
Abstract
Introduction

The metabolic reactions and energy status of plant cells are intimately connected with cellular and, especially, organellar redox metabolism, since cells rely on chloroplastic and mitochondrial electron transfer pathways for supply of reducing equivalents and adenosine-5′-triphosphate (ATP). In photosynthetic tissues, a particularly important role in the maintenance of cellular redox homeostasis has been assigned to chloroplasts, which utilize light energy to perform a wide variety of energy-demanding steps in the assimilation of carbon, sulfur, and nitrogen, in addition to the biosynthesis of amino acids, hormones, lipids, vitamins, and secondary metabolites. These compounds are vital not only for basic metabolism but also have a great impact on the acclimation status of plant cells. Both the metabolic intermediates and reducing equivalents undergo extensive exchange between chloroplasts, cytoplasm, mitochondria, and peroxisomes (86, 117). Such tight integration of redox metabolism through interorganellar crosstalk is considered one of the key determinants of redox homeostasis in plant cells (30, 117).
Oxidative modulation of redox-active compounds by ROS is another key mechanism that controls the status of cellular redox networks (30, 82). Plants attempt to maintain the cellular redox state fairly reduced, and they possess a multilayered network of antioxidants to maintain ROS and other oxidants at basal levels. The low-molecular weight antioxidants ascorbate and glutathione (GSH) are capable of quenching oxidizing agents chemically and form major redox signaling hubs in plant cells (29, 49, 82). In addition, ROS can be enzymatically detoxified by a range of cytoplasmic and organellar isoforms of antioxidative enzymes, such as superoxide dismutases, ascorbate peroxidases (APXs), peroxiredoxins (PRXs), glutathione peroxidases, and catalases (71). Together, the counteracting oxidizing and reducing activities control redox-dependent changes in plant cells.
Any deviations from the basal cellular redox balance may induce redox signals, which are translated into acclimation responses that continuously readjust cellular functions in short-term and long-term time scales. However, diversion of resources to stress responses may lead to attenuation of growth, and exaggeration of defensive reactions may thus be detrimental to the plant. The ultimate outcome of acclimation responses is therefore tightly controlled by the entire redox signaling network, where chloroplastic, mitochondrial, and peroxisomal signaling systems cross-communicate with redox signals that originate from the apoplast (42, 84, 114, 117, 135). Light- and hormone-induced rapid stomatal movements and redox regulation of photosynthetic membrane protein complexes are well-known examples of mechanisms that optimize plant performance upon transient environmental changes (102, 124). However, when an environmental challenge persists and the capacity of short-term mechanisms exceeds, redox imbalances in different subcellular compartments translates into signals that mediate local and systemic acclimation processes even in unexposed parts of the plant. These responses may be highly specific and generally involve coordinated changes in gene expression, optimization of metabolic pathways, and reprogramming of developmental cascades to best meet the needs of the prevailing environmental cues.
Plants have the capacity to sense environmental changes and receive signals from multiple sources to fine-tune cellular functions upon concurrent environmental changes. On the other hand, abrupt and often highly localized changes in ROS production and cellular redox state may also elicit highly specific stress signals that mediate the onset of defense reactions upon harsh environmental challenges. This has been evidenced as distinct gene expression changes observed in response to specific ROS signals under various experimental stress conditions (58, 66, 129). Increasing evidence also suggests that redox signaling is tightly coupled with hormonal regulation, where the particularly important role of abscisic acid (ABA) in abiotic stress signaling has long been recognized. By now, significant research efforts have started to reveal the molecular components and functional hierarchy in the sensing and signaling of environmental stress. Particularly the importance of intercompartmental crosstalk in cellular signaling is increasingly recognized. In this review, we discuss the currently emerging functional connections between the apoplast and chloroplasts in cellular redox signaling. Moreover, special emphasis is given for light and drought-induced stress responses and their connections with the plant immune system in photosynthetically active green tissues.
NADPH Oxidase as a Source of ROS in Animals and Plants
The apoplastic space is the subcellular compartment where several external stresses and other signals are first sensed. For example, the sensory components in innate immunity in biotic stresses are apoplastic, and also tension-sensing mechanisms active in the sensing of desiccation stress have apoplastic components. The Arabidopsis genome contains about 400 receptor-like kinases (RLK) that have the sensory domain in the apoplast and the kinase domain in the cytoplasm—the molecular and biological function of almost all of them is unknown, but it has been hypothesized that a significant portion of them would have a role in monitoring environmental changes. Apoplastic sensory mechanisms are also known to be connected to apoplastic ROS production. In plants, apoplastic ROS are actively produced through the action of NADPH oxidases and different cell wall peroxidases and they play important roles during plant development and cell growth, abiotic and biotic stress responses and adaptation, and symbiotic interactions (65, 114). The ROS production by the NADPH oxidases is an ancient, conserved mechanism that outdates the evolutionary separation of plants from the lineages leading to animals.
In human phagocytes, NADPH oxidase activity and the consequent generation of ROS contribute to the elimination of micro-organisms in host defense. In the innate immunity of plants, NADPH oxidases also promote pathogen-associated ROS production, however, the effect on disease resistance and cell death varies depending on the plant–pathogen interaction (119, 120, 122). Other ROS-generating enzymes, including peroxidases and amine and oxalate oxidases can also account for apoplastic ROS production. In particular, a role for cell wall peroxidase-dependent apoplastic oxidative burst in plant immunity has recently been established (6, 16, 22, 87). In this review, we will first compare the regulation of NADPH oxidases in animals and plants (Fig 2.). The roles of plant RESPIRATORY BURST OXIDASE HOMOLOGs (RBOHs) in abiotic stress cell signaling and plant immunity are discussed in more detail in later chapters of this review.
