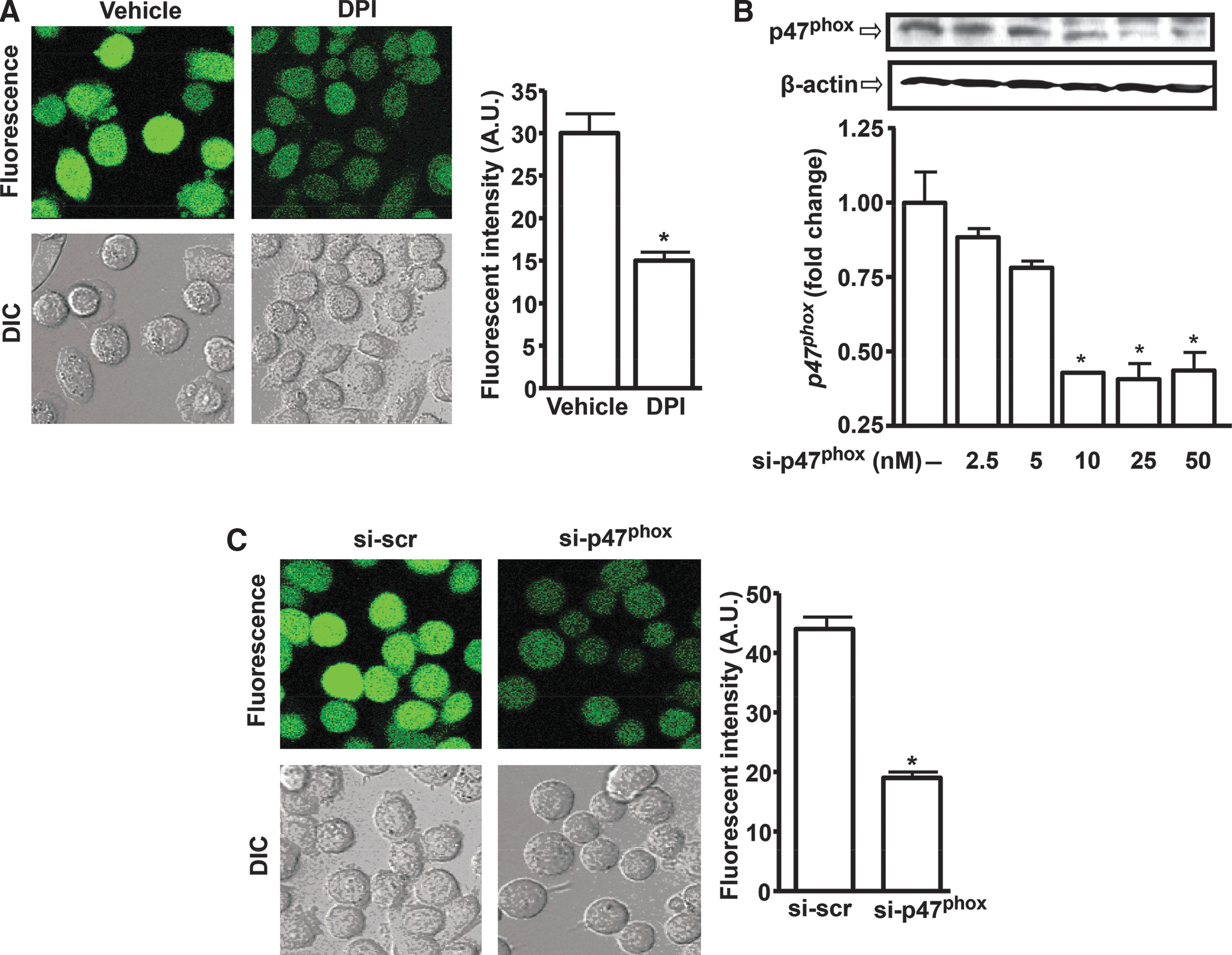
Prostate cancer cells exhibit high ROS generation, mediated by the NADPH oxidase enzyme system (17). We hypothesized that ROS promotes miR-21 expression in prostate cancer cells. Figure 3A shows that PC-3M-MM2 cells expressed high levels of ROS, as determined by 2′,7′-dichlorofluorescein diacetate (H2DCFDA) fluorescence. Treatment of these cells with Diphenyleneiodonium chloride (DPI), a known chemical inhibitor of NADPH oxidase (33), reduced ROS generation from 30±2 to 15±1 arbitrary units (Fig. 3A bar chart). To determine whether NADPH oxidase is essential for ROS generation in prostate cancer cells, PC-3M-MM2 cells were transfected with different concentrations of small interfering RNA (siRNA) against p47phox (si-p47phox), an essential subunit of NADPH oxidase activity (21). Maximal inhibition of p47phox levels was observed at 25 nM si-p47phox, and this concentration was used for future experiments (Fig. 3B). The reductions in p47phox levels by si-p47phox significantly decreased ROS generation by these cells. ROS generation averaged 19±1 arbitrary units in the si-p47phox group, compared with 44±2 arbitrary units in the scrambled siRNA (si-scr; 25 nM) group (Fig. 3C).
We next determined whether suppression of ROS generation could affect miR-21 expression. Treatment of PC-3M-MM2 cells with 5 μM DPI for 24 h reduced the expression of miR-21 to 0.46±0.02 fold from vehicle-treated controls (Fig. 4A). In addition, N-acetyl-L-cysteine (NAC, 500 μM) reduced miR-21 expression to 0.50±0.07 fold of control cells (Supplementary Fig. S2A). However, 2-4-carboxyphenyl-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide (C-PTIO, 100 μM), a nitric oxide scavenger, did not produce any change in miR-21 expression (Supplementary Fig. S2B). These results suggest that reactive nitrogen species are not involved in regulating miR-21 levels in prostate cancer cells.
