
Abstract
Introduction
We have recently demonstrated that HMOX1 may act in a similar manner to oncogene and increase the risk of hyperplastic growth of myogenic precursors through the regulation of microRNAs (miRNAs) (20). miRNAs are a class of small noncoding RNAs, which can very potently regulate the expression of genes and translation of proteins, interfering with ribosomal machinery. They commonly target 3′ untranslated regions (3′UTRs) of mRNAs, decreasing their stability and suppressing translation. However, they can also activate other genes (50). MiRNAs can affect various biological processes, such as proliferation, migration, and angiogenesis, which may lead to tumor progression. Down-regulation of miRNAs is a common feature of human malignancies (32). Aberrant expression of miRNAs may contribute to tumorigenesis by inhibiting the expression of tumor suppressor genes or by promoting the expression of proto-oncogenes and angiomirs. Accordingly, it has been suggested that miRNA expression profile in lung cancer can help predict survival of NSCLC patients (23).
An interesting example of the known oncomir and angiomir is miR-378. hsa-miR-378 precursor gives rise to two mature strands: hsa-miR-378 (hsa-miR-378a-3p) and hsa-miR-378* (hsa-miR-378a-5p). miR-378 promotes cell survival and tumor growth of glioblastoma by targeting two tumor suppressors: Sufu and Fus-1 (24). Its pro-tumorigenic and pro-angiogenic role was also demonstrated in A549 adenocarcinoma cells (4). miR-378 was also identified as a novel target of the c-Myc oncoprotein that is able to cooperate with activated Ras or human epidermal growth factor receptor 2 (HER2) in tumor development (9). Furthermore, miR-378* can induce molecular switch that is involved in the orchestration of the Warburg effect in breast cancer cells (8).
Based on the previously demonstrated effects of HMOX1, we hypothesized that it can influence human NSCLC growth, metastasis, and angiogenesis through modulation of miRNAs. Surprisingly, both in vitro and in vivo in an animal model, HMOX1 attenuated tumor cells proliferation and migration and decreased tumor growth, significantly affecting miRNA pathway. Interestingly, reciprocal interplay between HMOX1 and oncomir miR-378 influenced NSCLC in opposite ways. This interaction may be of significance for the tumor growth, angiogenesis, and metastasis.
Rebound Track
This work was rejected during standard peer review and rescued by Rebound Peer Review (Antioxid Redox Signal 16: 293–296, 2012) with the following serving as open reviewers: James F. George, Mahin D. Maines, Justin C. Mason, and Yasufumi Sato. Comments by these reviewers supporting the rescue are listed next:
Results
HO-1 modulates miRNA transcriptome in NCI-H292 NSCLC
To assess the effect of HO-1 on tumorigenic and angiogenic capabilities of NSCLC, we generated NCI-H292 cells stably overexpressing HMOX1 after transfection with plasmid (NCI-pcDNA-HO-1) and transduction with retroviral vectors (NCI-HO-1). NCI-H292 cells modified with pcDNA3.1 plasmid vector (NCI-pcDNA) and retroviral empty vector (NCI-EV-ctrl) served as controls. The main advantage of NCI-H292 cell line is its low basal level of HMOX1 in comparison to another commonly used model: A549 cells (Supplementary Fig. S1A; Supplementary Data are available online at
First, we demonstrated that HMOX1 overexpression slightly, but significantly, enhanced the expression of DiGeorge syndrome critical region-8 (DGCR8), Drosha, and Dicer1 mRNA (Fig. 1A–C), the genes that are responsible for miRNA processing. Accordingly, Drosha and DGCR8 proteins were elevated in HMOX1 overexpressing cells and in control cells with pharmacologically increased HMOX1 level by CoPPIX treatment (Supplementary Fig. S2A–C). Concomitantly, a global amount of small RNA in range 19–25 nt (miRNA) was increased in comparison to all <250 nt small RNA (Fig. 1D) and total RNA (Fig. 1E); whereas the amount of <250 nt small RNA compared with total RNA was unaffected (Fig. 1F).

