
Abstract
Introduction
The Trx Family of Proteins
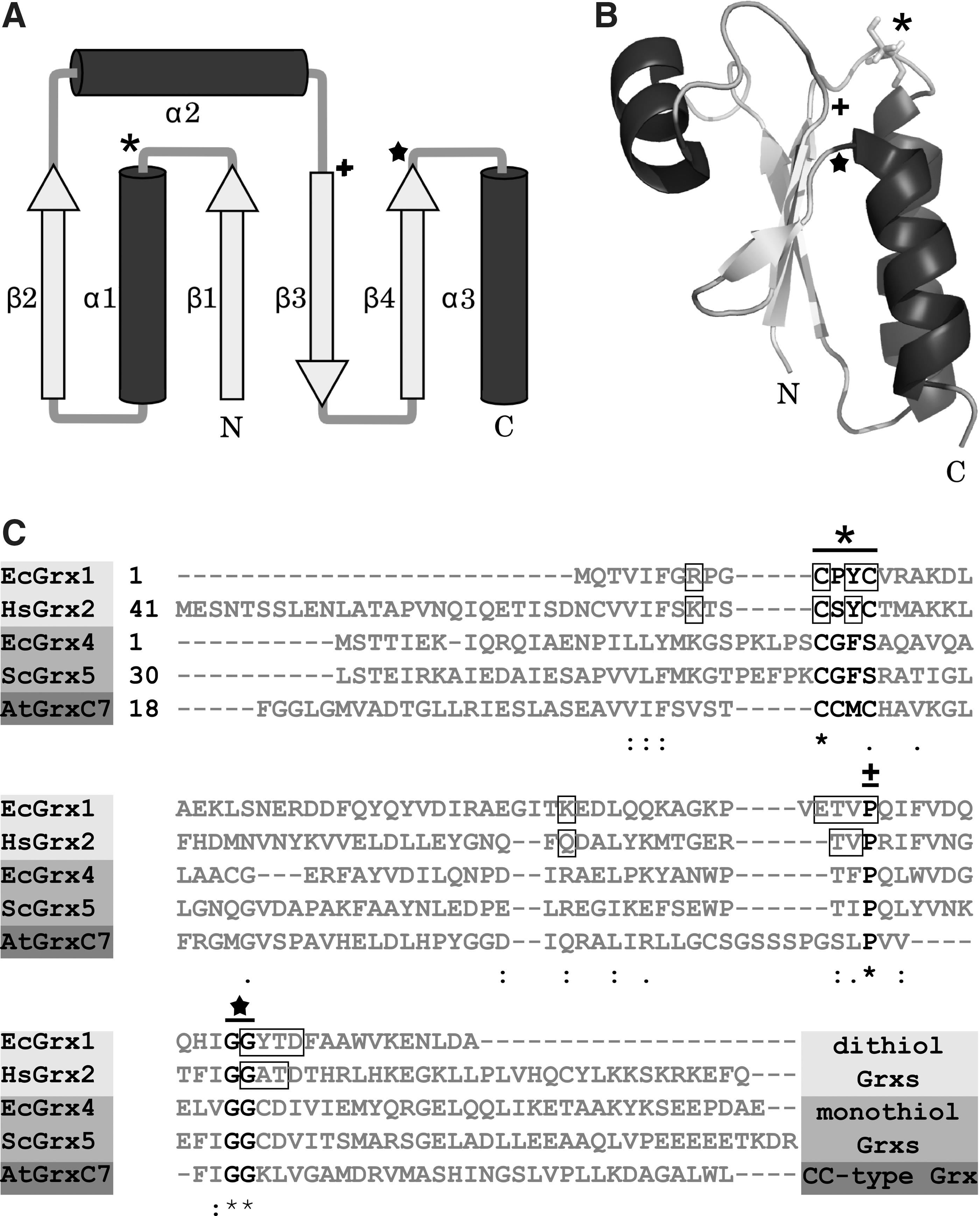
The name giving founding member of the Trx family of proteins was discovered in 1964 as an electron donor for ribonucleotide reductase (RNR) (53). RNR is the enzyme providing deoxyribonucleotides for DNA synthesis and, thereby, essential for proliferation. Today, hundreds of family members have been identified and characterized in all kingdoms of life. Enzymatically, most of them exhibit an oxidoreductase activity, but also proteins with other activities possess the hallmark of the Trx family, the Trx fold. The basic motif of this fold is displayed by Escherichia coli Grx1. It consists of a central four-stranded anti-parallel β-sheet surrounded by three α-helices (Fig. 1) (64). In Trx family members encoded in the genomes of higher organisms, the fold is supplemented by at least one additional β-sheet and one additional α-helix (64). The oxidoreductases of the Trx family are further characterized by their active site motif Cys-x-x-Cys, which is located on the loop connecting β-sheet 1 and α-helix 1, and a cis-proline adjacent to β-sheet 3 and located opposite to the active site (Fig. 1). The cis-proline is conserved in most of the Trx family members, including Grxs, and of suggested importance for substrate binding (64).
Glutaredoxins
In 1976, Grxs were introduced by Arne Holmgren as a glutathione (GSH)-dependent alternative electron donor for the reduction of E. coli RNR next to Trx (37). Today, we know that the Grx system is much more than a back-up system for Trx and impacts on nearly all aspects of life.