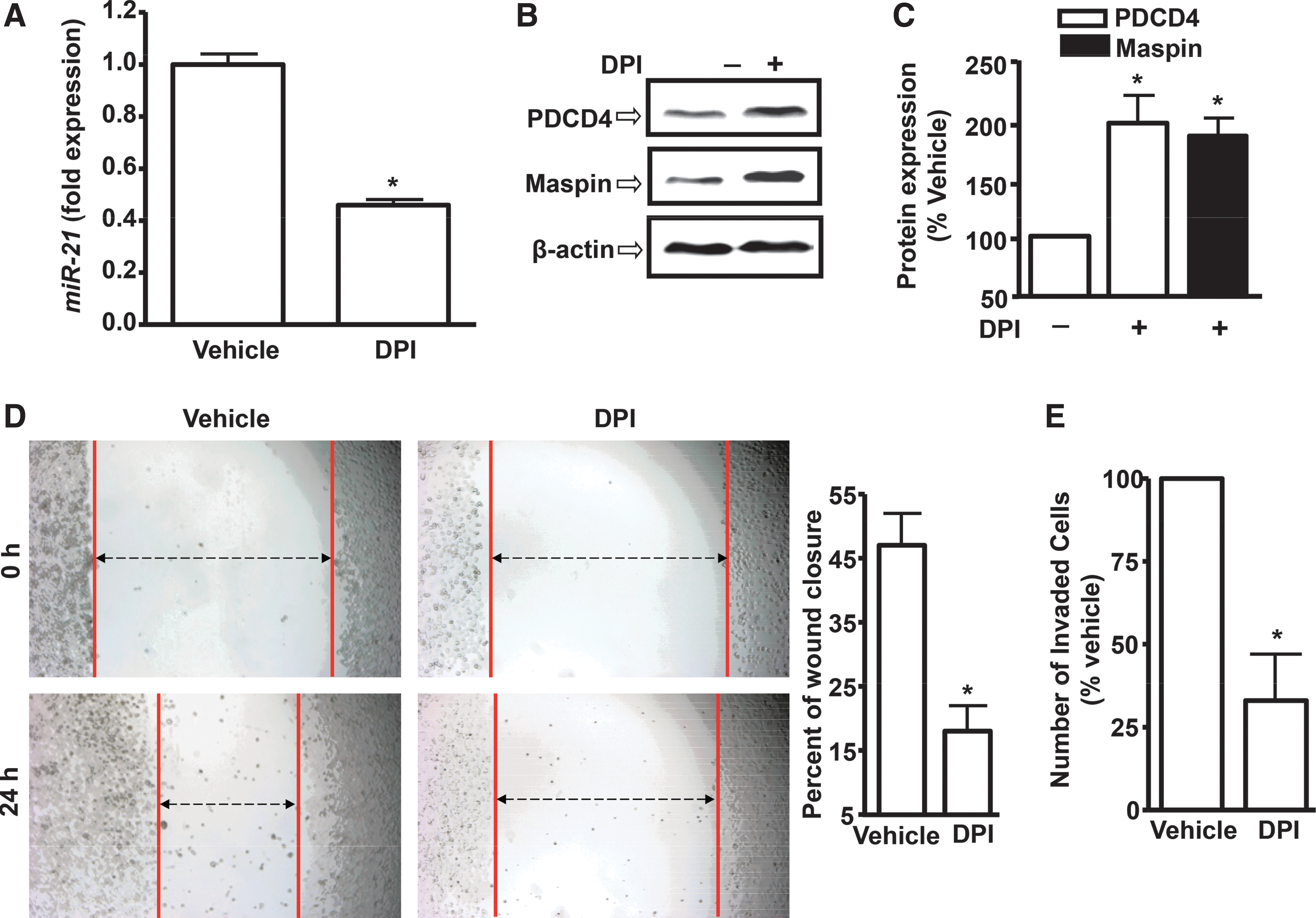
DPI treatment also increased PDCD4 and maspin expression. Their respective levels in the presence of DPI were 196%±24% and 188%±15% of vehicle-treated controls (Fig. 4B, C). DPI and NAC also decreased the expression of miR-21 in DU145 prostate cancer cells (Supplementary Fig. S3). Functionally, DPI decreased the wound closures, which are indicative of reduced migration of prostate cancer cells (Fig. 4D). Furthermore, DPI suppressed invasion of prostate cancer cells in matrigel assays. The percentage of invading cells was reduced to 33%±14% by DPI (Fig. 4E).
Inhibition of p47phox expression by si-p47phox decreased miR-21 expression. miR-21 levels were 1.00±0.04, 0.49±0.05, and 0.59±0.01 fold in the presence of si-scr and 25 and 50 nM si-p47phox, respectively (Fig. 5A). A similar effect was observed after knockdown of another NADPH oxidase subunit, p22phox. Transfection of these cells with si-p22phox (5 nM) reduced the expression of p22phox to 0.59±0.07 fold and the levels of miR-21 to 0.63±0.10 fold (See Supplementary Fig. S4A, B). si-p47phox also increased the levels of both PDCD4 and maspin. In the presence of 25 nM si-p47phox, PDCD4 and maspin levels were 200%±27% and 151%±13% of si-scr group, respectively (Fig. 5B, C). Additional experiments tested the effect of NADPH oxidase in regulating the binding of miR-21 to PDCD4 and maspin 3′-UTR by measuring luciferase activity. In the presence of si-p47phox (25 nM), the luciferase activity was 143%±13% and 142%±10% for PDCD4 and maspin, respectively (Fig. 5D), with si-scr (25 nM) controls as 100%. These results demonstrated that miR-21 expression and its transcriptional activity are regulated by NADPH oxidase-mediated ROS generation in prostate cancer cells.

