
Abstract
Trypanosomatids and Their Unique Thiol Redox Metabolism
Glutaredoxins Form a Large and Diverse Protein Family
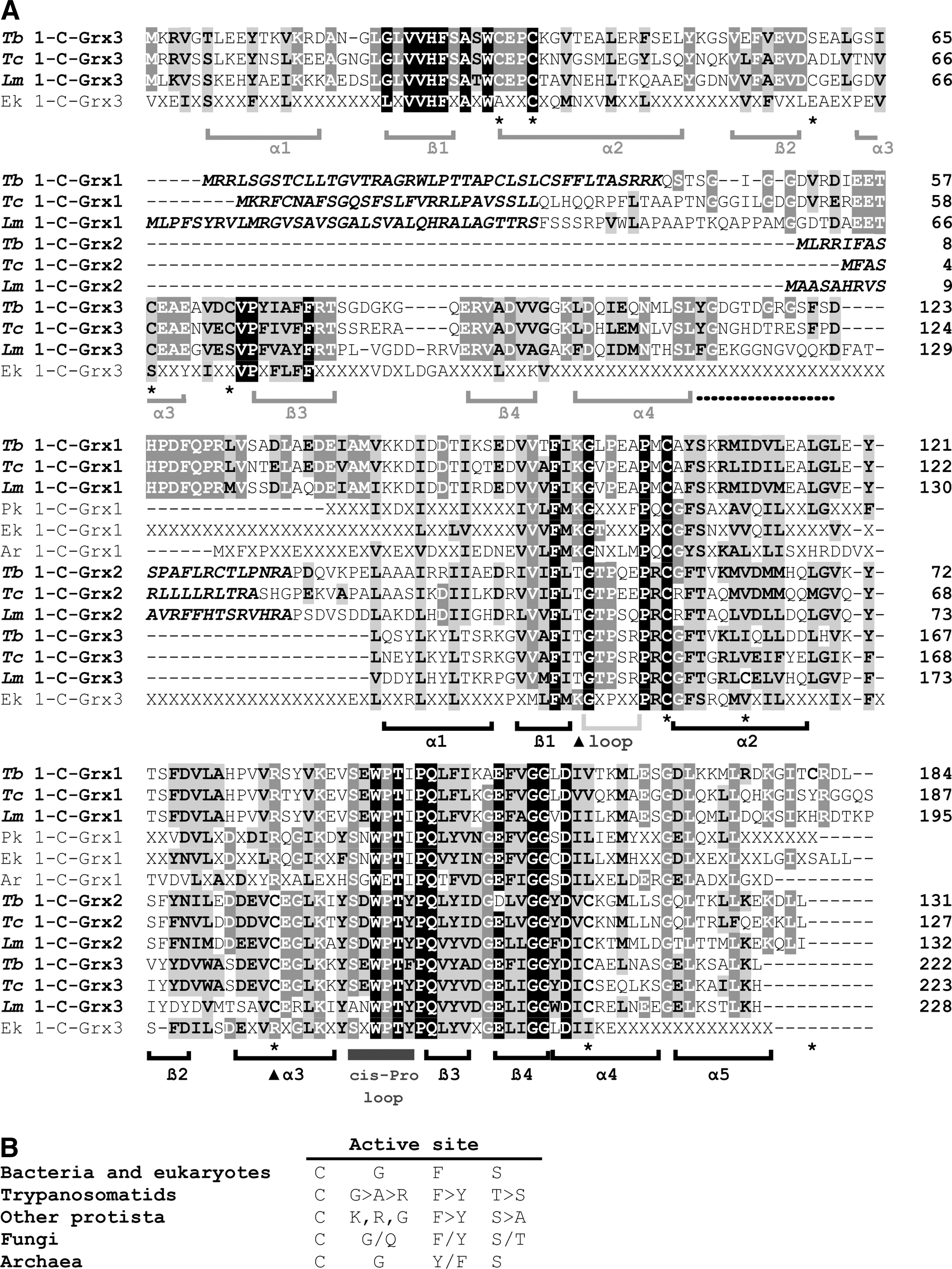
Glutaredoxins are ubiquitous small thiol proteins of the Trx-fold superfamily (68). According to the number of cysteines in the active site, glutaredoxins were initially classified as dithiolic (CXXC motif; 2-C-Grx) and monothiolic (CXXS motif; 1-C-Grx) proteins. However, a recent and thorough phylogenetic analysis identified six distinct classes of glutaredoxins, where the first two are the most widespread ones, present in most prokaryotic and eukaryotic organisms (74). Class I contains CPYC-type 2-C-Grxs and both mono- and dithiol closely related variants with a CPYS, CGYC, CPFC, or CSY[C/S] motif. Class II proteins are 1-C-Grxs with a conserved CGFS active site and can be subdivided into two groups: proteins with a single 1-C-Grx domain and those composed of an N-terminal Trx-like domain that is followed by one or more 1-C-Grx domains (33; 74; Fig. 2). The latter proteins occur almost exclusively in eukaryotes and represent a distinct functional group (35).
The Grx system was first described in the 1980s as “thioltransferase” (3) and as the donor of reducing equivalents for ribonucleotide reductase in an Escherichia coli strain lacking Trx 1 (33). Grxs are usually monomeric proteins that occur in different subcellular compartments (51, 53, 65). There they act as thiol/disulfide oxidoreductases catalyzing the (de)glutathionylation of proteins as well as the reduction of dehydroascorbate and protein disulfides (5, 76). Novel roles have recently emerged for some 2-C-Grxs such as human Grx2 (42, 46), poplar GrxC1 (25), and T. brucei 2-C-Grx1 (12) (see below) based on their capability to bind iron–sulfur clusters (ISCs).