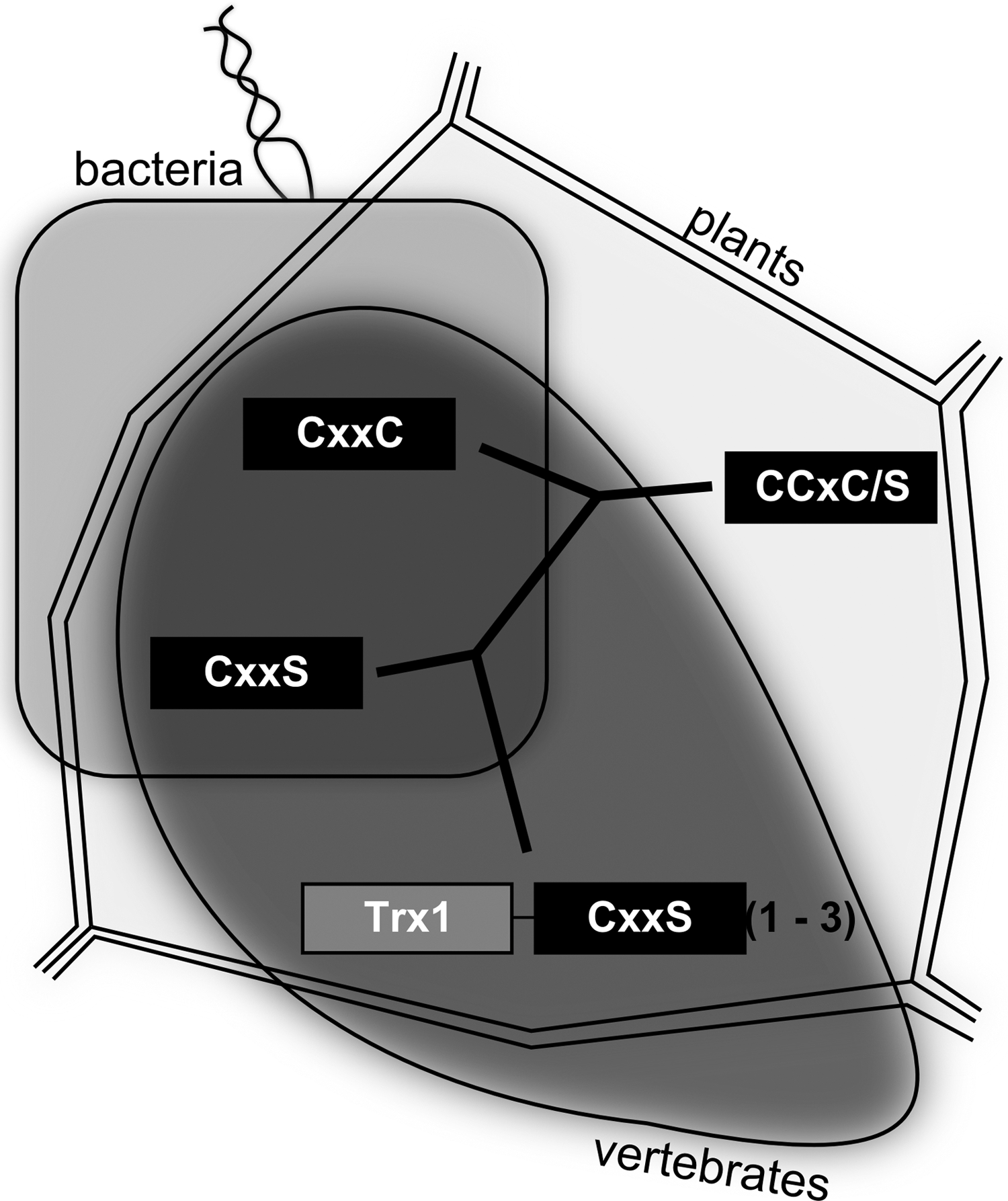
According to their active site motif, Grxs can be divided into dithiol (Cys-x-x-Cys) and monothiol (Cys-x-x-Ser) Grxs (Fig. 1C); the latter can be subdivided in single- and multidomain monothiol Grxs (35, 54). Dithiol and single-domain monothiol Grxs are present in all forms of life, namely, bacteria, plants, and vertebrates, whereas multidomain Grxs were hitherto not identified in bacteria (Fig. 2). An additional division of the Grx family is based on a cysteine residue on position two of the active site motif (Cys-Cys-x-Cys/Ser) (Fig. 1C). These so-called CC-type Grxs are exclusively found in land plants (Fig. 2), where they control plant physiology, for example, flower development (105). Intracellular Grxs are located in the cytosol, the nucleus, the mitochondria, and in yeast as well as in the golgi apparatus (54, 68). Moreover, human Grx1 can be secreted by a leaderless pathway (61). In the different organelles, dithiol Grxs catalyze the reduction of protein disulfides and glutathionylated thiols (see section “Physiological and Pathophysiological Conditions Regulated by Grx Mediated Thiol/Disulfide Exchange”). Thereby, Grxs contribute to local compartmentalized redox signaling. Most of the monothiol Grxs lack these enzymatic activites (35).
Oxidized Grx is reduced by the GSH system consisting of GSH, glutathione reductase (GR), and NADPH (Fig. 3). Some Grxs, such as E. coli Grx4 and human Grx2, can also utilize electrons provided by Trx reductase (48, 25). The enzymatic activity of Grxs depends on their interaction with GSH. Therefore, Grxs exhibit specific GSH binding sites (Fig. 1C). This binding area comprises the active site itself, especially the N-terminal active site cysteine, the residues (consensus sequence: GxdD) following the N-terminal glycine residue of a conserved GG-motif (Fig. 1), and two residues (consensus sequence: TV) before the cis-proline (54). Moreover, two positively charged amino acids located in the N-terminal of the active site and of the cis-proline, respectively, contribute to GSH binding (54). Besides the interaction with GSH during the catalytic cycle, some Grxs utilize noncovalently bound GSH molecules within the same binding area as ligands for the coordination of a [2Fe2S]2+ cluster (7, 47, 86) (see section Iron–sulfur cluster).