In order to determine the functional impact of NADPH oxidase on PC-3M-MM2 cell motility, we studied the effects of si-p47phox on the migration and invasion of these cells. Transient transfections of PC-3M-MM2 cells with si-p47phox suppressed wound closures from 44%±4% (for 25 nM si-scr-transfected cells) to 17%±2% (for 25 nM si-p47phox transfected cells) (Fig. 5E). Similarly, si-p47phox also decreased prostate cancer cell invasion in matrigel assays. The number of invading cells was 45%±9% of the si-scr control cells (Fig. 5F). These results suggest that NADPH oxidase regulates miR-21 expression and function in PC-3M-MM2 prostate cancer cells.
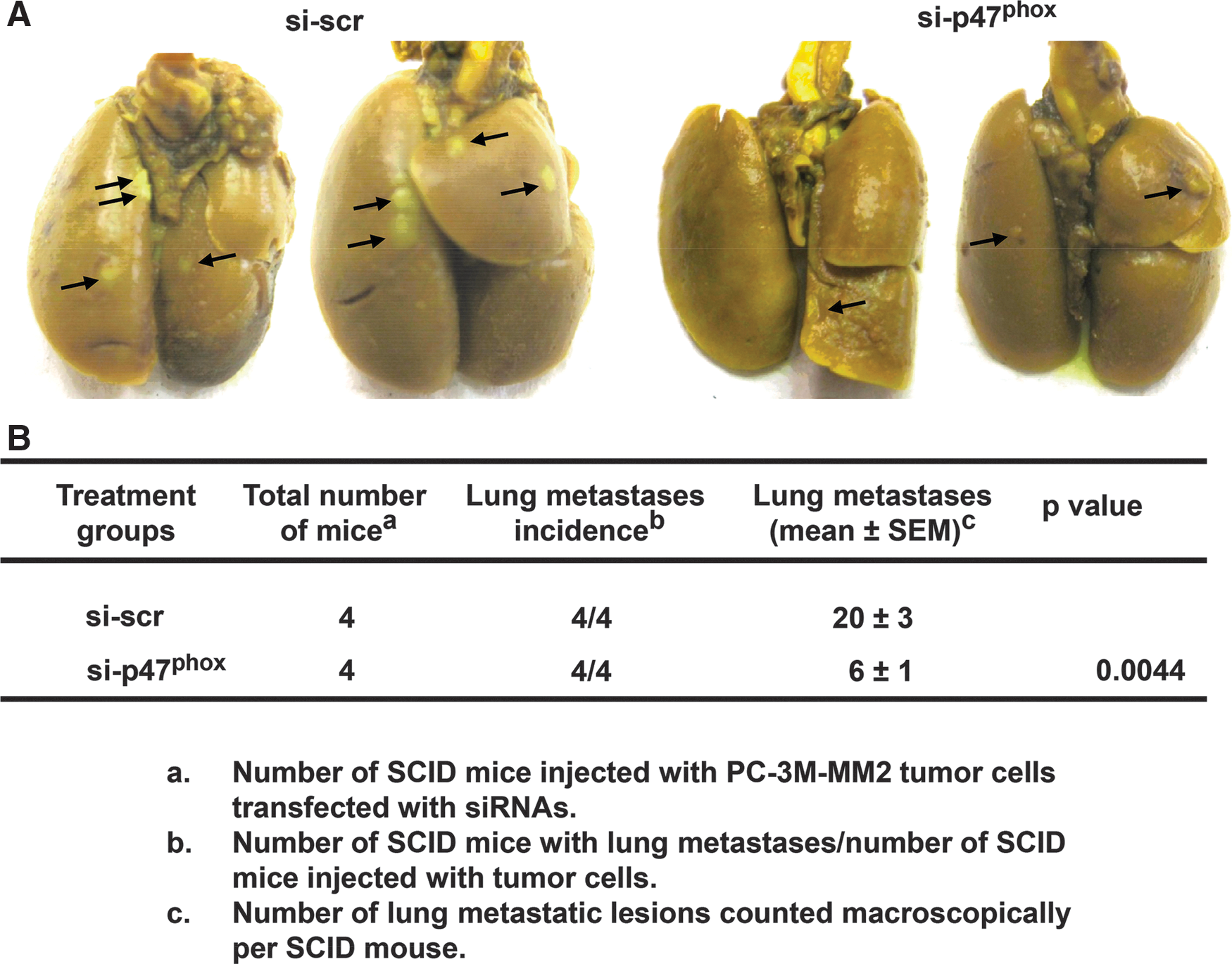
p47phox siRNA suppressed lung metastases of PC-3M-MM2 cells in a xenograft model of prostate cancer
Since NADPH oxidase regulates migration and invasiveness of prostate cancer cells in cultures, and miR-21 regulates these functions, we tested whether inhibition of NADPH oxidase reduces tumor metastasis in a xenograft model of prostate cancer. SCID mice injected (via tail vein) with PC-3M-MM2 cells transfected with 25 nM si-scr or si-p47phox showed 20±3 and 6±1 metastatic lung lesions per mouse, respectively (Fig. 6A, B). These studies indicate that NADPH oxidase activity regulates metastasis of PC-3M-MM2 prostate cancer cells to the mouse lungs and, based on the previous results, might suggest a role of miR-21 in this process.