1-C-Grxs were first identified in yeast and subsequently in different organisms (reviewed in 33). With few exceptions, such as yeast Grx 6 (54, 59) and trypanosomal 1-C-Grx1 (16, 27, Fleitas, Manta, and Comini, unpublished), 1-C-Grxs are monomeric proteins in the apo-state (13, 18, 25, 28, 47, 59, 69, 82). Similarly to 2-C-Grxs, distinct isoforms of 1-C-Grxs are found in the cytosol, nucleus, mitochondria as well as plant chloroplast (reviewed in 86). Except for yeast Grx6 and Grx7 (59, 60), most 1-C-Grxs lack or have negligible classical disulfide reductase activity (18, 26, 27, 43, 69, 82). Instead, the capability to coordinate ISCs appears to be a common feature for 1-C-Grxs (16, 40, 42, 43, 54, 67, 89) with yeast Grx7 being, so far, the only known exception (59, 60). The latter feature determines an evolutionary conserved and indispensable role of 1-C-Grxs in the biogenesis and assembly of iron–sulfur proteins (11, 61) and other cell-specific regulatory functions such as the (in)activation of nuclear transcription factors (35, 62, 86).
In trypanosomatids, five Grxs have been identified which are members of class I (T. brucei 2-C-Grx1 and 2) and II (T. brucei 1-C-Grx1, 2 and 3) proteins. The parasite Grxs have a variety of distinctive biochemical, structural and biological features, and are functionally linked to the trypanothione system (12, 16, 27, 56) (Table 1). These proteins are subject of this review.
Data taken from references a 12; b 15, 53; c 29, 70, 78, 79; d 16, 27, 55a, 58. eND, not determined, fsuggested localization based on the absence of a putative targeting signal, g in the mammalian bloodstream form. The light gray shading refers to biophysical and structural properties. The dark gray shading denotes biochemical and biological functions of the proteins.
Trypanosomatids Possess Two Distantly Related Dithiol Glutaredoxins
The genome sequences of trypanosomes revealed two single copy genes for 2-C-Grxs. One gene encodes a protein with an overall sequence identity of 40% compared to human Grx2c but with the active site motif (CPYC) of human Grx1. Consequently, the protein has been named 2-C-Grx1. The protein encoded by the second gene in T. brucei, 2-C-Grx2, possesses an unusual CQFC motif and shares only 25% of its residues with 2-C-Grx1. The 2-C-Grx2 sequences are highly conserved in trypanosomatids, while counterparts in organisms outside the order of Kinetoplastida have not been found. Remarkably, despite an overall sequence identity of 80% among the 2-C-Grx2-type proteins from different trypanosomatids, the Leishmania representatives have a serine residue instead of the second active site cysteine and thus are, by definition, 1-C-Grxs. Alignment of the parasite proteins with structurally characterized 2-C-Grxs from other organisms revealed that residues reported to form a binding groove for GSH on the protein surface are conserved (12). However, a Lys that precedes the active site motif and other charged or polar residues found to interact with the glycine carboxylate of GSH are replaced. These substitutions may support the interaction of the parasite 2-C-Grxs with T(SH)2 where the glycine does not carry a negative charge but forms an amide bond with the spermidine. However, it should be stressed that in E. coli Grx2, which has the highest HED reductase activity of the three bacterial dithiol Grxs (E. coli 2-C-Grx 1 to 3), the Lys is replaced by a Tyr. This supports previous reports that the characteristic γ-Glu-Cys moiety is more important for GSH recognition than the glycine carboxylate (23, 81). As will be outlined in this review, the trypanosomal 2-C-Grxs are involved in reactions with both GSH and/or T(SH)2. Thus, amino acid exchanges that would result in a mutually exclusive specificity for T(SH)2 or GSH cannot be expected.
The role of both 2-C-Grxs has been studied in African trypanosomes (12), and recombinant T. cruzi 2-C-Grx2 has been partially characterized (56). Neither 2-C-Grx1 nor 2-C-Grx2 of T. brucei possesses an obvious targeting signal. Immuno-fluorescence analyses revealed 2-C-Grx1 mainly in the cytosol in both the mammalian bloodstream form and the procyclic insect form of T. brucei (12). In contrast, 2-C-Grx2 occurs in the single mitochondrion of the parasite. Upon fractionated digitonin lysis of procyclic parasites, 2-C-Grx2 co-eluted with cytochrome c, suggesting its presence in the intermembrane space. This localization would correspond to that described for a subfraction of Grx1 in rat hearts (65).