Thiol/Disulfide Exchange
The overall reaction of Grx-mediated thiol/disulfide exchange follows a ping-pong mechanism (32) characterized by the successive separate enzymatic reaction with two substrates. Two interconnected mechanisms have been described, the dithiol and the monothiol mechanism. During the dithiol mechanism, Grxs catalyze the reversible reduction of protein disulfides using both active site cysteine residues. The monothiol mechanism utilizes only the N-terminal active site cysteine and reduces mixed disulfides formed between GSH and proteins or other small thiol compounds. All dithiol Grxs investigated so far are able to catalyze monothiol reactions, but not all were tested for their activity in dithiol reactions. However, until now, all dithiol Grxs tested for catalyzed dithiol reactions, suggesting that all members of this subfamily are able to use both the monothiol and the dithiol mechanism. Because of the lack of the C-terminal active site cysteine, monothiol Grxs are not able to perform the reduction of disulfides. Surprisingly, in addition, (de)glutathionylation reactions are not catalyzed by most of the monothiol Grxs (35, 54).
Disulfide reduction
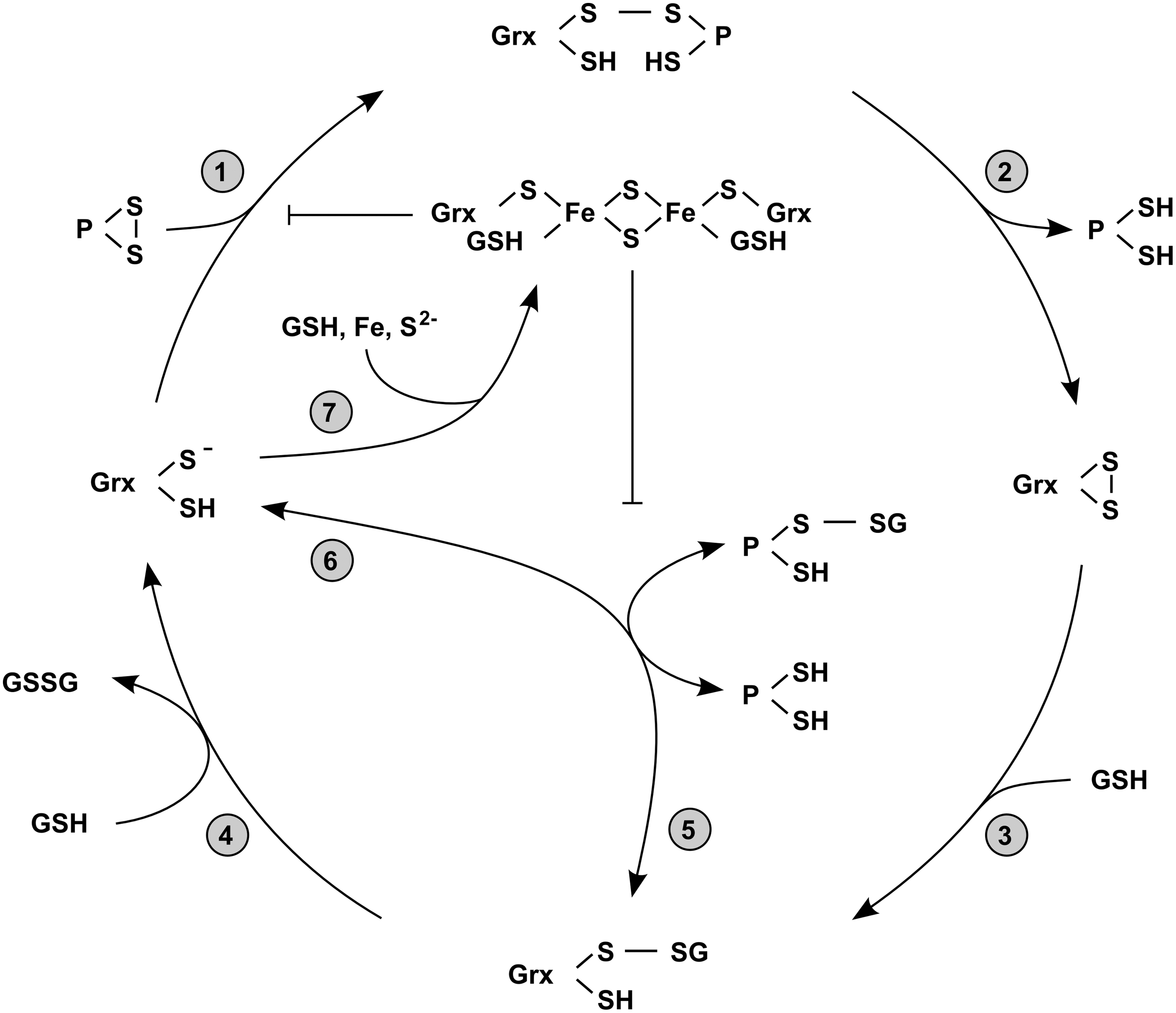
In the first step of a disulfide exchange reaction between Grxs and other proteins, the disulfide bond of the substrate is targeted by a nucleophilic attack of the N-terminal active site cysteine (Fig. 4, Step 1). This cysteine residue has an extraordinarily low pKa value and is solvent accessible, whereas the C-terminal active site cysteine is buried and inaccessible. Due to its low pKa value—around three pH units below the pKa value of free cysteine—the N-terminal active site cysteines of Grxs are present as thiolates at physiological pH (106). The rate constant of thiol/disulfide exchange reactions depends on the nucleophilicity of the attacking reactant and the stability of the leaving group. Thiolates are substantially better nucleophiles compared to thiols, which explains the catalytic efficiency of Grxs in thiol/disulfide exchange reactions at physiological pH. Next, the mixed disulfide between Grx and a substrate formed as result of the nucleophilic attack is reduced by the C-terminal active site cysteine. This step releases the reduced substrate (Fig. 4, Step 2). The remaining disulfide between the two active site cysteines is reduced by a molecule GSH leading to the formation of a mixed disulfide between GSH and the N-terminal active site cysteine of Grx (Fig. 4, Step 3), which is finally reduced by a second molecule of GSH (Fig. 4, Step 4).