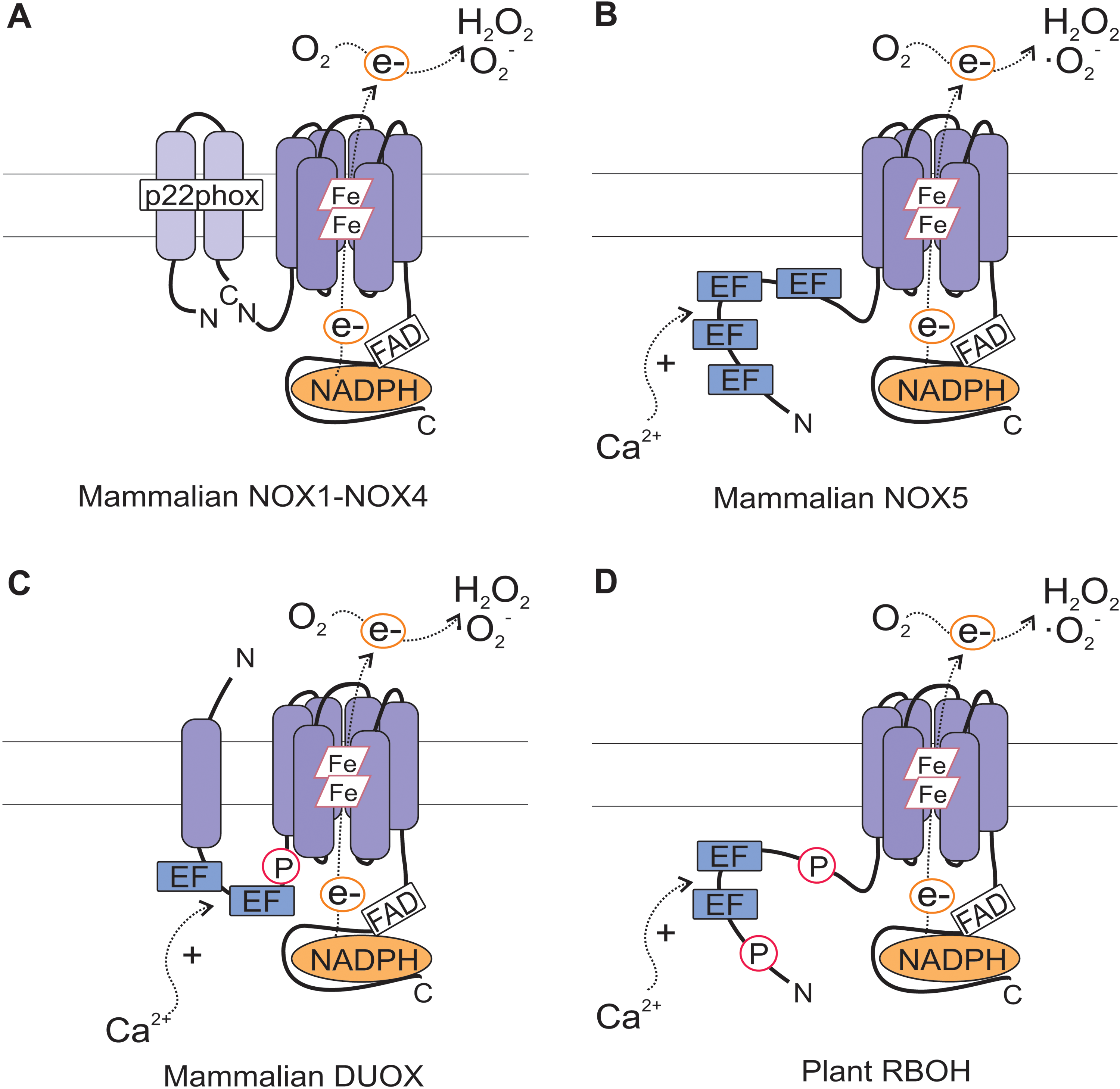
In animals, ROS production is catalyzed by the superoxide-generating NADPH oxidases termed NOXs, also known as RESPIRATORY BURST OXIDASE (RBO). The key redox component of the oxidase, flavocytochrome b 558, is composed of a catalytic subunit gp91phox (also known as NOX2) and a regulatory subunit p22phox. In addition, cytosolic regulatory subunits p40phox, p47phox, p67phox, and the small GTPase Rac are required for activation of the enzyme. Cytosolic C-terminal half of gp91phox forms a flavoprotein domain and harbors the NADPH-binding site, whereas the N-terminal hydrophobic domain binds two heme irons, which transfer electrons from intracellular NADPH to extracellular O2 to form superoxide (·O2 −). In a secondary reaction, O2 − is dismutated to hydrogen peroxide (H2O2).
Since the original discovery of NOX2, six additional members of this family have been identified from humans, namely NOX1, NOX3, NOX4, NOX5, DUOX1, and DUOX2 (Fig. 2). They all contain the catalytic flavocytochrome moiety, also referred to as the Nox domain. NOX1–4 requires p22phox, but unlike NOX1–3, NOX4 does not require additional regulatory subunits for activation. NOX5 and DUOX proteins have been classified into distinct groups based on the presence of additional domains in their N-termini. NOX5 has an N-terminal calmodulin-like domain that contains four elongation factor (EF)-hand motifs for calcium binding. DUOX1 and DUOX2 contain two calcium-binding EF-hands and an additional N-terminal peroxidase-homology domain (47, 59, 60).
The gp91phox is conserved with homologous sequences present across kingdoms including animals, insects, fungi, algae, and plants (47). Plant sequences with homology to gp91phox are termed RBOH for RESPIRATORY BURST OXIDASE HOMOLOG (37, 48, 121, 130, 131) (Table 1). In the genomes of Arabidopsis and rice, RBOHs are encoded by 10 and 9 genes, respectively. Plant RBOHs resemble NOX5 and DUOX proteins, since they contain an extended region with two Ca2+-binding EF-hand motifs in their N-terminus (Fig. 2). Given that plants also lack most of the pg91phox regulatory subunits, RBOHs may be regulated by a manner distinct from that of human gp91phox. Indeed, the importance of protein phosphorylation and Ca2+ in RBOH regulation has been highlighted in several studies. EF-hand motifs of plant RBOHs have the capacity to bind Ca2+ as demonstrated by binding of 45Ca2+ to synthetic versions of Arabidopsis AtRBOHF (48). In vitro studies on tomato and tobacco RBOHs further showed that the oxidase activity can be directly stimulated by Ca2+ (105). In AtRbohC-transfected human embryonic kidney (HEK) 293T cells, ROS production could be stimulated by Ca2+and decreased by substitution of a conserved Ca2+-binding amino acid residue E250A, indicating that Ca2+ binding was required for AtRBOHC function (116). AtRbohC carrying the EF hand mutation also failed to complement the root hair phenotype of atrbohc (rhd2) mutant, reflecting the biological importance of Ca2+ binding to AtRBOHC EF hand motif for root hair tip growth (116).
ABA, abscisic acid; NADPH, nicotinamide adenine dinucleotide phosphate, reduced form; ROS, reactive oxygen species.
Two additional EF-hand-like motifs were identified from the crystal structure of OsRBOHB, where Ca2+ binding to one of the EF hands was shown to induce a conformational change (88). E253A mutant, in which Ca2+ binding was disturbed, did not show Ca2+-induced conformational change (88). E253A mutation in OsRBOHB corresponds to E250A mutation in AtRBOHC, indicating that the Ca2+-mediated conformational change regulates RBOH activity (88, 116). Ca2+ induced activation of AtRBOHD and AtRBOHF has also been demonstrated upon heterologous expression in HEK 293T cells (52, 90).