Subsequently, miRCURY™ LNA array analysis revealed that elevated HMOX1 level significantly modulated the expression of miRNAs that were potentially involved in tumor growth and vascularization. Expression of known oncomirs (members of miR-17–92 cluster, miR-20b, miR-135a, miR-210, and miR-378) and angiomirs (miR-17–92, miR-210, and miR-378) was down-regulated; whereas several tumor-suppressive miRNAs (miR-22, miR-24, miR-129-3p, miR-130a, miR-181a, miR-193b, miR-424, and miR-1246) and anti-angiomirs (miR-424) were up-regulated (Fig. 1G, H). The most potently down-regulated was miR-378; so, we selected it for further investigations. Moreover, with qPCR, we also confirmed the expression pattern of the selected miRNAs involved in tumor growth and angiogenesis in the second stable model, with lower HMOX1 overexpression level after transfection with pcDNA3.1 plasmid vectors (Supplementary Fig. S3). Importantly, miR-378 was also down-regulated in cells overexpressing HMOX1 after plasmid transfection (Supplementary Fig. S3).
HMOX1 diminishes and miR-378 enhances proliferation, migration, and angiogenic capabilities of NCI-H292 cells in vitro
Since miRNAs expression profile in HMOX1 overexpressing NCI-H292 cells suggested diminished tumorigenic and angiogenic capabilities, we investigated them further in vitro. HMOX1 overexpression increased the number of cells with a high level of p53, a known tumor suppressor (Supplementary Fig. S4A, B). Moreover, we observed a transition of cell cycle to G1 phase (Supplementary Fig. S4C), which was concomitant with a significant attenuation in proliferation of HMOX1 overexpressing cells (Fig. 2A), as well as we demonstrated a significant decrease in migration that was estimated as an inhibition of cell motility in scratch assay (Fig. 2B). Furthermore, the effects of HMOX1 on the proliferation of NCI-H292 cells were mimicked by carbon monoxide, one of HMOX1 activity products (Fig. 2A, B). Cellular parameters were also verified in NCI-H292 cells with a stable lower HMOX1 overexpression level after transfection with pcDNA3.1 plasmid vectors. We again observed a slight, but significant attenuation in proliferation (Fig. 2C) and a significant decrease in cell migration (Fig. 2D). Accordingly, HMOX1 silencing with siRNA (Supplementary Fig. S5A, B) enhanced proliferation (Fig. 2E) and migration (Fig. 2F) in both control and HMOX1 overexpressing cells.