Reduction of glutathionylated thiols
Grxs reduce highly specific mixed disulfides containing GSH. Studies using the human Grxs 1 and 2 demonstrated that mixed disulfides containing other compounds are not recognized as substrates (32, 29). GSH-mixed disulfide substrates are not restricted to proteins. Glutathionylated small compounds, for example, Cys-SG, β-hydroxyethyl-SG, or GS-SG, can be reduced by Grxs as well; for a summary, see (90, 56, 54). Different Grxs showed different preferences for either protein-SG or small compound-SG. Whereas human Grx1 seems to be more effective in reduction of mixed disulfides between proteins and GSH, human Grx2 showed a higher affinity to glutathionylated small compounds (48). To reduce a glutathionylated thiol, again the first step is a nucleophilic attack of the N-terminal active site cysteine. In this reaction, the non-GSH substrate constitutes the leaving group. The release of the reduced substrate thus leaves a mixed disulfide between the N-terminal active site cysteine of Grx and GSH (Fig. 4, Step 5) (79). As in the last step of the dithiol reaction mechanism, this mixed disulfide is reduced by a second molecule of GSH (Fig. 4, Step 4).
Glutathionylation
Glutathionylation represents the addition of a GSH molecule to a protein thiol. This post-translational modification can lead to both activation or inhibition of proteins, for example, sarco/endoplasmic reticulum calcium ATPase (SERCA) (1) and mitogen-activated protein kinase/extracellular signal-regulated kinase kinase kinase 1 (MEKK1) (19), respectively. Glutathionylation contributes substantially to the regulation of energy metabolism, ion channel activity, redox signaling, transcription, cytoskeletal assembly, as well as of protein folding (71, 85, 20, 16). In addition, glutathionylation has been suggested as a potential mechanism to protect thiols against further irreversible oxidation (Fig. 3) (43). In line with this suggestion, glutathionylation is enhanced by Grxs in the presence of GSH disulfide (GSSG), mimicking intracellular conditions where the GSH:GSSG ration is decreased (88). Under these conditions, Grxs catalyze the back reaction, that is, the glutathionylation of thiols, equally well (Fig. 4, Step 6).
Iron–sulfur cluster
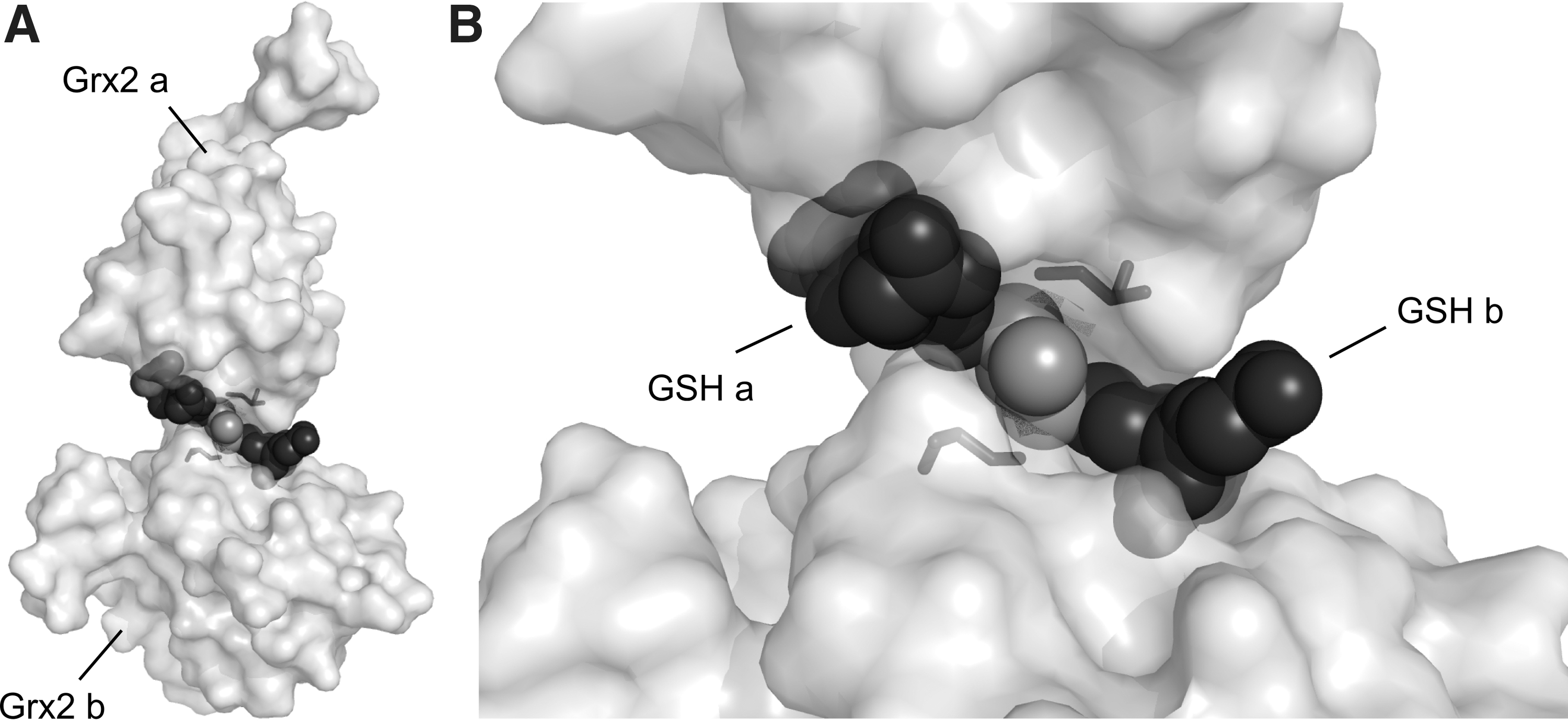
Active site thiols of some Grxs can complex iron–sulfur clusters. Human Grx2 was the first protein of the Trx family described as an iron–sulfur protein (55). Today, we know that a limited number of dithiol Grxs from human, plants, and trypanosomes (55, 86, 14) and essentially all of the monothiol Grxs (35, 54) coordinate a [FeS] cluster. The [2Fe2S]2+ cofactor is ligated between two monomers via the thiols of the N-terminal active site cysteine residues and two noncovalently bound GSH molecules (7, 47, 86) (Fig. 5). GSH bound in the holo–Grx complex is in constant exchange with the free GSH pool, which explains the important role of GSH for [FeS] cluster stability (7). Since the N-terminal active site cysteine is essential for the first catalytic step of thiol/disulfide exchanges, the holo-proteins of these [FeS] clusters coordinating Grxs are enzymatically