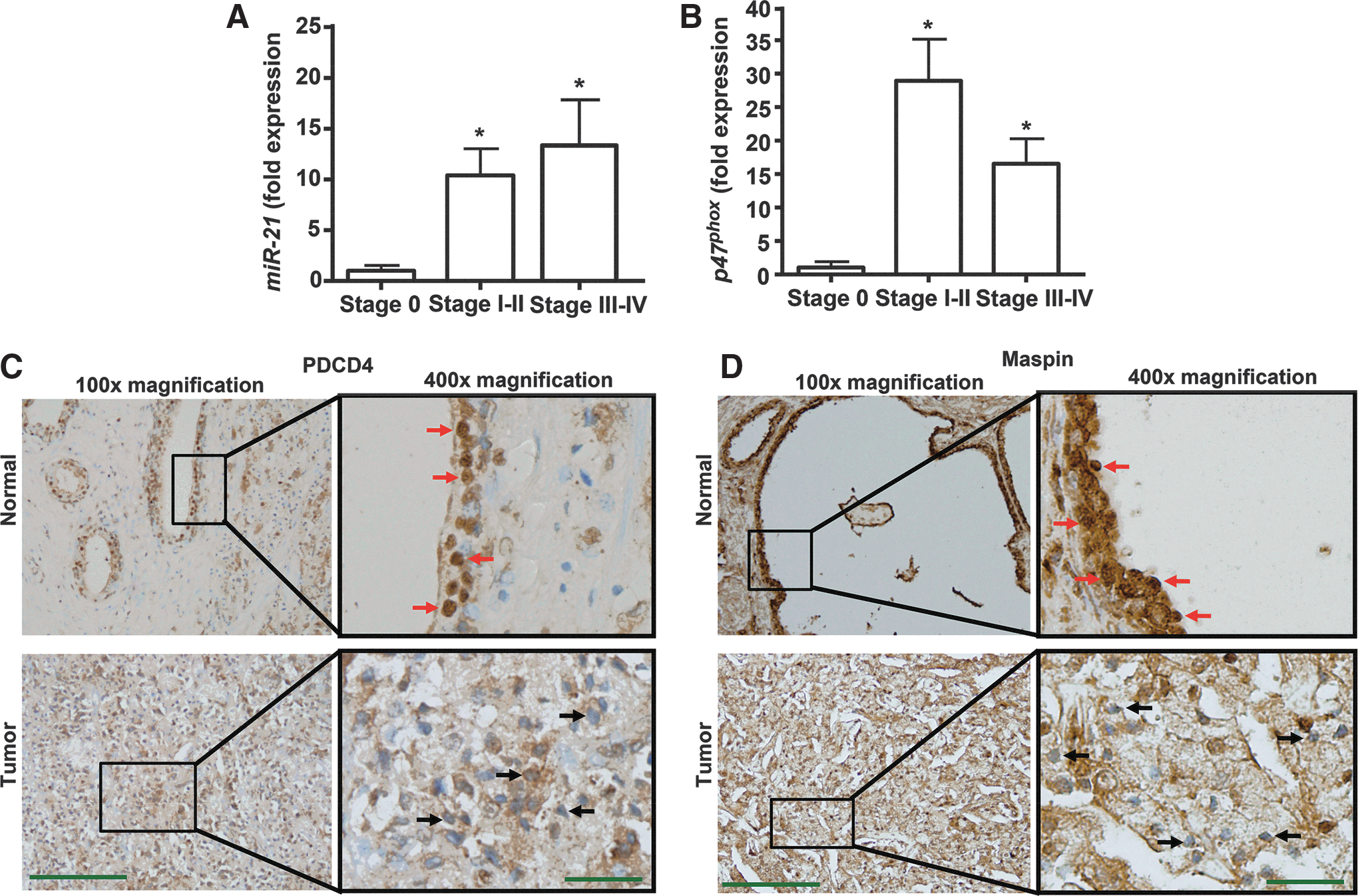
Human prostate cancer specimens demonstrate increased expression of miR-21 and p47phox
Human prostate cancer RNA samples were obtained from TissueScan qRT-PCR panels (Origene Technologies, Rockville, MD). Figure 7A shows that increased expression of miR-21 was observed in samples classified as Stages I-II (localized to the prostate) and Stages III-IV (spread to nearby or distant organs), as compared with the Stage 0. The expression levels were 1.0±0.53 (n=4), 10.4±2.6 (n=9), and 13.4±4.6 (n=5) fold for Stages 0, I-II, and III-IV, respectively (Fig. 7A). Similarly, we also observed increased expression of p47phox in different stages of prostate cancer, as compared with Stage 0. However, the levels of p47phox in Stages I-II and III-IV were 29±6 fold and 17±4 fold, respectively (Fig. 7B). PDCD4 and maspin were next determined by immunohistochemistry in Stage IV human prostate cancer formalin-fixed, paraffin-embedded sections. Diaminobenzidine tetrahydrochloride (DAB) staining of these tissues revealed high levels of both PDCD4 (Fig. 7C) and maspin (Fig. 7D) in the normal prostate cells, but reduced immunolabeling in the tumor cells (Fig. 7C, D). In normal conditions, PDCD4 is primarily localized in the nucleus (6). PDCD4 showed a predominant nuclear localization in normal prostate epithelial cells but not cancer cells, as indicated by the increased dark brown staining (red arrows). Similar results were found in colorectal carcinoma (32), in which reduced nuclear localization of PDCD4 was proposed as a biomarker for the disease (13).