Two phosphorylation sites have been identified by phosphoproteomics approach on AtRBOHD, S343, and S347, and their phosphorylation was strongly induced by elicitor treatment (85). Complementation of atrbohD with wild-type and S343/347A mutant constructs indicated that phosphorylation of these serine residues is necessary, but not sufficient for full elicitor-induced activation of AtRBOHD in planta. S343 and S347 are conserved among most of the Arabidopsis RBOH isoforms (85), suggesting importance in RBOH activation and also in other biological contexts. In another study using quantitative phosphoproteomics, seven different residues in AtRBOHD were found phosphorylated in response to elicitor treatment (5), however, biological relevance of these phosphorylation events is unclear.
When expressed in HEK 293T cells, AtRBOHD became strongly phosphorylated with a concomitant increase in oxidase activity upon treatment with the protein phosphatase inhibitor calyculin A (CA), indicating that phosphorylation directly activated AtRBOHD (90). Similarly, CA treatment induced ROS production in AtRbohF and AtRbohC (RHD2) transfected HEK 293T cells (52, 116). In vitro kinase assays indicated that S318 and S322 on AtRBOHC were the targets for phosphorylation (116). Evidently, phosphorylation plays a central role in the regulation of RBOH activation. The potato CALCIUM-DEPENDENT PROTEIN KINASE 5 (CDPK5) phosphorylated Serine 82 of StRBOHB and thereby activated the protein in planta (55), whereas in Arabidopsis, the N-terminus of AtRBOHF was phosphorylated by the protein kinase OPEN STOMATA 1 (OST1) (111). However, generally the protein kinases responsible for phosphorylation of the RBOHs have thus far remained unknown.
In conclusion, NADPH oxidase activity and the generation of apoplastic ROS is controlled via calcium-dependent conformational changes and phosphorylation-dependent modulation of specific sites through kinase and phosphatase activities that may themselves respond to a multitude of signals in plant cells. However, the upstream sensory- and activation mechanisms, and the downstream sensory mechanisms connecting ROS and apoplastic redox changes to several different processes known to involve apoplastic ROS production are mostly still unknown. Recent data indicate that the apoplastic ROS production also mediates intercellular communication in local and systemic acclimation (70, 72). In addition, signals from apoplastic ROS production are relayed to chloroplasts upon activation of plant immune signaling (84). In stomatal guard cells, the sensing of apoplastic ROS is also followed by chloroplastic ROS production (124). However, the details of the signaling involved in transmitting the apoplastic ROS signal to the chloroplasts, which seem to amplify the signal, is completely unknown.
Chloroplast as a Source of Cellular Redox Signals in Plants
Inside plant cells, the photosynthetic electron transfer chains of chloroplast thylakoid membranes is a major source of redox-related processes and redox-active compounds in green tissues. According to a traditional view of linear electron transfer (LET), photosystem II (PSII), plastoquinone (Pq), cytochrome b6f complex (Cytb6f), plastocyanin (PC), and photosystem I (PSI) function in series to transfer electrons from water to nicotinamide adenine dinucleotide phosphate, oxidized form (NADP+) to form NADPH. Electron transfer through Cytb6f is coupled with proton pumping to thylakoid lumen, generating a trans-thylakoid proton motive force that drives the ATP synthase. The reducing equivalents produced and ATP are in turn utilized in various biosynthetic and regulatory pathways in chloroplasts and other compartments of plant cells (Fig. 3).

The structure and function of thylakoid membrane protein complexes undergo continuous and highly dynamic modulations, since even slight variations for example, in light, wind, or temperature conditions constantly modify the accumulation of redox-active intermediates in the photosynthetic electron transfer chain. Without efficient regulatory mechanisms, this would lead to severe over-reduction or over-oxidation of electron transfer components. Pathways of cyclic electron transfer (CET) redirect electrons from PSI to Cytb6f, thus promoting the formation of proton gradient across the thylakoid membrane without production of reducing equivalents (40). Even though the exact molecular mechanisms of CET in C3 plants are not fully understood, it is considered a major contributor to the cellular ATP/NADPH ratio (28, 40). Further, CET may become enhanced under stress conditions when the demand for ATP is increased (28, 40). The two main pathways of cyclic electron flow involve thylakoid NADPH dehydrogenase and PROTON GRADIENT REGULATION 5 (PGR5), both of which also have a fundamental role in balancing the function of LET (17, 79). PGR5 was additionally proven indispensable for prevention of light-induced oxidative damage in PSI under fluctuating light conditions (112).
Besides driving the production of ATP, acidification of thylakoid lumen is also vital for photoprotection, since it promotes the onset of protective dissipation of excess excitation energy through PsbS and xanthophyll-dependent nonphotochemical quenching (NPQ) of excitation energy (81). In addition, plants maintain redox balance in chloroplasts through strictly redox-regulated phosphorylation of light-harvesting (LHCII) antenna proteins by the STATE TRANSITION 7 (STN7) kinase, which becomes activated upon reduction of the Pq pool, and inhibited via thioredoxin activity upon reduction of chloroplast stroma (97, 101). Besides the classical reduction-based regulation by thioredoxins (12, 63, 96), oxidation of protein thiols by ROS may equally well alter the activity of enzymes and regulatory proteins. Recently, Dangoor et al. (19) identified light-induced oxidation of a thioredoxin-like protein AHCT1 as a redox-sensory mechanism in chloroplasts. Accordingly, a complex interplay among redox-dependent short-term regulation mechanisms is recognized as a highly tunable system that allows operation of photosynthetic processes with optimal efficiency in chloroplasts (102, 118).
Despite the large repertoire of regulatory mechanisms, photosynthetic electron transfer chain may still undergo major changes in the redox potential, and the highly energetic nature of electron transfer reactions inevitably promotes formation of ROS in chloroplasts. Rather than causing oxidative damage, however, ROS accumulation is increasingly understood as a component of oxidative and redox signaling, which is one of the key mechanisms in the relay of stress signals in photosynthetic tissues (30, 100).
At the acceptor side of PSI, transfer of electrons to molecular oxygen results in formation of superoxide and H2O2 in chloroplast stroma (3). In the reaction center of PSII, excited triplet reaction center chlorophyll P680 and molecular oxygen may react to form singlet oxygen (1O2), which is highly reactive and has been associated with light-induced oxidative damage and photoinhibition of PSII (39). Currently, increasing evidence points to a key signaling role for singlet oxygen arising from PSII under abiotic and biotic stresses in plants (1, 50, 84, 100). To control photodamage to PSII, the LHCII antenna binds carotenoids, xanthophylls, and tocopherol that quench triplet chlorophyll and alleviate the formation of 1O2 in the thylakoid membrane (1, 100).