inactive (Fig. 4, Step 7) (55). Moreover, the ability to bind the cofactor goes along with a reduction of enzymatic activity for the apo-protein. Analysis of a mutant of human Grx2, which was unable to coordinate the cofactor due to the exchange of the active site serine to a proline revealed an almost fourfold increase in enzymatic activity (7). Despite the recent publication of the first [FeS]-Grx harboring a Cys-Pro-Tyr-Cys active site motif (14), the ability of Grxs to coordinate the cofactor correlates well with the presence/absence of a proline residue within the active site (7, 86). The weakening of noncovalent GSH binding by site directed mutagenesis increased the enzymatic activity of human Grx2 as well. By exchange of Thr95 to Arg, one of the two hydrogen bonds formed between Thr95 and GSH (47) was destroyed (7). In addition, the guanidino group of Arg should interfere with the coordination of the carboxy and the amino groups of the Glu residue in GSH. In fact, this mutant of human Grx2 displayed a twofold higher activity compared to the wild-type enzyme (7). Why evolution favored a lower thiol/disulfide exchange activity for the ability to coordinate [FeS] clusters remains a mystery.
Structural changes upon thiol/disulfide exchange
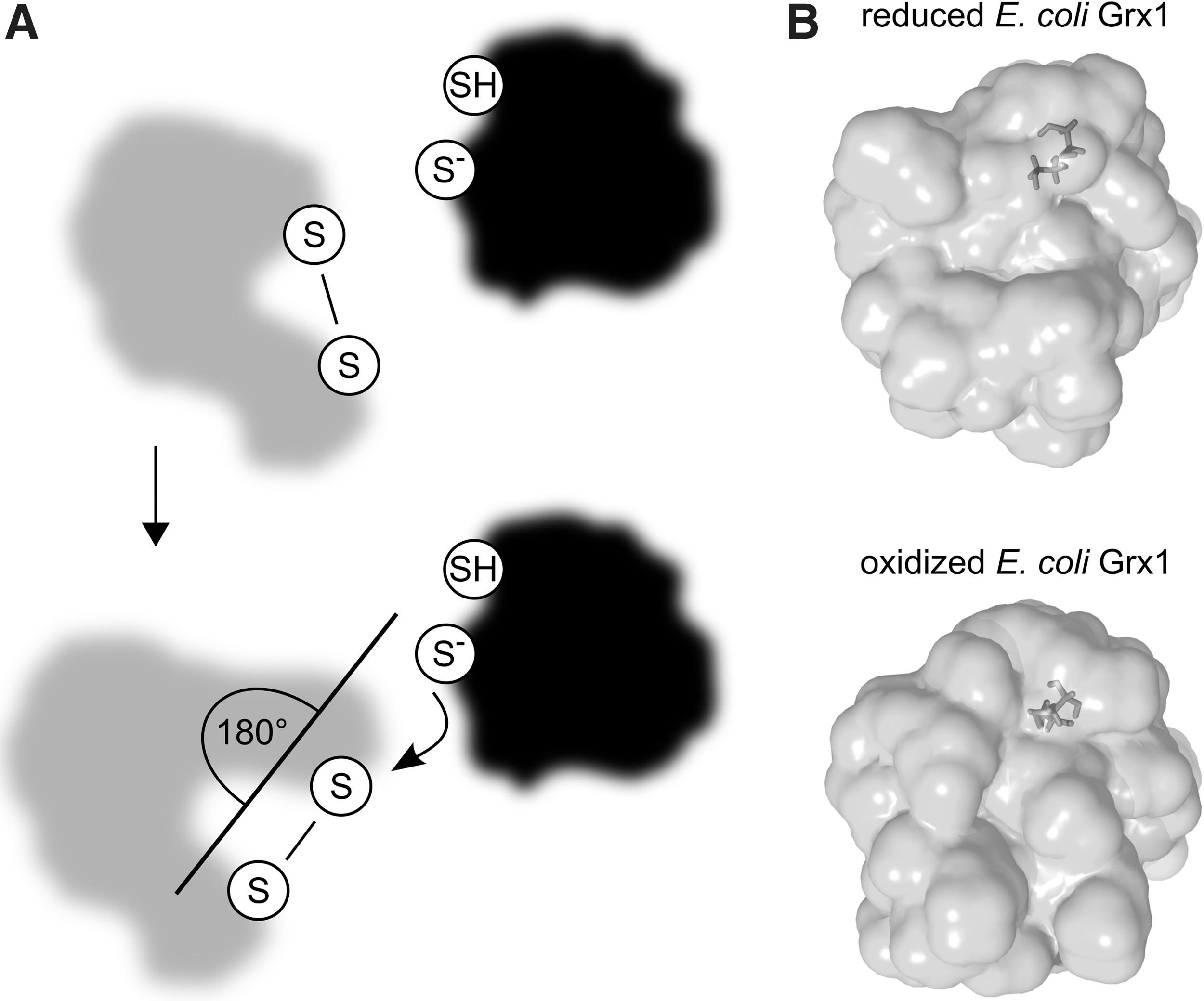
Single molecule force-clamp spectroscopy revealed a potential rate limiting effect of the orientation between the active site of E. coli Trx1 and the substrates disulfide (80). This study suggested the requirement of a reorientation of the N-terminal active site cysteine and the two disulfide bridged cysteines in a 180° angle (Fig. 6A). The rearrangement results in a shortened projection of the disulfide bond of the substrate. This displays the optimal condition for the nucleophilic attack by the N-terminal active site cysteine. Already in 1977 it was reported that changes of both the disulfide bond length and the dihedral angle are important prerequisites to increase efficiency of a nucleophilic attack (78). Because of the high similarities between the protein disulfide reduction of Trxs and Grxs, it can be assumed that these structural conditions are needed for enzymatic efficiency in the dithiol reaction mechanism of both proteins. Thiol/disulfide changes not only induce structural changes within the substrates, but also within Grxs (Fig. 6B). Although the overall structure is quite similar, small changes in E. coli Grx1 and bacteriophage T4 Grx1 are visible comparing the structures of the oxidized and reduced enzymes (104, 102). In a mixed disulfide with GSH, E. coli Grx1 shows more features of the oxidized protein structure (13). The structural changes occur around the active sites and suggest an increased flexibility of this area in the reduced proteins. In addition, surface areas involved in protein–protein interactions are hidden in oxidized Grxs. This indicates a diminished binding affinity once the substrate is reduced and the Grx–substrate complex has to dissociate. The conformational changes leading to a decreased binding capacity also position the C-terminal active site cysteine for nucleophilic attack on the intermediate mixed disulfide.