ROS regulates miR-21 by activating Akt
A previous study indicated that ROS could regulate invasiveness by activating Akt in prostate cancer cells (18). Akt was shown to regulate miR-21 in cardiac myocytes (36) and prostate cancer cells (40), suggesting that it could serve as a target of ROS for regulating miR-21. Treatment of PC-3M-MM2 and DU145 cells with DPI or NAC led to reduced phosphorylation of Akt (p-Akt, Supplementary Fig. S5A, B). LY294002, a known PI3K inhibitor, also reduced the levels of p-Akt (Supplementary Fig. S5B). These data suggest that ROS activation of Akt could account for the induction of miR-21 expression in prostate cancer cells.
Discussion
The principal finding of this study is that ROS are a major contributor to the high expression of miR-21, in prostate cancer cells. The main source of ROS generation is the NADPH oxidase system, whose activity is elevated in the prostate cancer cells (17). Functionally, high ROS, through its regulation of miR-21, contribute to increased metastatic potential of the prostate cancer cells. We also demonstrate constitutively high expression of NADPH oxidase subunit (p47phox ) and miR-21 in different stages of human prostate cancer. Finally, our data suggest that ROS activate Akt pathway to increase miR-21 expression. These findings provide a better understanding of the mechanism by which oxidative stress in prostate cancer cells promotes increased invasiveness and suggest that ROS and miR-21 could serve as useful targets for treating prostate cancer metastasis.
Our data show that the invasiveness of prostate cancer cell lines is dependent on miR-21 expression. For example, the highly aggressive PC-3M-MM2 cells show higher levels of miR-21 expression, as compared with LNCaP cells, which demonstrates a lower invasive phenotype and shows lower expression of miR-21. We previously showed that NADPH oxidase activity in prostate cancer cells is dependent on the ERK MAP kinase activity (17). Accordingly, ERK1/2 mediates phosphorylation of p47phox subunit of the NADPH oxidase, which promotes its activation (17). In this study, we further expand these findings to show that the invasive phenotype dictated by high NADPH oxidase activity is dependent on miR-21. This is supported by the finding that anti-miR-21 significantly suppressed the migration of these cells in cultures and reduced the number of lung metastases.
We demonstrate that NADPH oxidase activity is essential for miR-21 expression. p47phox is a subunit of NADPH oxidase, which is critical for its activation (21). Inhibition of p47phox (by siRNA) in PC-3M-MM2 cells not only reduced their metastatic potential, but also lowered the miR-21 levels. Similarly, down-regulation of p22phox by siRNA also reduced miR-21 expression. Furthermore, treatment of prostate cancer cells with DPI, a chemical inhibitor of NADPH oxidase, or the antioxidant NAC, reduced the expression of miR-21. These data strongly support the role of ROS and the NADPH oxidase system in the regulation of miR-21 expression in prostate cancer cells.
Only limited information is available with regard to the upstream regulators of miR-21 expression. For example, interleukin-6 (IL-6) increased miR-21 expression by stimulating signal transducers and activators of transcription 3 in multiple myeloma cells (26). Treatment of promyelocytic cells with 12-myristate 13-acetate (PMA) resulted in increased transcription of miR-21 via activator protein 1 (AP-1) (15). It is known that AP-1 is negatively regulated by downstream miR-21 target PDCD4 (54), which itself is down-regulated by miR-21 (14). Accordingly, AP-1-mediated activation of miR-21 resulted in reduced PDCD4 levels, which led to increased AP-1 activity and proliferation in thyroid cells (47). In addition to these, the upstream regulators of miR-21 also include epithelial growth factor receptor (37), nuclear factor-κB (58), estrogen receptor (5, 51), bone morphogenic protein-6 (12), transformation growth factor-β (10), and Foxo3a (50). In prostate cancer, an androgen receptor-dependent regulation of miR-21 has been suggested (35). These authors showed the presence of a highly conserved androgen-response element (ARE) in the miR-21 promoter. Furthermore, they showed that apart from androgen-dependent growth, miR-21 was also involved in the growth regulation of androgen-independent cells. Recent studies have implicated Akt in the regulation of miR-21 in prostate cancer cells (40) and cardiac myocytes (36). The current study shows that Akt could mediate ROS regulation of miR-21 expression in prostate cancer cells. Interestingly, miR-21 has been shown to increase ROS generation by decreasing the activity of superoxide dismutase 3 (SOD3) or by indirectly inhibiting SOD2 levels in cancer cells (57), suggesting the existence of an ROS-driven positive feedback loop in cancer cells to promote miR-21 expression. Overall, these data suggest that miR-21 serves as an integrator of signals derived not only from the androgen/androgen receptor axis (35) but also from multiple factors in the tumor microenvironment and intracellular environment.
Our data showing oxidative stress-dependent regulation of miR-21 expression in prostate cancer is supported by studies in vascular smooth muscle cells and cardiac myocytes, which showed increased expression of miR-21 by hydrogen peroxide (H2O2) (8, 24). miR-21 levels were also up-regulated in mouse macrophage cell line, RAW264.7 (48). However, our study is the first to document a role of NADPH-oxidase-mediated ROS generation in regulating miR-21 expression and the invasiveness of prostate cancer cells.