We suspected that miR-378 might be one of the crucial mediators of the observed effects. Interplay between HMOX1 and miR-378 was then demonstrated in different NSCLC cell lines: SK-MES-1, A549, NCI-H460, and NCI-H292 and cell lines with a higher HMOX1 basal level displayed lower miR-378 expression (Fig. 3A). Moreover, silencing of HMOX1 with siRNA (Supplementary Fig. 5C) up-regulated or showed a tendency to up-regulate miR-378 level in those cell lines (Fig. 3B).
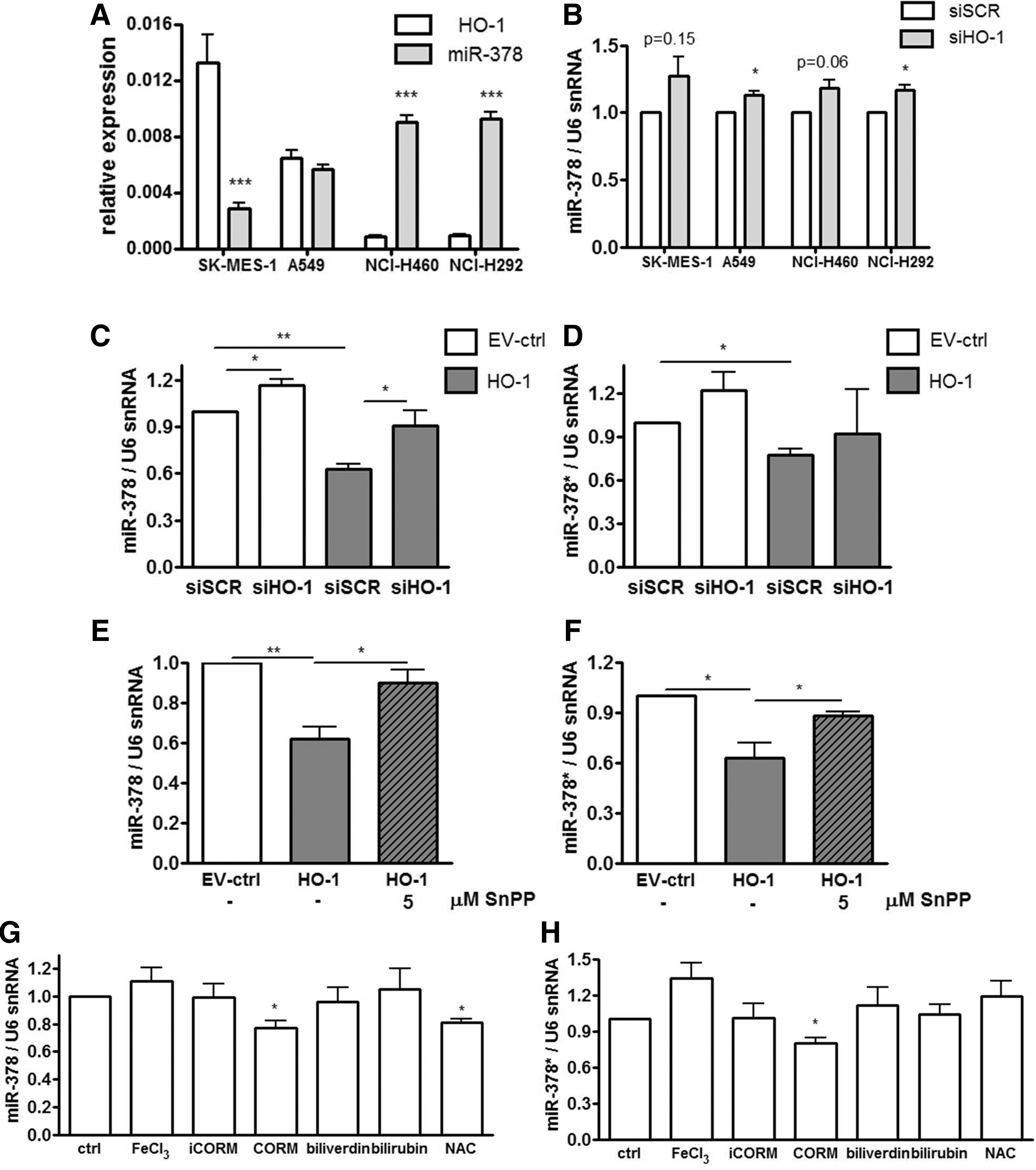
Of note, miR-378 (hsa-miR-378a-3p) is expressed along with the second strand miR-378* (has-miR-378a-5p), which was not detected in microarrays, but with real-time PCR we managed to evaluate its expression (Fig. 3C–H). Silencing of HMOX1 with siRNA in NCI-H292 cells partially but significantly reversed the influence of HMOX1 on miR-378 expression (Fig. 3C) and seemed to affect miR-378* (Fig. 3D). Importantly, the pharmacological inhibition of HMOX1 with SnPPIX also reversed the effects of HMOX1 on miR-378 (Fig. 3E) and miR-378* (Fig. 3F). To determine which product of HMOX1 activity might be responsible for miR-378 and miR-378* down-regulation, we treated NCI-H292 cells with FeCl3, CO-releasing molecule (CORM), inactive CORM (iCORM, negative control), biliverdin, bilirubin, and with a known reactive oxygen species (ROS) scavenger: N-acetyl-L-cysteine (NAC). It turned out that the effect of HMOX1 on miR-378/378* level can be mimicked by only one of its products: carbon monoxide (Fig. 3G, H). Furthermore, NAC down-regulated miR-378 (Fig. 3G), but not miR-378* level (Fig. 3H). Interestingly, restoration of miR-378 level by transfection with miRNA precursors in HMOX1 overexpressing cells did not reverse HMOX1 effects on proliferation (Supplementary Fig. S5D) and migration (Supplementary Fig. S5E), which was presumably due to the high expression of HMOX1. However, control NCI-H292 cells transfected with pre-miR-378 tended to behave in an opposite manner to HMOX1 overexpressing cells.