Interplay between the 2-C-Grxs and the trypanothione system
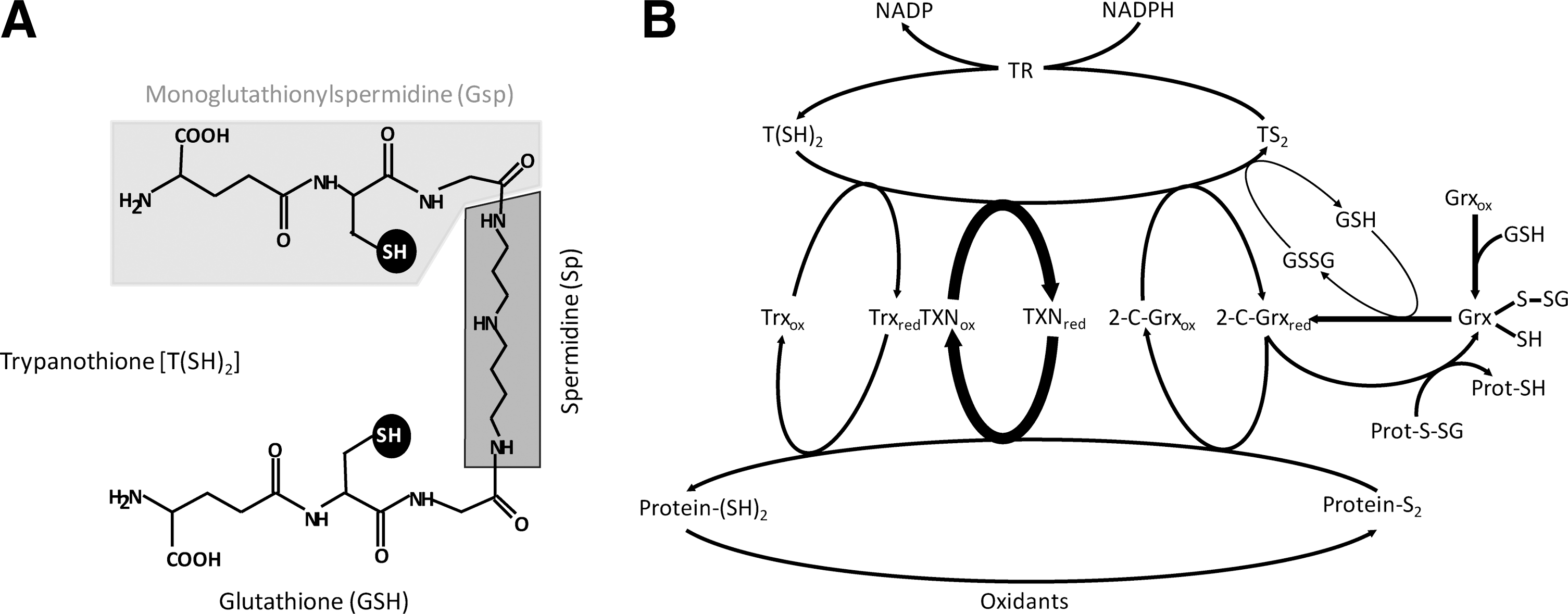
The mode of action of 2-C-Grxs is generally closely linked to the glutathione system. The dithiol form of 2-C-Grxs is regenerated by the spontaneous reaction of the intramolecular disulfide of the protein with two molecules of GSH, and the resulting GSSG is then reduced by NADPH and glutathione reductase (GR). Trypanosomatids lack GR, while containing about 300 μM of each free GSH and T(SH)2 (for reviews see 44, 45). Both thiols spontaneously reduce the parasite 2-C-Grxs. However, the apparent second order rate constants differ significantly, being 100 M −1s−1 and 105 M −1sec−1 for the reaction of GSH and T(SH)2, respectively (12). Thus, T(SH)2 is about 1000-fold more efficient compared to GSH and should be the main physiological reducing agent of the parasite 2-C-Grxs.
In trypanosomatids, GSH is regenerated from GSSG by direct thiol/disulfide exchange with T(SH)2. However, this spontaneous reaction is probably not sufficient at lowered T(SH)2 concentrations as they may occur under severe oxidative stress conditions. To mimic this situation, the reaction of 9 μM T(SH)2 with 20 μM GSSG—conditions at which the spontaneous reaction is negligible—was measured. 2-C-Grx1 and 2-C-Grx2 catalyzed the reaction with 106 U/mg and 1.8 U/mg, respectively (12), showing that 2-C-Grx1 is significantly more efficient compared to 2-C-Grx2 both from T. brucei (12) as well as T. cruzi (56). Thus, in trypanosomes the cytosolic 2-C-Grx1 may substitute for GR providing kinetic control for keeping GSH in the reduced state (Fig. 1B).
Oxidoreductase activity of the parasite 2-C-Grxs
The T. brucei 2-C-Grxs reduce the mixed disulfide between GSH and either 2-mercaptoethanol (HED assay) or cysteine residues of bovine serum albumin (BSA) and T. brucei peroxiredoxin. The kcat/Km values towards the glutathionylated model proteins were about 1×105 M −1 s−1 and 1×104 M −1 s−1, respectively, values very similar to those reported for human Grx1 and Grx2 (30, 41). In contrast, T. brucei TXN catalyzes the deglutathionylation of BSA-SSG with a much lower catalytic efficiency of only 4×102 M −1 s−1. Thus, protein deglutathionylation may indeed be a specific in vivo role of the parasite 2-C-Grxs (Table 1).
The trypanosomal 2-C-Grxs also catalyze the reduction of protein disulfides such as insulin disulfide, used as in vitro model substrate. Although this reaction was first shown to be catalyzed by Trxs (37), many Grxs also act as protein disulfide reductase, although mostly with lower efficiency. With T(SH)2 as reducing agent, E. coli Grx1 (used as a control) displayed the highest activity, followed by T. brucei 2-C-Grx2, whereas 2-C-Grx1 had only low activity (12). When comparing the reduction of insulin disulfide by DTE and T(SH)2, both parasite proteins had the highest activity with T(SH)2, whereas E. coli Grx1 exerted identical activity with both dithiols. The superior reducing capacity of T(SH)2 towards the parasite Grxs suggests some specific interactions. However, the fact that E. coli Grx1 equally well accepted DTE and T(SH)2 strongly suggests that individual glutaredoxins do not display a strict thiol specificity. Taken together, the parasite 2-C-Grxs may act as physiological reductases for both protein disulfides and protein-glutathione mixed disulfides (Figs. 1b and 3). In the case of the 2-C-Grx2—especially in the insect stage of the parasite with its fully elaborated mitochondrion—these types of substrates may originate from reactive oxygen species leaking out from the respiratory chain (Fig. 3). In this respect, mammalian Grx1 and Grx2 have been suggested to regulate the local thiol redox homeostasis in the intermembrane space and matrix, respectively (65).