Several miRs have recently been shown to be regulated by oxidative stress (44, 50). MiR-200c, along with other members of miR-200 family, was up-regulated in the presence of H2O2-induced oxidative stress in human umbilical vein endothelial cells (28). This H2O2-mediated increase in miR-200c involved p53 and retinoblastoma proteins, which further led to down-regulation of miR-200c target ZEB1, and increased apoptosis and senescence of the endothelial cells (28). Oxidative stress-induced up-regulation of miR-200 family of miRs was also seen in various human cancer cells such as ovarian (SKOV3), melanoma (MDA-MB-435S), and breast (MDA-MB-436 and BT-549) (29). In particular, miR-141 and miR-200a targeted p38α MAPK in ovarian adenocarcinoma, resulting in its down-regulation and thus increased tumor growth (29).
The role miR-21 plays in promoting cell survival and metastasis has been delineated in various studies. Downstream targets of miR-21 that could explain its pro-invasive function include tumor suppressor genes, such as PDCD4, maspin, TPM1, and PTEN. miR-21 negatively regulates the expression of these targets, thereby reducing their tumor suppressor functions (38). PDCD4 acts by inhibiting translation of proteins through its interaction with RNA helicase eIF4A and scaffold protein eIF4G (52, 53). Thus, PDCD4 plays an important role in carcinogenesis by regulating various proteins involved in cell transformation, cell cycle, tumor growth, and invasion (20). Maspin, a member of the serine protease inhibitor (serpin) superfamily, inhibits cell invasion and metastasis in breast and prostate cancer cells (39, 60). In prostate cancer, maspin can suppress prostate tumor metastases by inhibiting urokinase-type plasminogen activator (uPA)/uPA receptor axis and angiogenesis (9). Additional tumor-suppressive effects of maspin in prostate cancer include increased cell-extracellular matrix (ECM) interaction by promoting cell interaction with ECM components fibronection and laminin (1), and increased apoptotic sensitivity of cancer cells (25, 46). Maspin has also been shown to affect the redox signaling in prostate cancer (55). These authors showed that maspin interacted with glutathione S-transferase (GST) under basal and oxidative stress conditions in prostate cancer cells. This interaction was boosted by more oxidative stress, which, in turn, also led to protection of GST. Furthermore, maspin/GST interaction reduced ROS generation along with lower levels of vascular endothelial growth factor A (55).
In summary, we demonstrate that increased expression of miR-21 in prostate cancer cells is dependent on the high ROS produced by NADPH oxidase. Activation of Akt appears important for the induction of miR-21. Since high levels of ROS are experienced in the ischemic tumor micro-environment, the induction of miR-21 could provide a mechanism by which the tumor cells could escape to a region that is more conducive for survival. The data are corroborated by findings in human prostate cancer tissues and demonstrate high expression of both miR-21 and p47phox subunit of NADPH oxidase. Finally, these data provide evidence that ROS and miR-21 are viable targets for drug development that is aimed at treating metastatic prostate cancer.
Materials and Methods
Materials
DPI, NAC, C-PTIO, and LY294002 were obtained from Sigma-Aldrich (St. Louis, MO). Antibodies against p47phox, maspin, and t-Akt were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA); while those against β-actin, PDCD4, and p-Akt were from Sigma Chemical Co., Epitomics, Inc. (Burlingame, CA) and Cell Signaling Technology, Inc. (Danvers, MA), respectively. Cell culture media RPMI 1640 and penicillin/streptomycin were obtained from Invitrogen (Carlsbad, CA), while fetal bovine serum (FBS) was from Atlanta Biologicals, Inc. (Lawrenceville, GA). Human prostate cancer TissueScan qPCR panels with the clinical information, including the disease stages, were obtained from Origene Technologies (Rockville, MD), and FFPE sections for human prostate cancer were purchased from US Biomax, Inc. (Rockville, MD).
Cell cultures
The androgen-receptor negative and highly metastatic human prostate cancer cell line PC-3M-MM2 was obtained from Dr. Kounosuke Watabe (SIU School of Medicine, Springfield, IL), while DU145 and LNCaP were kindly provided by Dr. Daotai Nie (SIU School of Medicine, Springfield, IL). All these cells were cultured in RPMI 1640 media supplemented with 10% FBS, 50 units/ml penicillin, and 25 μg/ml streptomycin. Cells were grown at 37°C in the presence of 5% CO2 and 95% ambient air. The medium was changed every second day, and the cells were passaged twice a week. The immortalized prostate epithelial cells, RWPE-1, were purchased from ATCC (Manassas, VA), and cultured in keratinocyte serum-free medium (K-SFM) with 0.05 mg/ml bovine pituitary extract (BPE) and 5 ng/ml human recombinant epidermal growth factor (EGF) added to it to make a complete medium. BPE and EGF were supplied with K-SFM as a part of the K-SFM kit by Life Technologies (Grand Island, NY). All the experiments were performed using confluent monolayers.