Production of singlet oxygen and photodamage to PSII are linked to the high turnover rate of the reaction center protein D1, which becomes degraded and replaced by de novo protein synthesis in the so-called PSII repair cycle (78). Limiting the damage to the D1 protein is generally considered a mechanism to avoid more extensive oxidative damage to the photosynthetic electron transfer chain. Components related to the repair or photoprotection of PSII have also been connected with the onset of immune reactions in plants. These include the FtsH protease DS9 associated with degradation of the damaged D1 protein in tobacco (109), NPQ in Arabidopsis (35), and the CALCIUM SENSING RECEPTOR (CAS) required for photoprotection of PSII in the green alga Chlamydomonas reinhardtii (98). It is therefore tempting to speculate that the vulnerability of PSII was conserved during the evolution of higher plants, since it forms a powerful regulatory mechanism in the process of eliciting ROS burst involved in defense mechanisms against plant pathogens. The characteristic inactivation of chloroplast APXs and PRXs upon abrupt accumulation of H2O2 could provide plants with a mechanism to further amplify the ROS burst in chloroplasts (3, 53).
Crosstalk in Chloroplast Redox Signaling in Light Acclimation
Naturally-grown plants are frequently exposed to light stress caused by excess excitation energy that cannot be consumed by carbon metabolism. Moreover, different biotic and abiotic stresses may affect photosynthesis and other enzymatic activities in chloroplasts and promote conditions where excitation energy is in excess. The extent of light stress can therefore vary considerably depending on light intensity and the plant's capability to consume the reducing equivalents, which is largely determined by the preacclimation status of the plant.
When the concept of light stress is considered, moderate light stress can be expected to result in saturation of carbon metabolism and consequent reduction of electron carriers and thiol redox regulators in chloroplasts. Thus, reduction of the Pq-pool is commonly observed upon a sudden exposure of plants to high irradiance levels, and this phenomenon has been correlated with gene expression changes in the nucleus (26, 76). Extreme photo-oxidative stress in turn manifests itself as enhanced accumulation of ROS, thereby promoting oxidative conditions in chloroplasts.
In time scales from seconds to minutes, redox-dependent adjustments in the photosynthetic light harvesting and activity of stromal enzymes aim at balancing the production and consumption of reducing equivalents, thereby preventing unnecessary stress signals from chloroplasts (102, 118). Photosynthetic carbon metabolism is also tightly coupled with interorganellar metabolic shuttles that coordinate the production and dissipation of reducing power, and have a strong impact on cellular redox signaling activities in plant cells (Fig. 4). The malate/oxaloacetate shuttle mediates excess reducing equivalents to the mitochondria, where they become dissipated through electron transfer bypasses including the alternative oxidase and the uncoupler complex (107, 115, 117). If, however, the deviation from redox balance persists, signaling cascades become activated, promoting the onset of acclimation processes by regulating the expression of organellar and nuclear genes (25, 99). A typical response to moderately high light, for example, involves downregulation of genes encoding components of photosynthetic light-harvesting and a concomitant upregulation of genes related to carbon metabolism and protective systems (26, 51, 103).

In natural habitats, light stress is often paralleled by increased desiccation or even drought stress. Changes in leaf water status result in stomatal closure and the consequent limitation in the availability of CO2 may slow down carbon assimilation thus promoting over-excitation of the photosynthetic machinery. On the other hand, stomatal closure and over-reduction of the photosynthetic electron transfer components may promote photorespiration, which alleviates the redox poise in chloroplasts, but leads to production of H2O2 in peroxisomes (27, 28, 83). In this respect, it is intriguing that signals elicited upon accumulation of photorespiratory H2O2 in peroxisomes repressed the expression of genes related to the biosynthesis of anthocyanins, which act as protective pigments and may also have antioxidative activities (125). Rather, peroxisomal H2O2 promotes the onset of cell death and has a strong impact on salicylic acid (SA)-dependent signaling in plant immunity (14, 15). Severely stressed or damaged chloroplasts may also rapidly respond to elicit “ROS signals” to induce rapid defense reactions against biotic and abiotic stresses. Indeed, besides mediating redox signals upon abiotic stress conditions, organelles carry out vital functions in determining appropriate immune reactions against a variety of biotic stress agents (Fig 4) (44, 67, 68).
Photosynthetic organisms have evolved a multitude of mechanisms for sensing, signaling, and responding to a wide range of redox imbalances, including over-reduction or over-oxidation of intersystem electron transfer components, PSI electron acceptors, and thiol redox components and formation of ROS, all of which are inevitable in the course of light-driven metabolic reactions in chloroplasts (Fig. 3). However, the molecular identity of the sensors and signaling components and the molecular signaling interactions are still largely unknown. Comparison of stress-induced gene expression changes by complementary DNA microarray analysis demonstrated that up to 70% of the high-light-inducible genes were also upregulated under drought stress (51), reflecting the extensive overlap observed among light and ABA-regulated gene expression profiles (32). Accordingly, the ABA-response element is significantly over-represented among the promoter regions of genes that respond to light-induced chloroplast signals (56). Thus, crosstalk between light stress and drought-induced pathways of organellar signaling seem particularly tightly connected among hormonal signaling networks.
The use of genetic approach has advanced the identification of signaling components that participate in the relay of information from chloroplasts to the nucleus. Identification of the conditional Arabidopsis mutant fluorescent (flu) provided a model system in which dark-induced accumulation of protochlorophyllide causes release of 1O2 upon re-illumination of flu seedlings. This has allowed studies on singlet oxygen signaling in chloroplasts (91). The accumulation of 1O2 lead to rapid transcriptional reprogramming and programmed cell death (PCD), evidenced as microlesions in green tissues (50, 91). The genes induced are largely distinct from those induced by H2O2 or ·O2 − and include both SA and jasmonic acid defense marker genes (21, 58), suggesting that 1O2 mediates specific ROS-dependent interactions in plant immunity. Identified components in chloroplast singlet oxygen signaling include EXECUTER1, EXECUTER2 and CRYPTOCHROME1, which also modulates the high light response in Arabidopsis (20, 54, 61, 91).
Pharmacological approach with norfluorazon (NF) has also been utilized as a tool to identify chloroplast signaling components (56, 89, 106, 113). NF inhibits carotenoid biosynthesis (13) and thus causes photo-oxidative damage to chloroplasts, which correlates with suppression of photosynthesis-associated genes in the nucleus (56, 113). A screen for mutants that express photosynthesis-related genes despite the NF-induced photobleaching led to identification of genomes uncoupled (gun) mutants, most of which were affected in the tetrapyrrole pathway (56, 73, 113) (Table 1). Through a yet unknown mechanism, GUN2-6-dependent reactions of chlorophyll biosynthesis appear to modulate ROS or redox-dependent signaling cascades in chloroplasts (74, 75).