The data indicated that higher expression of miR-378 may be required to overcome the effect of HMOX1. Accordingly, stable miR-378 precursor overexpressing cells were obtained using lentiviral vectors with feline immunodeficiency viral vector (FIV) or human immunodeficiency viral vector (HIV) backbones. Control cell lines were transduced with vectors harboring a scrambled sequence. Transduction resulted in overexpression of miR-378 and miR-378* (Supplementary Fig. S6A–D). Accordingly, and as expected, mRNA expression levels of known miR-378 targets, namely Sufu, Fus-1 (24) and cytochrome P450 2E1 (CYP2E1) (39), were decreased in miR-378/378* overexpressing cells (Supplementary Fig. S6E–G).
Interestingly, interplay between HMOX1 and miR-378 was evidenced, as miR-378 precursor overexpression down-regulated HMOX1 mRNA and protein (Fig. 4A and Supplementary Fig. S6H). miR-378 also reduced luciferase activity after transfection with constructs harboring Renilla luciferase with 3′UTR of HMOX1 (pSL-3′UTR-HO-1) and TUSC2 (pSL-3′UTR-Fus1) in comparison to empty constructs with luciferase (pSL-empty), suggesting that miR-378 may directly bind to 3′UTR region of HMOX1 mRNA (Fig. 4B). miR-378 overexpression also decreased the number of NCI-H292 cells with high p53 level (Supplementary Fig. S7A, B), diminished the percentage of cells in G1 phase, and showed a tendency to increase the percentage of cells in S and G2/M phases (Supplementary Fig. S7C), displaying enhanced proliferation (Fig. 4C) and migration (Fig. 4D). The effects were opposite to those observed in HMOX1 overexpressing cells (Fig. 2A–D). Accordingly, blocking of miR-378 with anti-miR-378 reversed the effects of miR-378 overexpression on proliferation (Fig. 4E) and partially on migration (Fig. 4F). To determine whether HMOX1 can reverse the effect of miR-378 on proliferation and migration of lung cancer, NCI-H292 cells overexpressing miR-378 were transduced with adenoviral vectors harboring HMOX1 (AdHO-1) or GFP (AdGFP) as a control. Temporal HMOX1 overexpression with adenoviral vectors tended to reverse the effect of miR-378 on proliferation (Fig. 4G) and restored migration (Fig. 4H).

Since HMOX1 and miR-378 were previously demonstrated to influence angiogenesis, we first estimated the production of pro-angiogenic factors. stromal cell-derived factor-1 (SDF-1) was expressed at the limit of detection. HMOX1 overexpression did not significantly modify vascular endothelial growth factor (VEGF) and interleukin-8 (IL-8) secretion (Supplementary Fig. S8A), whereas stable miR-378 overexpression significantly up-regulated both of them (Supplementary Fig. S8B). Subsequently, expression of other pro-angiogenic and pro-oncogenic mediators was evaluated with real-time PCR. It revealed that angiopoietin-1 (Ang-1) and mucin-5AC (MUC5AC) were down-regulated by HMOX1 (Fig. 5A); whereas Ang-1, Ang-2, and MUC5AC were up-regulated by miR-378 (Fig. 5B).