Transfection of PC-3M-MM2 cells
Transfections of PC-3M-MM2 cells with siRNA and anti-miRs were performed by using RNAifectin from Applied Biological Materials, Inc. (ABM, Inc., Richmond, Canada) according to the manufacturer's protocol. Briefly, 2×105 PC-3M-MM2 cells were plated in 12-well cell culture plates. The next day, solution A was prepared with different concentrations (0–50 nM) of anti-miR-21, si-p47phox, or si-p22phox added to 125 μl SFM. Solution B was made by adding 6 μl of RNAifectin transfection reagent (ABM, Inc) in 125 μl SFM. Both the solutions were then mixed and incubated at room temperature for 40 min to allow the formation of transfection complexes. For control groups, 5 nM of anti-miR-scr and si-scr were used. After 40 min, complexes were diluted with 750 μl media containing serum and added to the cell cultures by gently swirling the plates. Cells were incubated for another 24 h, and later used for Western blotting and real-time RT-PCR studies. For in vitro invasion chamber assay, modified wound-healing assay, and in vivo tumor inoculations, the same protocol was used but with 50 nM of anti-miR-scr and anti-miR-21 or 25 nM of si-scr and si-p47phox.
In vivo studies
To determine the role of miR-21 and p47phox in the spontaneous metastatic ability of PC-3M-MM2 cells, cells were transfected with 50 nM anti-miR-scr or anti-miR-21, or 25 nM si-scr or si-p47phox for 24 h. After that, cells were harvested, and 0.5–1×106 cells in 0.2 ml of 1×PBS were injected intravenously via the tail vein of 5-week-old SCID mice with an average weight of 20 g (Harlan Sprague-Dawley, Indianapolis, IN). Mice were sacrificed 4–5 weeks after the injection of the cells, and metastatic lesions on the lungs were counted macroscopically. All the animal studies were performed in compliance with the SIU School of Medicine's Laboratory Animal Care and Use Committee-approved protocols.
In vitro invasion chamber assay
The ability of prostate cancer cells to migrate through matrigel-coated membranes was measured using BD Biocoat Matrigel invasion chambers (BD Biosciences, San Jose, CA). PC-3M-MM2 cells were transiently tranfected with anti-miR-scr or anti-miR-21, or si-scr or si-p47phox (as described above) for 24 h. These cells were then suspended in the culture media without serum and plated on the top compartment of the invasion chamber followed by respective treatments. At the end of 24 h, the cell inserts were removed, and cells were carefully wiped from the top surface of the membrane with a cotton swab. The invasive cells adhering to the bottom surface of the membrane were stained with toluidine blue and quantified under a light microscope using a 20×objective. Total number of invaded cells was counted in four randomly chosen fields per treatment per insert. Each treatment group was in triplicate.
Modified monolayer scratch assay
After transient transfections of the PC-3M-MM2 cells with anti-miRs and siRNAs, an equal number of cells (2×105) was plated in each well of 12-well culture plates. After the cells reached 70%–80% confluency, a wound was created with a 1000 μl pipette tip. The differential interference contrast (DIC) images were recorded before and just after the wound (time 0 h). The media was changed, and the cells were allowed to grow for another 24 h. DIC images were again captured by confocal microscopy, and results were quantitated by calculating the percentage of wound closure in the treatment groups where the scramble treatment group was assumed to be 100%.
Western blotting
Total cell lysates were used for western blotting studies for PDCD4, maspin, p47phox, and p-Akt. The cells were homogenized in ice-cold 50 mM Tris HCl, 10 mM MgCl2, and 1 mM EDTA in the presence of protease inhibitors cocktail (Sigma-Aldrich), and these lysates were subjected to sodium dodecyl sulfate-plyacrylamide gel electrophoresis. Proteins were then transferred to nitrocellulose membranes, which were developed as previously described (17). The blots were visualized by LAS 4000 chemiluminescence imaging system (GE Healthcare Biosciences, Pittsburgh, PA) using ECL reagent (Pierce Biotechnology, Rockford, IL). The bands were normalized to β-actin, and quantitative analysis was performed with Multi Gauge software (version 3.0) from Fujifilm.
RNA isolation, reverse transcription, and real-time reverse transcriptase polymerase chain reaction
To measure the p47phox and p22phox transcripts the RNA isolation, reverse transcription and RT-PCR was performed as described earlier (17).
PCR primers
The set of primer sequences were from Sigma Genosys (St. Louis, MO):.
p47phox: 5′-CATGTTCCTGGTTAAGTGGCA-3′ (sense);
5′-AGGTGTGGGCAGCGGGAGATC-3′ (antisense).
p22phox: 5′-ACCACGGCGGTCATGTACTT-3′ (sense);
5′-CGCTTCACCCAGTGGTACTT-3′ (antisense).
β-actin: 5′-TGAGACCTTCAACACCCCAGCCATG-3′ (sense);
5′-CGTAGATGGGCACAGTGTGGGTG-3′ (antisense).
TaqMan® Real-Time PCR
Total RNA was isolated from PC-3M-MM2 cells using QIAzol Lysis Reagent (Qiagen, Valencia, CA) as directed by the manufacturer. miR-21 cDNA was generated from 200 ng of total RNA, which was reverse transcribed using hsa-miR-21 qRT-PCR primer set (Applied Biosystems, Foster City, CA) and TaqMan MicroRNA Reverse Transcription (RT) kit (Applied Biosystems). Each RT reaction contained 1×RT specific primer, 1×RT reaction buffer, 0.15 μl of 100 mM dNTPs, 50 U/μl MultiScribe RT enzyme, and 3.8 U/μl RNase inhibitor. The 15 μl reaction mix was then incubated for 30 min at 16°C, 30 min at 42°C, and 5 min at 85°C and then held at 4°C in a PCR cycler. The real-time PCR was performed on Applied Biosystems StepOnePlus™ Real-Time PCR System using a standard TaqMan PCR kit (Applied Biosystems) protocol. Briefly, after the RT step, 2 μl of the RT reaction was combined with 1 μl of 20×TaqMan MicroRNA Assay (forward primer, reverse primer and probe) and 17 μl of TaqMan Universal PCR Master Mix in a 20 μl final volume. The reactions were incubated at 95°C for 10 min, followed by 40 cycles of 95°C for 15 s and 60°C for 1 min. Mature miR-21 expression was calculated using the 2−ddC t method relative to U6-snRNA and expressed as fold change. All TaqMan-PCRs were performed in triplicate.