As described above, in contrast to the 2-C-Grx2 proteins from T. brucei and T. cruzi, the Leishmania orthologues display a serine instead of the second active site cysteine. One may speculate that the proteins have distinct functions in the individual parasites, but this is not very likely. A serine replacing the second cysteine may also stabilize the thiolate of the nucleophilic first cysteine. By precluding formation of an intramolecular disulfide, this substitution may even result in a protein species with higher activity in reactions that follow a monothiol mechanism such as protein deglutathionylation (19, 30, 41). The precise role of the second active site cysteine residue in the catalytic mechanism of 2-C-Grxs is, however, still an open question. Replacement of the cysteine by a serine can result in increased (19, 30, 41) or decreased (19) activities, depending on the individual protein. As shown recently, the GrxC5, localized in chloroplasts of Arabidopsis thaliana, exclusively relies on a monothiol mechanism to reduce insulin disulfide (17), a reaction regarded to follow a dithiol mechanism (9). On the other hand, in yeast Grx8 both active site cysteines are involved in the (low) activity towards glutathione-mixed disulfides and a refined catalytic model for both mono- and dithiol Grxs has been suggested (22).
Functional comparison of the parasite 2-C-Grxs with thioredoxin and tryparedoxin
Besides the two 2-C-Grxs described above, trypanosomatids encode two other types of dithiol oxidoreductases, namely Trx(s) and the parasite-specific TXN(s). The physiological role of Trx in both African trypanosomes (79) and T. cruzi (66) is not yet known. Silencing of the gene by RNA interference in procyclic T. brucei and the deletion of both alleles in infective cells did not affect the proliferation of the parasites, at least under culture conditions (79). TXNs have a molecular mass of about 16,000 Da and are characterized by a CPPC active site motif. They are mainly cytosolic proteins (84). In vitro, TXN transfers the reducing equivalents from T(SH)2 onto ribonucleotide reductase (20), methionine sulfoxide reductase (2), as well as 2-Cys-peroxiredoxins (8, 63) and non-selenium glutathione peroxidase-type enzymes (34, 77). Thus, this parasite-specific oxidoreductase exerts many functions that in other organisms are fulfilled by Trxs and/or Grxs. Recombinant T. brucei 2-C-Grx1 can replace TXN in the transfer of electrons onto ribonucleotide reductase, although the reaction is less efficient. In vitro, TXN is a more potent protein disulfide reductase compared to the 2-C-Grxs, while its activity towards glutathionylated proteins is very low (12). Thus, protein (de)glutathionylation is probably a physiological function of the parasite 2-C-Grxs that is not accomplished by TXN.
Pathogenic Trypanosomes Encode Three Monothiol Glutaredoxins
The genomes of T. brucei, T. cruzi, and L. major (
Expression of the 1-C-Grxs in African trypanosomes
So far, the proteins from T. brucei represent the best studied 1-C-Grxs from a protozoan organism. All three proteins are expressed in both the mammalian infective as well as the insect stage of T. brucei, with the highest levels found in stationary phase and starving parasites (16). The single domain proteins 1-C-Grx1 and 1-C-Grx2 reside in the mitochondrion in both life stages of T. brucei (16, 27, Comini: unpublished data). The subcellular localization of 1-C-Grx3 has not been studied but is likely to be cytosolic due to the absence of recognizable targeting signals. The total cellular concentration of 1-C-Grx1 in logarithmically growing bloodstream T. brucei is about 5 μM (16). This indicates an unusually high mitochondrial concentration of about 200 μM when taking into account that the rudimentary organelle of this parasite form has been reported to account for only 2.3% of the total cell volume (64). The concentration of 1-C-Grx2 is at least two orders of magnitude lower than that of 1-C-Grx1 (16).
Oligomeric state and redox properties of the T. brucei 1-C-Grxs
Recombinant T. brucei 1-C-Grx1 exists as a noncovalent homodimer in both the apo-form and with bound iron–sulfur cluster, whereas the apo-forms of 1-C-Grx2 and 1-C-Grx3 are monomeric proteins (16, 27). An N-terminal region of about ∼37 residues, exclusive of the kinetoplastid 1-C-Grx1s, has recently been identified to be responsible for dimerization of the T. brucei and T. cruzi proteins (55a). In vitro, T. brucei 1-C-Grx1 lacks any general thiol-disulfide oxidoreductase activity (27) but is susceptible to oxidation by GSSG and Gsp disulfide (16, 58). In addition to the active site Cys104, T. brucei 1-C-Grx1 contains a second cysteine, namely Cys181, located at the fourth position from the C-terminus (Fig. 2A). In contrast to the specificity displayed by E. coli Grx4 and S. cerevisiae Grx5, Cys181—not Cys104—forms a mixed disulfide with GSH and Gsp (58), which can trigger the formation of an intramolecular Cys104–Cys181 disulfide bridge (16, 58). In vitro, both oxidation states are reduced by TXN or the T(SH)2/TR system (27, 58). The physiological significance of the oxidized forms of 1-C-Grx1 as well as the potential reducing agent in vivo remains to be elucidated since Cys181 is not conserved. Interestingly, preliminary data indicate that also the active site cysteine in the Grx domain of T. brucei 1-C-Grx2 and 1-C-Grx3 is refractory to thiolation by GSH and Gsp while other cysteine residues get specifically modified (Melchers J. PhD Thesis, Heidelberg University, 2008). Future work should reveal if this is a peculiarity of the trypanosomal 1-C-Grxs and/or may play a regulatory role in protein function.