Measurement of thiol/disulfide exchange reactions catalyzed by Grxs
The thiol/disulfide exchange activity of Grxs can be determined by a limited set of assays. These assays can be used to distinguish between the monothiol and dithiol mechanism. For this purpose, several model substrates are used that are recognized by most of the Grxs. We are not aware of a thorough comparison of rate constants and kinetic data of one Grx with several physiological substrates. Moreover, a comparison between different Grxs measured with one of the available assays is not possible, because of the lack of standard conditions. All assays are in vitro assays, which represents another disadvantage. The establishment of in vivo assays for Grxs and other oxidoreductases will be the biggest challenge during the following years.
RNR assay
Both active site cysteines of Grxs are utilized to reduce E. coli RNR (12); an assay of this reaction therefore assesses the dithiol reaction of Grxs. The assay is based on the formation of deoxycytidine diphosphate (dCDP) from cytidine diphosphate (CDP) by RNR using electrons provided by NADPH, GR, GSH, and Grx (37). Reduction of RNR can be performed using both radioactive labeled substrate and unlabeled substrate. In case of unlabeled CDP, the reaction is followed spectrophotometrically measuring NADPH consumption (38). Using 3H labeled CDP, consumed NADPH is regenerated by glucose 6′-phosphate and glucose 6′-phosphate dehydrogenase. The reaction is followed via determination of the amount of formed 3H-dCDP, after hydrolysis to 3H-dCMP and chromatography on Dowex-50 columns by scintillation counting.
Phosphoadenylysulfate reductase assay
Activity of phosphoadenylylsulfate (PAPS) reductase requires reduction by Grxs via the dithiol mechanism and is measured by determination of 35S-SO3 2− formation from 35S-PAPS (89, 58). To quantify produced 35S-SO3 2− by scintillation counting, sulfite is removed from the assay mixture in form of gaseous SO2 by acidification and absorbed by the base tricotylamine. Since PAPS reductase essentially lacks background activity with dithiotreitol (DTT) as an electron donor, Grx can be re-reduced by DTT in this assay. To measure PAPS reductase activity spectrophotometrically via consumption of NADPH, Grx can be reduced by GR, GSH, and NADPH as a subsequent electron source (82).
Hydroxylethyl disulfide assay
Although the name indicates that the hydroxylethyl disulfide (HED) assay measures the dithiol mechanism of Grxs similar to the RNR and PAPS reductase assays described before, it actually assays the monothiol mechanism. Functional characterization of E. coli Grx1 C14S, a mutant lacking the C-terminal active site cysteine revealed that the preferred substrate in this assay is not the HED itself (12). Instead, it is a mixed disulfide between GSH and β-mercaptoethanol (β-ME-SG) that is formed during preincubation of HED with GSH. The reduction of β-ME-SG by subsequently added Grx can be followed continuously as consumption of NADPH by GR after reduction of the product GSSG (38, 63). By this coupled optical assay, the reduction of any glutathionylated substrate can be measured as well. Besides HED, a variety of proteins and small compounds have been introduced as model substrates to determine Grx activity based on the monothiol mechanism, such as ribonuclease A-SG (62), cysteine-SG (63), and bovine serum albumin-SG (48). Recently, it has been shown that Grx mediated de-glutathionylation of bovine serum albumin glutathionylated with di-eosin-GSSG can be followed fluorimetrically (49).
Physiological and Pathophysiological Conditions Regulated by Grx Mediated Thiol/Disulfide Exchange
Thiol modifications
The impact of Grxs on numerous aspects of life is based on the regulation of reversible oxidative thiol modifications, but not all thiol groups are accessible for those modifications. In addition to surface exposure, some publications claim a specificity of distinct thiols, which is based on the electrostatic and hydrophobic environment of the thiol (33, 97). Two structural motifs, an acid-base motif and a hydrophobic motif, for instance, contribute to the susceptibility of certain thiols to S-nitrosylation (36).