Unlike the tetrapyrrole pathway-related GUN2-5, GUN1 is a chloroplast-localized pentatricopeptide-repeat protein that connects the status of chloroplast gene expression with the regulation of nuclear genes, classically exemplified by lhcb2 encoding the major light-harvesting antenna protein of PSII (56). Similar signaling effects can be obtained by using lincomycin, which is an inhibitor of chloroplast translation (56, 77). The GUN1 pathway was suggested to operate through the ABA-responsive transcription factor ABSCISIC ACID-INSENSITIVE 4 (ABI4), providing a point of crosstalk with ABA signaling (56). In fact, organellar redox signals from both chloroplasts and mitochondria are known to converge at ABI4 in the nucleus (49).
Induction of the antioxidant enzyme APX2 is a classical marker for high light stress (46). Studies utilizing a transgenic Arabidopsis expressing luciferase (LUC) under the promoter of APX2 (pAPX2:LUC) as a tool revealed that the induction of APX2 is governed by overlapping signals from chloroplastic and apoplastic ROS, GSH metabolism, and ABA signaling (4, 24, 32, 46, 103). An important step forward was the identification of SAL1, an enzyme that controls the levels of 3′-phosphoadenosine 5′phosphate (PAP), a mobile metabolite capable of translocating from chloroplasts to the nucleus (24). PAP induces expression of APX2, accumulates under high light and drought stresses, and is likely to exert its signaling effects by inhibiting exonuclease activity in the nucleus (24). The sal1 mutant turned out to be allelic to alx8 and fry1 with previously assigned functions in the regulation of RNA silencing pathways in the nucleus (38).
The sal1 phenotype resembles high light acclimated plants with reduced rosette size, compact morphology, and tolerance to drought stress (24, 103, 126). Additionally, sal1 shows constitutive upregulation of a specific subset of HL-responsive genes, whereas the expression of lhcb genes is not changed and the transcript profile does not correlate with that induced by lincomycin. The PAP-dependent pathway thus seems to operate in parallel with other retrograde signaling pathways. Indeed, SAL1 was found to regulate nuclear gene expression in both ABA-dependent and ABA-independent manner (24). A range of chloroplast signals originating from various processes presumably allow specific adjustments depending on the eliciting signal and environmental condition. All together, the light-dependent redox-signaling pathways are tightly integrated with networks that control biotic and abiotic stress responses in plants.
Functional Connections Between Apoplastic and Chloroplastic Processes in ROS Signaling
Different environmental stresses may provoke similar initial signaling mechanisms, which gain their specificity through inputs from other signaling pathways. On the other hand, distinct signals that arise from specific recognition events may have extensive crosstalk among the signaling networks, and even converge on common components. Clues to similarities in signaling effects in the course of stress responses can be obtained by comparing gene expression profiles from the wealth of microarray datasets that have been deposited into publicly available databases.
Changes in redox signaling pathways and ROS metabolism represent common nominators for different stress responses, including those induced by light, salinity, drought, cold, osmotic stress, or plant pathogens. To shed light on commonalities among plant responses to chloroplastic and apoplastic redox signals under such stress conditions, we analyzed microarray datasets from ArrayExpress, Gene Expression Omnibus, and NascArrays [see full description of the data sets and the applied preprocessing in Ref. (34)], and compared experiments that have utilized chemical ROS-inducing systems or in vivo stress treatments to assess transcriptomic adjustments in Arabidopsis plants.
Of the chemical treatments included in the analysis, NF causes photo-oxidation of chloroplasts in illuminated leaves. Methyl viologen (MV), also called paraquat (PQ) is an herbicide that induces generation of ·O2 − and H2O2 through electron transfer reactions in chloroplasts and mitochondria. Rotenone in turn promotes mitochondrial ROS production by blocking the respiratory electron transport chain. Flg22 is a short peptide derived from bacterial flagellin and generally used as a tool to study receptor signaling and oxidative burst in pathogen-associated molecular pattern (PAMP) triggered immunity (8). Similar ROS signaling effects can be achieved by exposure to ozone (O3), which enters the leaves through stomatal pores and degrades into H2O2 and ·O2 − upon entry into the apoplast. Thus, O3 has been widely used as a tool to study the role of apoplastic ROS in plant cell signaling (7, 43, 124, 128, 129).
Altogether 138 genes with at least twofold change in transcript abundance and a statistically significant fold change (Benjamini-Hochberg adjusted p-value<0.05) in at least one of the experiments were included in hierarchical clustering in two dimensions; the transcripts and the experiments (Fig. 5). This revealed temporally highly dynamic transcriptional responses to treatments, which forms two main clusters. Cluster 2 comprises of acute responses at early time points to cold, salt, UV light, and MV-induced stresses, which clustered together with unchallenged zat12 mutant plants, 6 h time points of MV and drought, and different time points of treatment with rotenone.

Cluster 1 divides into two subclusters. In subcluster 1A, the response to 1-h treatment with flg22 is most similar to rapid responses to osmotic shock and cold stress (Fig. 5). Four-hour treatment with flg22 instead groups together with NF, ozone, H2O2, and later 12-h time points of MV and drought stress. These experiments were distinguished from 3–12 h of cold stressed shoots and all time points of salt-stressed roots, which form a distinct branch inside the subcluster 1A. In subcluster 1B, acclimation responses to osmotic, salt, and drought stresses cluster together with ABA treatment of shoots.
As different stress conditions rarely occur only one at a time in nature, it is conceivable that responses to environmental changes are mediated through overlapping signaling pathways and involve both synergistic and antagonistic responses. Similarities among signaling mechanisms and responses may however also be explained by similar physiological stress symptoms and subcellular origin of the stress signals. The transcriptional response of wild-type Arabidopsis plants to NF is characterized by distinct downregulation of genes related to chloroplastic functions, and a similar transcriptional reprogramming is evident in the horizontal gene cluster II also for treatments with flg22, ozone and H2O2 (Fig. 5). The latter three induce their initial signaling effects in the apoplast, where ozone and H2O2 mimic the RBOH-dependent apoplastic oxidative burst elicited for example, upon recognition of a bacterial flagellin in the extracellular space. The apparent downregulation of photosynthesis-associated genes is therefore well in agreement with the reported downregulation of photosynthesis and a shift toward nonassimilatory metabolism in response to infection by various types of pathogens (10). How the downregulation of photosynthesis-related genes and upregulation of immune responses are elicited has, however, long remained a key outstanding question in studies concerning plant immunity.
Recognition of PAMPs by plasma membrane-spanning pattern recognition receptors elicits a biphasic oxidative burst composed of fast and slow phases; the first one has a maximum at ∼1 h and the second begins around 4 h and lasts for several hours and correlates with the onset of defense reactions in plants (36) (Fig. 6). PAMP-induced changes in host gene expression attempt to prevent the microbe from colonizing the host tissue (9). Host-adapted pathogens secrete effector molecules that target the mechanisms of PAMP-triggered immunity. Plants, however, have co-evolved mechanisms that specifically recognize the effectors and elicit effector-triggered immunity, a second layer of immune reactions accompanied by the second and often more prominent burst in ROS and hypersensitive cell death at the attempted site of infection (41, 133).