The Potential Role of Glutaredoxins in the Parasite Iron–Sulfur Metabolism
Features of the iron–sulfur complexes coordinated by 2-C-Grx1 and 1-C-Grxs
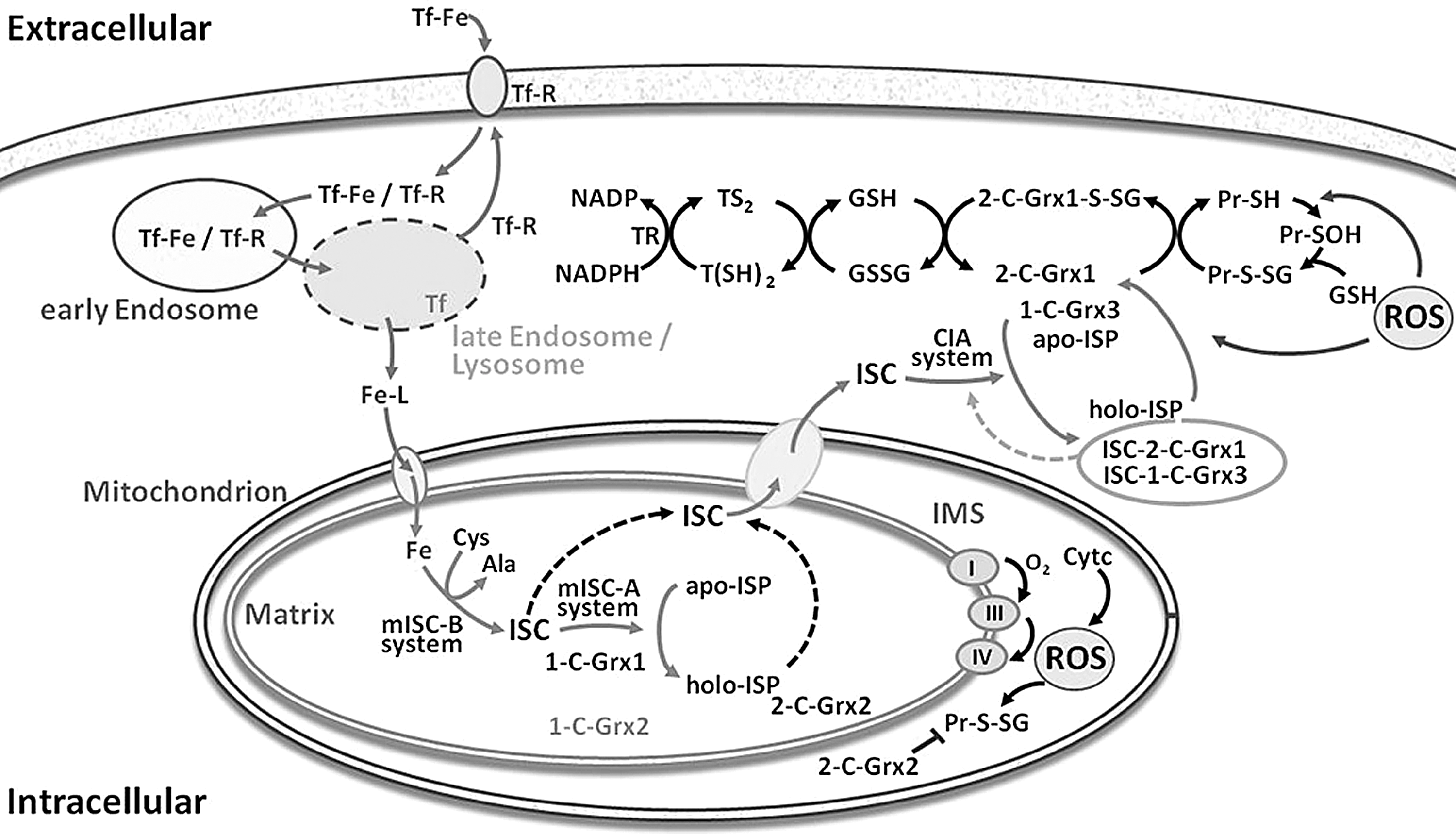
The distinct physiological function(s) of the glutaredoxins in T. brucei are not yet known. Remarkably, in vitro all three 1-C-Grxs as well as 2-C-Grx1 are capable of binding an iron–sulfur complex (ISC). Upon purification from recombinant E. coli, the proteins were obtained as light brown solutions with absorption maxima at 320 nm and 420 nm, in addition to the protein peak at 280 nm (12, 16), indicative of a protein/iron–sulfur complex (6, 75). Size exclusion chromatography of these species revealed that binding of the ISC led to the dimerization of the monomeric apo-forms of 2-C-Grx1, 1-C-Grx2, and 1-C-Grx3 (12, Manta and Comini, unpublished; Fig. 4). In the case of 1-C-Grx1, ISC-binding increased the hydrodynamical radius of the homodimeric protein (16, 55a). Other 1-C-Grxs shown to form dimeric apo-proteins are S. cerevisiae Grx6 and Grx7 (59). Binding of an ISC to yeast Grx6 results in tetramerization of the protein (59). The [2Fe–2S] nature of the ISC has been determined for 1-C-Grx1 (16). As reported for several ISC-glutaredoxin complexes (6, 25, 40, 42, 43, 54, 67, 75), most probably two GSH molecules act as the low molecular mass ligands of the ISC in the holo-form of the trypanosomal proteins extracted from E. coli. In vitro reconstitution of the iron–sulfur proteins in the presence of Gsp or T(SH)2 revealed that these parasite specific thiols can easily substitute for GSH as the nonprotein ligand(s). Strikingly, the dithiol T(SH)2 was capable to ligate the chromophor also to the monomeric form of 2-C-Grx1 which probably represents an intermediate during formation of the holo-protein (12, 55a).

A cysteine in the first position and the absence of a proline in the second position of the active site motif, as well as the noncovalent binding of GSH are conserved features among glutaredoxins capable of coordinating an ISC (6, 25, 40, 42, 43, 54, 67, 75). For cluster coordination, T. brucei 1-C-Grx1 also relies on its active site cysteine but the protein is not definite with respect to the nature of the thiol ligand and the mechanism of ISC binding (55a). T. brucei 2-C-Grx1 was the first authentic 2-C-Grx with the canonical CPYC active site shown to incorporate an ISC (12). These data suggest that additional molecular features determine the productive binding of an ISC to the trypanosomal Grxs. The protein–ISC complexes formed by T. brucei 2-C-Grx1 and 1-C-Grx1 are labile to air and/or H2O2 exposure (12, 16). The yield of holo-complexes obtained upon in vitro reconstitution is dependent on the low molecular mass thiol coordinated, being highest with T(SH)2 followed by Gsp and finally GSH (12, 55a). This may suggest that in the parasite T(SH)2 serves as the ligand resulting in a comparably stable ISC. Interestingly, T(SH)2, but not the monothiols GSH and Gsp, can form a protein-free iron-containing complex which subsequently can be incorporated into 2-C-Grx1 (12, 55a).