Reactive oxygen species can lead to the formation of inter- or intramolecular disulfides via several mechanisms. Thiols may also react with hydrogen sulfide to form persulfides, with hydrogen peroxide to form sulfenic, sulfinic, and sulfonic acids, or with nitric oxide resulting in S-nitrosylation (Fig. 3). The thiol redox state reflects local redox conditions. In contrast to the extracellular redox potential, the intracellular redox state, although highly dynamic, is more reducing. The two GSH/GSSG redox couple, is one of the major determinators of the cellular redox potential. The tripeptide GSH, γ-glutamyl-cysteinyl-glycine, is the major biological compound with intracellular concentrations up to the millimolar range (66). The two GSH/GSSG redox potential in cells was linked to several cellular functions, such as proliferation (approximately −240 mV), differentiation (approximately −200 mV), or apoptosis (approximately −170 mV) (103). It was demonstrated before that Grxs act as GSH-dependent reductases at −240 mV and as GSSG-dependent oxidases at −170 mV (2). The midpoint potentials of Grxs are close to the above-described cellular potential of the two GSH/GSSG redox couple, in the range of −175 mV (yeast Grx5) (94) to −233 mV (E. coli Grx1) (2). Grxs are therefore ideally suited for the regulation of cellular processes connected to changes in the two GSH/GSSG redox potential.
In the following, we will focus on disulfides and glutathionylated thiols, the two redox modifications reduced by Grxs, and their impact in Grx mediated redox signaling during health and disease. Grxs have been shown to be important for numerous physiological and pathological situations, for example, floral development and plant defense against pathogens via interaction with TGACG motif binding (TGA) transcription factors (105, 74, 75). Unfortunately, only for a limited number of functions both the interaction partners and the underlying mechanisms have been characterized. Established substrates are summarized in the following two chapters.
Disulfides
As described above, the first identified substrate of Grxs was RNR (37). During the catalytic cycle, the reduction of a disulfide formed in the subunit harboring the catalytic center is needed. Grxs have been identified as electron donors for this reduction in E. coli (38), yeast (73) (52), and mammals (41).
Shortly after DNA synthesis, the process of sulfate assimilation has been identified as Grx-dependent (100). Sulfate assimilation summarizes the utilization of inorganic sulfate for the production of sulfur containing amino acids and cofactors in bacteria, fungi, and plants. The second step in this process requires the reduction of activated sulfate to sulfide performed by PAPS reductase. Upon reduction of sulfate, the enzyme gets oxidized resulting in the formation of a disulfide between the two active site cysteines of two monomers (6). This disulfide is a substrate for Grx in E. coli (58), Bacillus subtilis (9), and poplar (87).
Grxs are electron donors for the reduction of Trx in plants (31), peroxiredoxins in plants and human (30, 34), and arsenate reductases, for instance, in the cyanobacterium Synechocystis spec. (60).
Also OxyR-mediated transcription in E. coli is regulated by Grx1. The OxyR transcription factor induces in E. coli and S. typhimurium expression of proteins involved in protection against hydrogen peroxide (21, 15). OxyR is activated by formation of an intramolecular disulfide bond following hydrogen peroxide exposure. Grx1, whose expression is also regulated by OxyR, inactivates the transcription factor by reduction of the disulfide (108).
Recently, the dithiol Grx mechanism has also been described to be important for vertebrate embryonic development (11). The collapsin response mediator protein 2, a key protein in semaphorin 3A signaling regulating axonal outgrowth, has been characterized as a substrate reduced by Grx2. Without Grx2, the collapsin response mediator protein 2 stayed oxidized and the establishment of the axonal scaffold failed leading to neuronal cell death and the impairment of brain development in zebrafish (11).
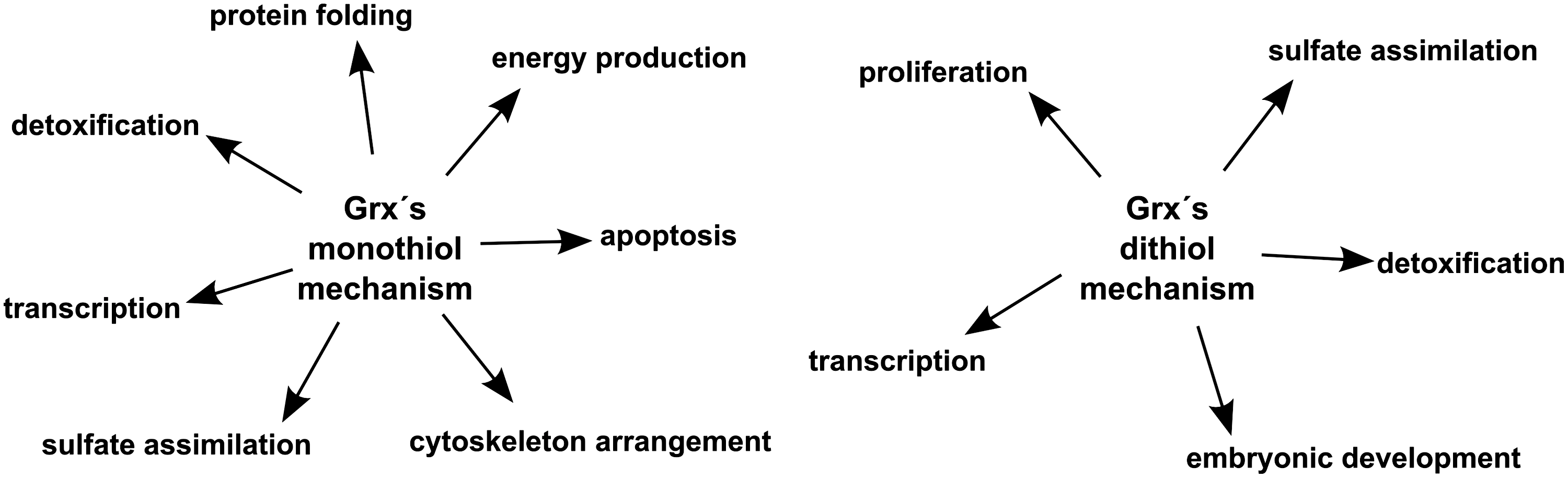
In summary, Grxs performing the reduction of inter- or intramoleular disulfides of the above-mentioned substrates via the dithiol mechanism are important for proliferation, sulfate assimilation, detoxification, transcription, and embryonic development (Fig. 7).