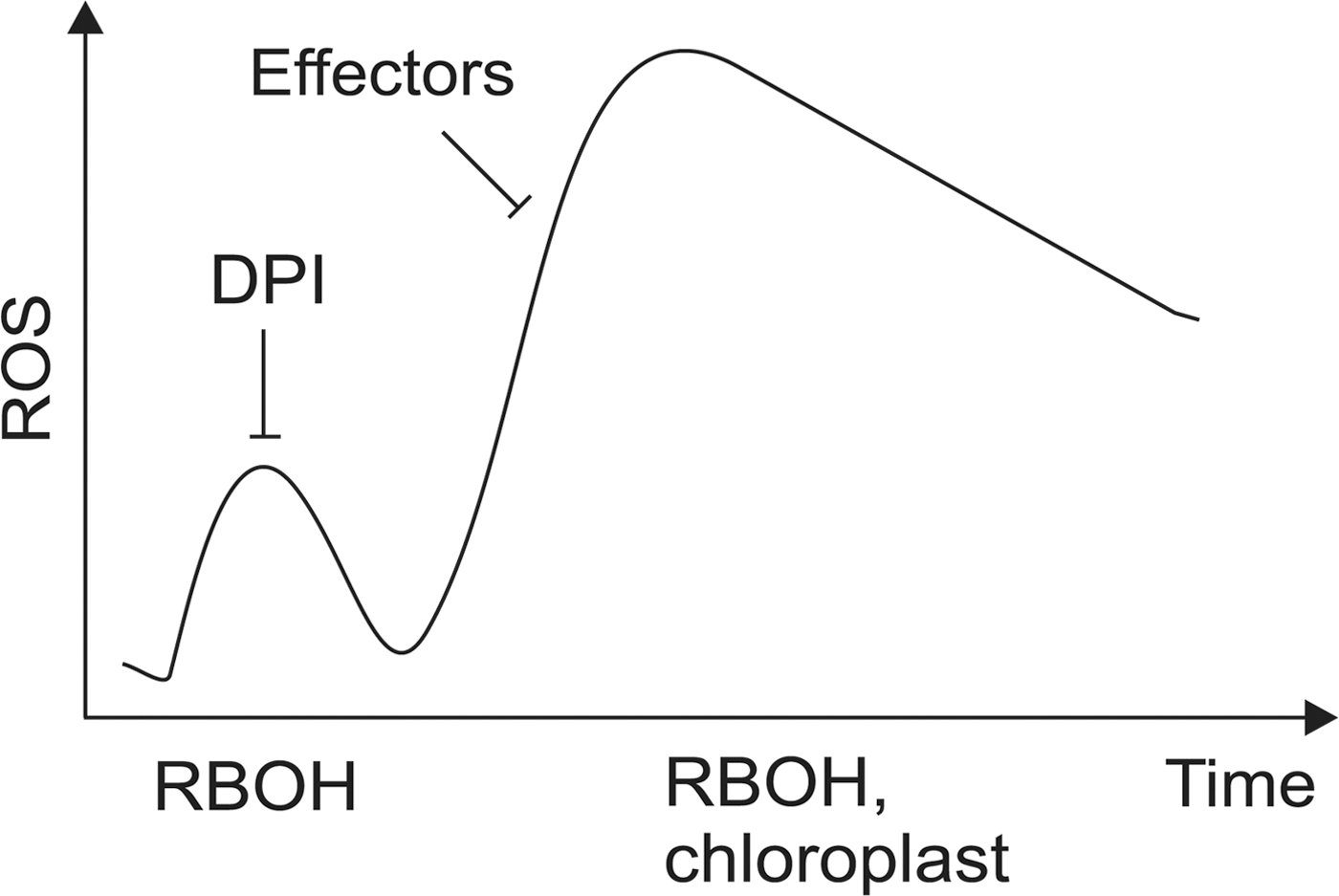
During the early stages of infection, the rapid burst in ROS accumulation is triggered by plasma membrane-bound NADPH oxidase and occurs in the apoplast (120). The source of the slow ROS burst however has not been unequivocally demonstrated, albeit the contribution of chloroplastic and peroxisomal ROS production to plant immunity has long been recognized (15, 44, 46, 135).
Ozone exposure initiates a biphasic oxidative burst, similar to that induced by pathogen challenge (95, 108, 127), and O3-induced lesion formation displays the characteristic morphological and biochemical hallmarks of PCD (92, 93). Joo et al. (42) analyzed the sources of O3-induced oxidative burst and the role of RBOHs and heterotrimeric G-proteins in this process and found that chloroplasts can mediate and even amplify ROS signals that originate from the apoplast. Loss of Gα or Gβ subunit of the heterotrimeric G protein markedly attenuated the early phase of O3-induced oxidative burst arising from chloroplasts. Only the Gα subunit was, however, required for the late phase of the oxidative burst. Consistent with this, plants lacking the Gα protein were also more resistant to O3-induced tissue damage (42).
Recently, Nomura et al. (84) showed that extracellular stress signals are rapidly mediated from the apoplast to chloroplasts, where they induce specific calcium signatures in the soluble stroma. Treatment of Arabidopsis seedlings with flg22, which rapidly activates apoplastic RBOH-dependent ROS production (132), first induced a rapid and transient increase in cytosolic calcium concentration, which was followed by a more long-lasting increase in the concentration of free calcium in chloroplast stroma (84). The PAMP-induced transients in chloroplastic calcium levels were largely absent in cas-1 mutant plants deficient in the thylakoid-bound CAS (84). The function of CAS was also proven indispensable for flg22-induced suppression of photosynthesis-related genes and induction of SA biosynthesis and the subsequent upregulation of the pathogenesis-related genes PR1 and PR2 (84). Microarray analysis further revealed that 2 h after the application of flg22, one-third of the genes with reduced induction in cas-1 overlapped with genes induced by 1O2 in the flu system (58, 84, 91). Intriguingly, cas-1 plants also displayed reduced hypersensitive reaction (HR) cell death in response to an incompatible interaction with the bacterial pathogen Pst DC3000 expressing the effector protein avrRpt2 (84).
It seems conceivable that PAMP-induced signals on the plasma membrane elicit an immediate cytoplasmic calcium transient, which enhances NADPH oxidase activity and ROS burst in the apoplast (Fig. 7). Through a yet unidentified mechanism, these signaling events are relayed into the chloroplasts, where they elicit a more durable increase in stromal calcium concentration. This seems to provoke the second ROS-burst, which manifests itself as a CAS-dependent chloroplastic 1O2 signal that modulates the onset of SA-dependent defense genes and is also required to induce HR cell death in plants (84).