Glutaredoxins and regulation of the iron–sulfur metabolism
The mitochondria of eukaryotic cells are responsible for the de novo synthesis of ISCs that are either assembled to iron–sulfur proteins or transferred to the cytosol for further incorporation into target proteins via a specialized assembly system (48). This system is operative in African trypanosomes and the depletion of a cysteine desulfurase and a primary ISC-scaffold protein has been shown to be deleterious for the insect stage of the parasite (80). 1-C-Grxs have been initially recognized as key components of the mitochondrial machinery probably being involved in the chaperone-assisted transfer of ISCs to acceptor proteins (Fig. 3) (11, 73, 87, 88). Complementation studies using a yeast mutant defective in mitochondrial Grx5, which is unable to maturate iron–sulfur proteins and accumulates toxic levels of intracellular iron, demonstrated the evolutionary conserved function for 1-C-Grxs from different eukaryotic and prokaryotic organisms (11, 13, 61, 87). Intriguingly, respective experiments with the three T. brucei 1-C-Grxs revealed that only 1-C-Grx1 modestly rescued the mutant phenotype (27). The striking lack of functional conservation is indicative of a significant structural and/or functional divergence of the parasite proteins. Any genetic manipulation of T. brucei aimed at abrogating or downregulating 1-C-Grx1 failed, which strongly suggests an essential role of the protein in the mitochondrion of the parasite and the incapacity of 1-C-Grx2 to take over this function (16). A regulatory function for 1-C-Grx1 in the mitochondrial iron/redox metabolism can also be envisaged. Bloodstream T. brucei takes up iron via a transferrin receptor (Fig. 3). Cells overexpressing (>10-fold) an ectopic copy of 1-C-Grx1 are more affected in vitro upon iron deprivation as well as oxidative stress and cannot sustain infection in the mammalian host when compared to wild-type parasites (16, 55a).
In yeast, the cytosolic Trx/1-C-Grx hybrids Grx3 and Grx4 are involved in intracellular iron sensing and trafficking. The mechanism includes the ISC-mediated interaction of Grx3 and Grx4 with transcription factors that, depending on the iron levels, regulate operons controlling iron usage or uptake (35, 62). It seems unlikely that the parasite 1-C-Grx3 plays a similar role, owing to the fact that transcription factors are almost absent in trypanosomes and gene expression is mainly controlled at post-transcriptional level (14). However, 1-C-Grx3 and 2-C-Grx1 may indeed play a role in the cytosolic ISC assembly machinery, for example, as ISC scaffold protein and/or by mediating ISC transfer to apo-proteins (Fig. 3). Alternatively, as proposed for the mitochondrial human Grx2 (49), the ISC formation may serve as a redox sensor for the regulation of 1-C-Grx3 and 2-C-Grx1. Under conditions with lowered free thiol levels, the complex may be broken down releasing the catalytically active monomeric protein species.
Structural Basis for Monothiol Glutaredoxin Function
The Trx fold, a four-stranded ß-sheet flanked by three α-helices, is amongst the most conserved structural motifs found in proteins that are not necessarily related in function (57; Fig. 5). Modifications of this core fold lead to remarkable structural diversity that allows for classification of its members into eleven distinct evolutionary families (68). A network-based computational approach, used to create similarity-based maps of the Trx fold class, has demonstrated that Grxs are not a cohesive superfamily and that 1-C-Grxs are distinctly related with other classes within the superfamily (4). Two key signatures distinguish 1-C-Grxs with a CGFS active site (class II) from classical 2-C-Grxs and other 1-C-Grxs (class I, see above). These are an insertion of five residues forming a loop that precedes the active site, as well as a highly conserved WP motif within another loop that contains also the conserved cis-Pro that is part of the putative binding site for GSH (Fig. 2A). Mesecke et al. (59) already suggested that these structural elements may be relevant in determining the redox activity of 1-C-Grxs. Remarkably, 1-C-Grxs that contain these structural elements lack general oxidoreductase activity (e.g., yeast Grx5, E. coli Grx4; Plasmodium falciparum Glp1, T. brucei 1-C-Grx1) (26, 27, 69, 72). In contrast, yeast Grx6, classified as 1-C-Grx, for its CSYS active site, but lacking this loop and the conserved WP motif, maintains the ability to reduce the mixed disulfide of glutathionylated ß-mercaptoethanol (54, 59). Mutational analysis of 1-C-Grxs should reveal whether these structural motifs are sufficient to determine the lack of redox activity in certain protein subgroups.