Glutathionylated thiols
The reversible formation of mixed disulfides of protein thiols with GSH is a key mechanism in redox regulation and signaling comparable to reversible protein phosphorylation. As described above, Grxs catalyze both the formation and the reduction of mixed di-sulfides between protein thiols and GSH (88). In general, the reduction of these mixed disulfides is favored, but when the 2GSH/GSSG ratio decreases mixed disulfides are formed (29, 42, 26, 85).
In human and other mammalian cells, glutathionylation has been shown to regulate a number of key proteins and processes in response to redox signals. So far, more than 200 mammalian proteins that are post-translationally modified by the thiol/disulfide exchange with GSH have been reported (28, 70). Thereby, glutathionylation might contribute to a number of the pathophysiological alterations observed in numerous disease conditions. For instance, increased and specific glutathionylation has been demonstrated in Alzheimer's disease (22, 76, 24), Parkinson's disease (10), and several inflammatory diseases (17, 91, 46). Rodriguez-Rocha et al. (84) proposed a role of Grx1 in protection against dopaminergic cell death during Parkinson's disease via glutathionylation of the proteins flightless-1 and RalBP1-associated extracellular protein secretion domain-containing protein 2.
In addition, actin (101), interferon regulator factor 3 (83), complex I of the inner mitochondrial membrane (99, 43) Na,K-ATPase (81), glyceraldehyde 3-phosphate dehydrogenase (18), protein tyrosine phosphatase 1B (3), heat shock protein 70 (27), caspase-3 (40), or Nf-κB signaling through I-κB kinase (65) are regulated by glutathionylation in mammalian cells. Grxs catalyze the (de)-glutathionylation of all of the mentioned proteins (101, 83, 5, 81, 59, 39, 3, 77), thereby contributing to the arrangement of the cytoskeleton, transcription, energy production, protein folding, and apoptosis.
The number of established glutathionylated proteins in plants is increasing due to recent proteomic studies (107, 23, 44). Some of the identified proteins have been characterized as substrates for Grxs, for example, methionine sulfoxide reductase (98), cytosolic glyceraldehyde 3-phosphate dehydrogenase (4), and Trx-f (69). Trx-f is located in chloroplasts and under oxidizing conditions a cysteine residue outside the active site becomes glutathionylated resulting in decreased reduction of Trx-f by ferredoxin-Trx reductase (69).
So far, only few glutathionylated proteins were identified in yeast (92). Some of them are targets for monothiol Grxs (see second to last paragraph, below). The dithiol Grx1p contributes to detoxification and resistance against arsenate via reduction of arsenate reductase (Acr2p) (72).
A study by Kehr et al. (51) identified 493 proteins potentially undergoing glutathionylation in Plasmodium falciparum covering most of the central metabolic pathways, for example, nitrogen and aromatic compound metabolism, or metabolic processes regarding proteins, RNA, lipids, organic acids, ketones, and carbohydrates. Since the proteins were identified by a method based on Grx-induced de-glutathionylation, all the identified glutathionylated proteins are potential substrates of Grx. The de-glutathionylation of ornithine δ-aminotransferase by Grx was confirmed in vitro.
In E. coli, redox regulation via Grx-dependent reversible (de)glutathionylation has been reported for sulfate assimilation through PAPS reductase (57), and for arsenate reduction via an arsenate reductase (R773 ArsC) (94).
Although most characterized monothiol Grxs lack both dithiol as well as monothiol activities (35, 54), few members of this group were able to act via the monothiol mechanism in vitro. The determined efficiencies are extremely low compared to dithiol Grxs, therefore the physiological significance of these enzymatic activities is still elusive. Yeast monothiol Grxs 5, 6, and 7 displayed activity in the HED assay (67) and catalyzed the de-glutathionylation of carbonic anhydrase III (96) and glyceraldehyde-3-phosphate dehydrogenase (93). In a yeast mutant lacking Grx5, glyceraldehyde-3-phosphate dehydrogenase glutathionylation was increased and the recovery of enzyme activity was inhibited (93). Human Grx5 displays very low, yet significant activity in the de-glutathionylation of bovine serum albumin and RNR (49).
In summary, Grxs contribute to transcription, energy production, protein folding, protection against apoptosis, detoxification, sulfate assimilation, and cytoskeleton arrangement via their monothiol reaction mechanism (Fig. 7). Moreover, Grx-mediated de-glutathionylation seems to be important for the progression of several diseases characterized by accumulating redox modifications.
Conclusions
Although the importance of Grxs was demonstrated in several aspects of life, we are still only scratching on the surface of Grxs impact on cellular functions. Many new characteristics of Grxs have been revealed during the last years, which need further investigations, for example, the role of monothiol Grxs and the role of the [FeS] cluster coordinating Grxs. In addition, some Grx functions independent of the thiol/disulfide exchange have been suggested; for instance, the regulation of apoptosis signaling kinase 1 (95). As described above, several hundreds of proteins within all kingdoms of life have been characterized as redox regulated by glutathionylation, but just a small fraction of these proteins have been already determined as substrates of Grxs. Since de-glutathionylation is a unique function of Grxs, it can be assumed that at least the majority, if not all glutathionylated proteins, are reduced and maybe even oxidized by Grxs. Therefore, we are convinced that many substrates reduced by Grxs using either the monothiol or the dithiol mechanisms are awaiting their discovery, which will be closely connected to the development of new tools to visualize redox signaling in vivo.
Footnotes
Acknowledgments
The authors wish to express their gratitude to their former supervisors, academic teachers, and mentors Jens-Dirk Schwenn and Arne Holmgren for their inspiration and continuous support.
Author Disclosure Statement
No competing financial interests exist.