Besides biotic stress responses, specific calcium signatures in chloroplast stroma seem to modulate abiotic stress responses to cold, salt, and hyperosmolarity as well (84). It is therefore not surprising that early responses to osmotic stress and cold clustered together with the early response to flg22 (Fig. 5). By utilizing estrogen-inducible silencing of THYLAKOID APX (tAPX) to assess the effects of abrupt accumulation of H2O2 in chloroplasts, Maruta et al. (66) also found increased expression of cold and pathogenesis-related genes. However, expression of CBF1/DREB1B, encoding one of the main transcription factors mediating cold responses, was suppressed rendering the mutant plants with diminished tAPX levels sensitive to cold stress (66). Cold stress enhances the formation of 1O2 (18) and H2O2, the antagonistic effects of which on 1O2 driven signaling have been well-documented (58). It is therefore possible that low-temperature-induced inhibition of Calvin cycle and the consequent production of H2O2 may negatively regulate cold acclimation.
Apoplast-Chloroplast Continuum in Systemic Signaling
Besides eliciting local responses, chloroplasts also initiate systemic signals that spread through the vasculature and confer stress resistance in shaded parts of the plant (Fig. 8) (45, 46, 76, 104). Such preacclimation mechanism is termed systemic acquired acclimation, which shares a level of commonalities with systemic acquired resistance (SAR), which becomes induced upon exposure of plants to various types of pathogens (23, 64, 76). A pioneering observation of systemic light signaling connected accumulation of H2O2 in the chloroplasts of bundle sheath cells with acquired tolerance against photo-oxidative stress in Arabidopsis leaves (31, 46). Subsequently, ABA synthesized in vascular parenchyma was shown to interact with adjacent bundle sheath cells where it induced ROS production in the apoplast, most likely through NADPH oxidase activity (32). This signaling interaction most likely also involves heterotrimeric G-proteins (32, 42).

In addition to their role in governing local responses, a role for NOXs as mediators of rapid systemic signaling over long distances has recently emerged, a concept that has been called the “ROS wave” (70, 72). Using a reporter system composed of a luciferase reporter gene driven by a ROS-responsive ZAT12promoter, Miller et al. (70) observed that application of local stress treatment led to detection of luciferase activity not only at the site of stress application but also at systemic untreated tissues. The signal propagation was rapid, traveling at a rate of 8.4 centimeters per minute along the stem axis in both directions. This systemic signal was initiated by wounding, cold, heat, high light, and salinity and in all cases it was dependent on AtRbohD. Apoplastic ROS accumulation coincided with the propagation of the signal in wild-type plants; however, both local and distal ROS accumulation was suppressed in atrbohD mutant plants. Further, application of the NADPH oxidase inhibitor diphenyliodonium (DPI) at the wound site or at distal locations inhibited the initiation or progression of the signal, respectively. Thus, AtRBOHD-generated ROS can trigger and maintain an auto-propagating ROS wave that travels from cell to cell across long distances.
In shaded leaves, the SAA response is mediated by the ZAT10 transcription factor, which accumulates in leaf vasculature within 5 min of the onset of light stress (104). The induction of ZAT10 correlated with enhanced expression of antioxidant genes and increased tolerance against light-induced damage of PSII in the light-exposed leaves (104). APX2 was one of the first genes shown to be systemically upregulated in the vascular tissue of distal leaves in response to H2O2 accumulating during local HL exposure (46). APX2 expression is induced in response to multiple signals including ABA and H2O2 and moderate heat and subsequent H2O2 production in the apoplast (4, 31). In HL, APX2 also responds to reduction of the Pq pool and the PAP-dependent retrograde signaling pathway (31, 46, 24). More recently, the high light inducible expression of APX2 was found to be regulated positively by OST1 and negatively by the G protein coupled receptor (32). In ost1-1 mutants, APX2 was not induced under HL, whereas the high light markers ELIP1 and HSP17,6 were expressed like in wild-type plants (32). Thus, mechanisms governing systemic light acclimation and stomatal movements seem to share common components in ABA signaling (Fig. 8).
Stomatal Guard Cells as a Model to Explore Cellular ROS and Redox Signaling
As described above, extensive research efforts have revealed key components and interactions among apoplastic and chloroplastic redox signaling mechanisms. Even so, major gaps still remain in our understanding of signaling continuums between the different cellular compartments. Many of the apoplastic and chloroplastic signaling mechanisms identified thus far are also present in stomatal guard cells, and are known to mediate the regulation of stomatal aperture (32, 42, 80). Guard cells therefore comprise a single cell system that can be used as a model to explore these signaling interactions in more detail.
Pairs of two guard cells form stomatal pores, which control the influx of CO2 and the loss of water vapor through plant leaf epidermis. Stomatal aperture is regulated by a number of environmental factors. For example, closure of stomata under drought conditions protects the plant from desiccation. High ambient CO2 levels in turn promote stomatal closure to limit unnecessary increases in the leaf intra-cellular CO2 concentration. Stomata also restrict the entry of the air pollutant ozone and certain pathogens into the apoplast of plant cells. Related to these, ROS are important signals mediating stomatal movements in response to various external and internal factors.
The phytohormone ABA plays a central role in regulating plant water status upon drought conditions through promotion of stomatal closure and inhibition of stomatal opening. Biosynthesis of ABA is enhanced upon drought conditions. Foliar ABA accumulates in guard cells triggering the closure of the stomata thereby preventing transpirational water loss. ROS function as second messengers in ABA signaling in guard cells. There are several reports showing the integral role of ROS in the regulation of stomatal aperture. Early reports indicated that treatment with MV, which generates ·O2 − and H2O2 in chloroplasts, regulated stomatal aperture (2, 69, 94), ABA has been shown to induce the production of H2O2 in Arabidopsis and Vicia faba guard cells (57, 80, 94, 134), and application of NADPH oxidase inhibitor DPI inhibited ABA-induced stomatal closure, suggesting a role for RBOHs in the process (94, 134). Using fluorescence-based assay, Zhang et al. (134), detected ABA-induced H2O2 accumulation first in the chloroplast, followed by other parts of the guard cells. Increase in fluorescence was also detected in epidermal cells surrounding the guard cells. Addition of DPI prior to ABA application reduced the overall fluorescence of guard cells and adjacent cells; however, ROS accumulation was still clearly detected in the chloroplasts. These results suggested NADPH oxidases and chloroplasts as sources of ROS in ABA-induced stomatal closure (134). The hierarchy of the events is not, however, completely clear. The results of Joo et al. (42) and Nomura et al. (84) suggest that the NADPH oxidase dependent apoplastic ROS burst is followed by the chloroplastic ROS production and that there is a regulatory hierarchy and transmission of a signal from the apoplast to the chloroplast between these processes. This is actually not so surprising since, unlike in schematic figures, as in Figure 7, where the chloroplast is usually drawn in the center of the cell, in reality (as shown in Fig. 8), the chloroplast envelope is almost in membrane contact with the plasma membrane that should allow a rapid and efficient communication between the apoplast and the chloroplast.