Immunohistochemistry
FFPE human prostate tumor sections on glass slides were obtained from US Biomax. Immunohistochemistry staining was performed using DAB solution for the detection of PDCD4 and maspin. Antigen retrieval was performed by deparaffinizing, rehydrating, and immersing the tumor section containing glass slides in 10 mM sodium citrate buffer (pH 6.0) for 15 min in 90°C water bath. After cooling down to room temperature, the slides were dipped in 3% H2O2 for 15 min at room temperature to block endogenous peroxidase activity. The sections were then washed twice with 1×PBS and blocked with 5% goat serum in 1×PBS for 20 min at room temperature. The sections were then incubated with respective primary antibodies (1:200 dilution) for 12 h at 4°C after which they were incubated with horseshoe radish peroxidase tagged secondary antibody (Vectastain ABC kit, Burlingame, CA) for 45 min at room temperature according to the manufacturer's protocol. The peroxidase activity was identified by a reaction with DAB solution kit (Vector Laboratories, Inc., Burlingame, CA) for 5 min at room temperature. Sections were then counterstained with hematoxylin, dehydrated with ethanol, and, finally, mounted in Cytoseal 60 (Richard-Allan Scientific, Kalamazoo, MI). Slides were imaged using an Olympus light microscope (Olympus imaging America Inc., Center Valley, PA) using Olympus DP controller software Ver. 2.3.1.231.
Plasmid and siRNA/anti-miRs co-transfection
PC-3M-MM2 cells were simultaneously transfected with plasmid DNA (PDCD4, Maspin, and pGL3 Renilla, a kind gift from Dr. Y.Y Mo, SIU School of Medicine) using DNAfectin and siRNAs (scramble or p47phox) or anti-miRs (scramble or miR-21) using RNAifectin. Briefly, for each well of a 24-well plate, the following mixes were made and added together. (i) Plasmid DNA mix: Solution A was prepared by adding 0.4 μg maspin or PDCD4 luciferase plasmid and 0.1 μg of pGL3 Renilla luciferase plasmid in 32 μl of SFM. In a separate tube, solution B was made by adding 2 μl of DNAfectin in 32 μl of SFM. Both the solutions were combined (about 65 μl total for each well), gently mixed, and incubated for 20 min. (ii) Anti-miR (or siRNA) mix: 50 nM of anti-miR-scr or anti-miR-21 were added to 32 μl of SFM to make solution A, while solution B was prepared by adding 2 μl of RNAifectin to 32 μl of SFM. Both the solutions were combined (about 65 μl total for each well), mixed, and incubated for 20 min at room temperature. At the end of 20 min, 65 μl of the plasmid mix and 65 μl of anti-miR (or siRNA) mix were added to each well containing 130 μl of SFM. SFM was replaced with serum-enriched media after 8 h, cells were incubated for another 24 h, and lysates were used for luciferase assays. For siRNA transfections, a similar protocol was followed except that the concentration used for si-scr and si-p47phox was of 25 nM each.
Luciferase assay
PC-3M-MM2 cells transfected with plasmid and anti-miRs (or siRNAs) were used for this experiment. Transfections were done as described earlier, and luciferase activity was assessed by Dual-Luciferase Reporter Assay kit (Promega, Madison, WI) according to the manufacturer's protocol. Briefly, the cells were harvested using the lysis buffer provided in the kit. 25 μl of the lysate was mixed with 25 μl of luciferase assay substrate, and luciferase activity was measured using a Sirius single-tube luminometer (Titertek Instruments, Inc., Huntsville, AL). To measure the activity of the control Renilla luciferase, 25 μl of stop and glo substrate was then added to the lysate mix, and luminescence was again measured. Renilla luciferase was used for normalization.
H2DCFDA assay
PC-3M-MM2 cells were cultured on glass coverslips in 6-well culture plates. 24 h later, cells were transfected with either si-scr or si-p47phox for 24 h. Cells were then loaded with 5 μg/ml of H2DCFDA (EMD Biosciences, San Diego, CA) in 2 ml of 1×PBS. After incubation at 37°C for 20 min, cells were washed with 1×PBS. Images were recorded by a confocal microscope (Olympus America, Inc., Melville, NY) at 488 nm. For DPI treatment group, cells were pre-treated with vehicle or DPI (5 μM) for 30 min before adding the dye.
Protein determination
The levels of protein in samples were determined by Bio-Rad protein detection reagent, using bovine serum albumin to prepare standard curves.
Statistical analyses
Statistical analyses were performed by using analysis of variance followed by Tukey's honestly significant difference post hoc test to compensate for multiple pairwise comparisons or by paired Student's t-test.
Footnotes
Acknowledgments
The authors would like to thank Drs. Kounosuke Watabe, Yin Yuan Mo, and Daotai Nie (Southern Illinois University School of Medicine) for kindly providing PC-3M-MM2 cells, plasmids for luciferase assays, and LNCap and DU145 cells, respectively. This research was supported by NIH grant (R15 CA135494-01) and an EAM grant from the Southern Illinois University School of Medicine.
Author Disclosure Statement
The authors declare that they have no conflicts of interest.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.