Structural analysis of a truncated version of T. brucei 1-C-Grx1 as well as 1-C-Grx3
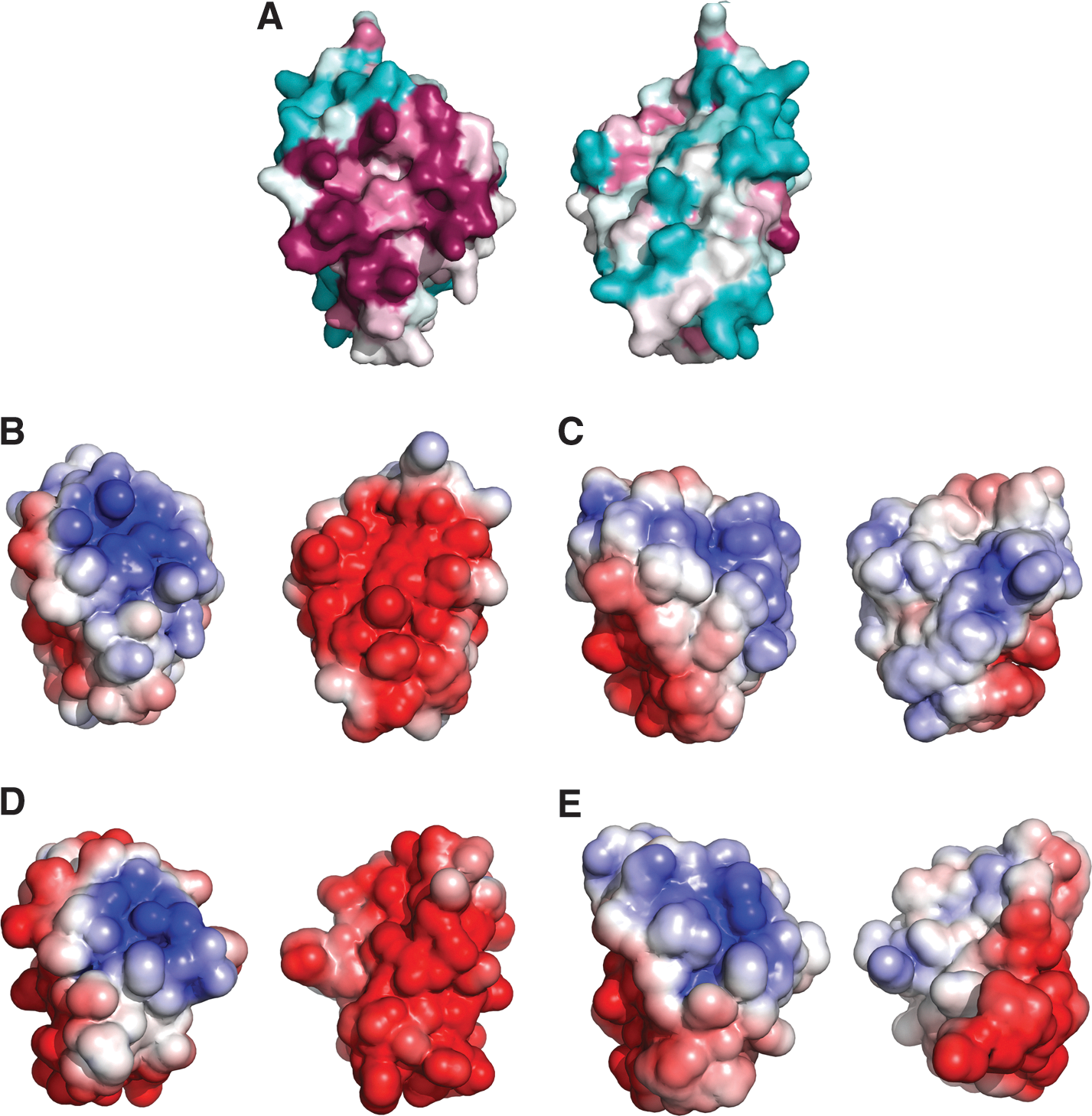
Very recently, we achieved the complete NMR assignment of a truncated version of T. brucei 1-C-Grx1 (residues 76–184, protein lacking the mitochondrial targeting signal and the stretch responsible for dimerization) and the structure was solved (PDB 2LTK; [55a]) based on the semi-automated analysis of NOE-derived interproton distance (32). The overall fold comprises of a four-stranded ß-sheet, flanked by five α-helices (Fig. 5A) and thus is very similar to those of other 1-C-Grxs (Fig. 5B), as well as classical Grxs (not shown). As expected, the active-site Cys104 is exposed to the solvent and the side chain of Lys96 is in proximity of the thiol group, probably contributing to decrease its pKa-value (76a). Most of the residues shown to be involved in GSH binding and ISC assembly in other 1-C-Grxs (e.g., Lys96, Cys104, and Asp 159 of T. brucei 1-C-Grx1) (10, 42, 54, 67) are conserved in T. brucei 1-C-Grx1 (Figs. 2A and 5C) and show a spatial orientation similar to that observed in the dimeric structure of E. coli Grx4 coordinating a 2Fe-2S cluster which presents a noncovalently bound GSH for each monomer (PDB 2WCI; 40; Fig 5C). A remarkable exception is Thr144. The residue is part of a loop whose conformation differs from that of the respective residues in the crystal structures of yeast Grx5 (PDB 3GX8; Wang et al. unpublished) and E. coli Grx4. In the latter structures, the conserved loop, containing the monothiol WPT motif, adopts a very similar conformation stabilized by a network of hydrogen bonds. Intriguingly, T. brucei 1-C-Grx1 is not capable of establishing this H-bond mediated anchoring of the loop, mainly because Phe149 replaces an otherwise conserved tyrosine or tryptophan residue (Tyr108 in yeast Grx5, Trp 75 in E. coli Grx4) that contributes with its side chain to fix the conformation of the structural element. The different conformation of the loop in T. brucei 1-C-Grx1, which significantly distorts the GSH binding pocket, is dictated by steric interactions between Ile145 and Val136. Experimental evidence indicates that this loop presents structural plasticity and we have proposed that the backbone dynamics of this region may modulate the binding of the glutathione moiety and/or influence the catalytic properties of the protein (55a). Interestingly, Ile145 precedes the conserved cis-Pro which has been proposed to be crucial for determining the redox properties of different Trx fold proteins (71). In addition, Tb 1-C-Grx1, as well as other kinetoplastid's 1-C-Grxs, present a 5-residues insertion upstream the active site (see Fig. 2A) that lacks secondary structure (Fig. 5D) and has been proposed to contribute to the almost negligible redox activity of 1-C-Grxs from other organisms (see above; 59).