Genetic evidence for a role of NADPH oxidases in ABA-induced stomatal closure was provided by a study by Kwak et al. (57). Arabidopsis AtRbohD and AtRbohF are expressed in the guard cells and expression of both genes was upregulated in guard cells in response to ABA treatment. ABA-induced stomatal closure was partially impaired in atrbohF mutant compared with wild-type and this impairment was stronger in atrbohD/F double mutant. Unlike wild-type, atrbohD/F double mutant failed to produce ROS in response to ABA treatment, suggesting that AtRBOHD and AtRBOHF are involved in the ABA-induced ROS production in guard cells, a notion that had been previously postulated based on studies using DPI as NADPH oxidase inhibitor (94, 134). Further, guard cells of atrbohD and atrbohF single and double mutants responded to exogenously applied ROS as wild-type indicating that H2O2 can rescue WT responses in the mutant plants (57). This established the role for NADPH oxidase-produced ROS as second messengers in guard cell ABA signaling leading to stomatal closure. The formation of ROS by the activity of the plasma membrane NADPH oxidase AtRBOHD and/or AtRBOHF, which are activated by perception of the bacterial flagellin (or the flg22 peptide) by the FLAGELLIN SENSITIVE 2 (FLS2)-receptor, lead to rapid stomatal closure most likely to prevent the entry of the bacteria (132). It can be hypothesized that the apoplastic ROS formation acts as an overriding mechanism that rapidly closes the stomatal pore.
OST1 can phosphorylate AtRBOHF in vitro, and these two proteins interact in vivo, suggesting that NADPH oxidase could be an in vivo substrate of OST1 (111). It is of interest that OST1 kinase has also been shown to phosphorylate and thereby activate SLOW ANION CHANNEL 1 (SLAC1), a guard cell anion channel that is a central regulator of stomatal responses triggered by a number of stimuli, including ozone and ABA (11, 33, 62, 123, 124). OST1 kinase is therefore emerging as a component linking NADPH oxidase activation and stomatal function.
The guard cell model system has allowed the elucidation of the sequence of the events presented in Figure 7, which is applicable to both hormonal and stress regulation: when guard cells perceive the presence of bacteria, the stress hormone ABA, or other relevant stresses, signals activate the plasma membrane NADPH oxidases AtRBOHD and/or AtRBOHF producing an apoplastic ROS burst, which is either sensed apoplastically, or the resulting H2O2 is transported through the plasma membrane trough H2O2-specific aquaporins. At the same time Ca2+-fluxes trough plasma membrane are activated, followed by Ca2+-sensing at the cytoplasm and subsequent transfer of the signal to the chloroplast. This coincides with the activation of anion fluxes through the SLAC1, followed by initiation of rapid stomatal closure and ROS production in the chloroplast, which is apparently involved in the regulatory processes. This gives a frame for the studies in ROS and redox regulation of stress acclimation also for the photosynthetic mesophyll cells.
Current Status of Research on Apoplastic and Chloroplastic Redox Signaling Networks
Extensive research efforts have started to reveal the nature of apoplastic and chloroplastic redox signaling networks in plants. Also, the sequence of events, starting from stress perception, proceeding through activation of apoplastic ROS burst by NADPH oxidases, cytoplasmic and chloroplastic Ca2+-transients, and organellar redox signals, and finally culminating in physiological responses is starting to emerge. Moreover, the vast functional overlap between light acclimation and plant immunity governed by cross-communication between organellar and apoplastic compartments has become generally accepted.
Although it is clear that external signals are first sensed in the apoplast, great majority of the sensory systems and the biological functions of plasma membrane RLK remain poorly understood. Activation of NADPH oxidases upon stress perception in turn has become common currency in plant signaling research. Even so, regulation of NADPH oxidase activity by calcium and reversible phosphorylation, identity of the underlying protein kinases and phosphatases, and the mechanisms by which apoplastic redox changes become translated into cellular signaling events remain to be established.
Organellar signaling and metabolic crosstalk have also proven central for environmental sensing. The complex metabolic interactions connecting cellular energy status with redox metabolism, however, are far from being well understood. The mechanisms for responding to a wide range of redox imbalances in chloroplasts have long been matters of intensive research, and the use of genetic approach has led to identification of key signaling components that relay information from chloroplasts to the nucleus. Their modes of action, however, are not yet well understood. More recently, studies have shown that besides abiotic stress responses, organelles determine immune reactions against biotic stress agents. How these signaling mechanisms connect with photosynthetic electron transfer reactions is drawing increasing attention in the field.
Besides eliciting local responses, chloroplasts and apoplast cross-communicate to initiate systemic signals that spread through the vasculature and confer stress resistance in shaded parts of the plant. In addition, signals from apoplastic ROS production are relayed to chloroplasts upon activation of plant immune signaling. In stomatal guard cells, the sensing of apoplastic ROS is also followed by chloroplastic ROS production. However, details of the signaling involved in transmitting the apoplastic ROS signal to the chloroplasts are completely unknown. As an experimental single cell system, guard cells can be used as a model to explore these signaling interactions in more detail.
Major gaps still remain in our understanding of the entire redox signaling network where signaling continuums between different cellular compartments ultimately determine the outcome of acclimation responses. Among major challenges, identification of the sensory mechanism for ROS and the components involved in relaying the signals from the apoplast to the chloroplast are awaiting discovery.
Future Perspectives
The emerging picture on redox/ROS signaling indicates a complex interplay among apoplastic and chloroplastic redox signaling and hormonal regulation in plants. Considerable efforts are, however, still needed before a number of key open questions concerning the perception and relay of environmental stresses can be answered. Major breakthroughs can be expected in identification of sensory systems that have the capability to make a distinction between types of ROS. Also, the mechanisms of and components involved in signal transduction between the apoplast and chloroplasts are awaiting discovery. A comprehensive view of regulatory networks will facilitate the understanding on how environmental factors affect the production of phytonutrients and biomass in plants. Translation of such information from model plants to crop species will be on the cutting edge of research in the near future.
Footnotes
Acknowledgments
This work was financially supported by the Academy of Finland projects 250336 to J.K., 263772, 218157, and 130595 to S.K., and by the Finnish Funding Agency for Technology and Innovation, project 424/31/2011 to J.K. Grzegorz Konert is acknowledged for sharing graphics for illustrations.