Analysis of the electrostatic surface of T. brucei 1-C-Grx1 discloses two main areas that are characterized by a concentration of identical charges: a conserved, positively charged surface patch that comprises most of the putative GSH binding residues and a negatively charged region on the opposite side of the molecules (Fig. 6). The latter region of negative electrostatic potential is also observed in yeast Grx5 where it is even more extended (PDB 3GX8). In contrast, in E. coli Grx4 the same region is occupied by a hydrophobic patch (28, 40). The functional role of these regions has yet not been addressed, and one possibility is that they serve as specific protein–protein interaction areas.
In comparison to the other types of parasite 1-C-Grxs, the trypanosomatid 1-C-Grx1s show a 4–10 residues long C-terminal extension. In the case of T. brucei 1-C-Grx1, these residues form a relatively rigid segment (as probed by 15N relaxation experiments), without regular secondary structure. At the fourth position form the C-terminus Cys181 is located (Figs. 2A and 5A) which in vitro can form an intramolecular disulfide with the active-site Cys104 (58). In the available structure the two residues are positioned at a distance far beyond that required for a disulfide bridge, and only a substantial conformational change or a local unfolding would allow formation of this covalent link. Interestingly, a structural rearrangement has also been hypothesized for E. coli Grx4 to explain the reaction between two cysteine residues distantly located in the solved structure (28). The segment comprising residues 180–182 adopts an extended conformation with parallel backbone NH and CO bonds oriented toward the exterior of the protein. Under oxidative conditions, this could promote a transient association of GSSG via antiparallel β-strand backbone hydrogen bonds with subsequent formation of a cross-strand disulfide between the cysteine of GSH and Cys181. Cross-strand disulfides are highly strained (38) and this intermediate complex could in turn trigger the conformational transition required for the formation of the intramolecular disulfide. The conformation of Cys181, together with the distortion of the GSH pocket described above, might explain why this cysteine, and not Cys104, is glutathionylated in Tb 1-C-Grx1.
So far, there is no structural information available for multi-domain 1-C-Grxs. As described above, T. brucei 1-C-Grx3 is a Trx/1-C-Grx hybrid protein for which, due to the high structural conservation of this class of proteins, the backbone conformation of both domains can be easily predicted in silico. The recently achieved backbone assignment for both domains of T. brucei 1-C-Grx3 and the measurement of a set of residual dipolar couplings (RDCs) allowed us to validate the models in term of type and orientation of secondary structure elements (unpublished). The two domains of 1-C-Grx3 are linked by eleven residues (residues 112–122, Fig. 2A), which are not conserved and therefore cannot be modeled. Interestingly, our 15N relaxation measurements and proteolysis experiments (unpublished) show that this linker is not particularly flexible compared with the rest of the protein. Accordingly, RDCs measurements support a defined relative orientation of the two domains. Ongoing studies aim at determining the relative spatial arrangement of the two domains and their interaction interface. This should shed light on the role of the N-terminal Trx domain that at least in yeast has been shown to be essential for the function of multi-domain Trx/1-C-Grxs whose physiological role cannot be replaced by single domain 1-C-Grxs (35).
Summary and Outlook
Trypanosomatids are equipped with two dithiol and three monothiol glutaredoxins. The specialization of the trypanosomal proteins in using T(SH)2 as cofactor for redox reactions and/or ISC binding constitutes a remarkable evolutionary adaptation of Grxs to the unique thiol redox metabolism of these parasites. The ability of 2-C-Grx1 to catalyze the reduction of GSSG by T(SH)2 has probably evolved to keep glutathione in the reduced state–especially under oxidative stress conditions–in organisms that lack a GR. Both 2-C-Grxs reduce protein disulfides as well as GSH-protein mixed disulfides. While the former type of reaction, at least in vitro, is even more efficiently catalyzed by TXN, protein deglutathionylation seems to be a specific function of the parasite 2-C-Grxs.
Interestingly, four of the five T. brucei Grxs can coordinate an ISC in vitro. Putative roles in ISC and/or iron metabolism can be expected for the single domain 1-C-Grxs, considering that this is an almost strictly conserved function for this protein subgroup (33, 74). The precise role of 1-C-Grx1 in the parasite mitochondrion remains unknown. The protein is indispensable for the mammalian infective form and not functionally redundant with 1-C-Grx2. Recent progresses in the biochemical and structural characterization of the parasite 1-C-Grxs have displayed distinct properties such as T(SH)2 as preferred thiol ligand, homodimeric structure, electrostatic surface, and the inferred mechanism for ISC binding. These features are likely responsible for the specificity of the trypanosomal proteins and, hence, their inability to substitute Grx5 in yeast mitochondria. 2-C-Grx1 and probably also 1-C-Grx3 are cytosolic proteins. In vitro, 2-C-Grx1 coordinates an ISC, but the significance of this reaction in vivo is not yet known. As suggested for human Grx2, formation of the complex may convert the protein into a catalytically inactive form unless the level of low molecular mass thiols is dramatically lowered such as under severe oxidative stress conditions. Alternatively, 2-C-Grx1 may play a role in the maturation of cytosolic ISC proteins. Future work, including oxidative challenge, alteration of iron homeostasis, and in vivo studies, on 2-C-Grx1-depleted or -deficient parasites should reveal the physiological functions of the protein. 2-C-Grx2 is the only parasite Grx that does not bind an ISC in vitro. It may play an important role in the mitochondrial redox metabolism, especially of procyclic parasites with their fully elaborated organelle. The identification that the 2-C-Grx2 orthologues from all Leishmania species have a single active site cysteine poses additional challenges for the functional role(s) and classification of (the parasite) glutaredoxins.
Footnotes
Acknowledgments
Financial support by the Access to Research Infrastructures activity in the 7th Framework Programme of the EC (Project number: 261863, Bio-NMR) for conducting the research is gratefully acknowledged by Massimo Bellanda. Marcelo Comini acknowledges support from the Agencia Nacional de Investigación e Innovación (Grant Innova Uruguay, agreement No. DCI–ALA/2007/19.040 between Uruguay and the European Commission).
Author Disclosure Statement
No competing financial interests exist.