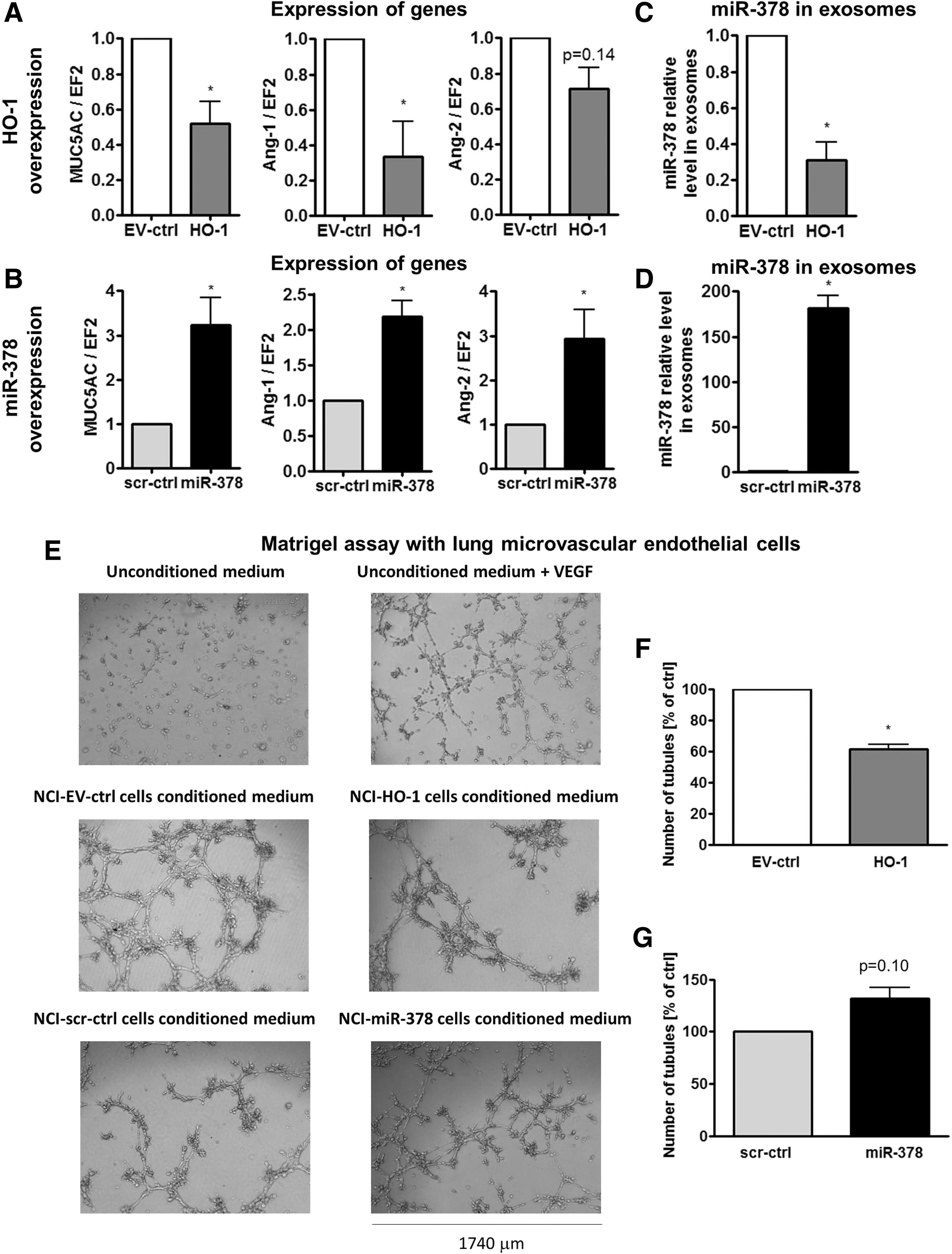
It has been suggested that miRNA can be transferred between cells. Accordingly, here we demonstrate that angiomir miR-378 is exported from NCI-H292 cells in exosomes, and HMOX1 could down-regulate not only expression of miR-378, but also its exosomal export (Fig. 5C). Inversely, cells overexpressing miR-378 secreted more miR-378 in exosomes (Fig. 5D). Therefore, we wanted to determine whether the secretion of miR-378 can affect the endothelial cells (EC). Indeed, lung microvascular EC treated with conditioned media harvested from NCI-H292 cells proliferated more potently than those cultured with unconditioned media, evidencing pro-angiogenic properties of the cancer cells. NCI-HO-1 conditioned media slightly, but significantly, inhibited proliferation of EC (Supplementary Fig. S8C); however, NCI-miR-378-conditioned media did not exert significant effects (Supplementary Fig. S8D). Furthermore, EC treated with media harvested from NCI-HO-1 formed less tubule-like structures on Matrigel (Fig. 5E, F); whereas the effect of media on NCI-miR-378 tended to be opposite (Fig. 5E, G). The effect of conditioned media harvested from NCI-H292 cells on the formation of tubule-like structures was also confirmed using human umbilical vein endothelial cells (Supplementary Fig. S8E–G).
HMOX1 diminishes and miR-378 enhances NSCLC growth, vascularization, oxygenation, and distal metastasis in vivo
For in vivo experiments we utilized NCI-H292-Luc cells in subcutaneous xenografts in a Swiss nude immunodeficient murine model. We confirmed the interplay between HMOX1 and miR-378/miR-378* in vivo, showing the mutual regulation of the expression of each other (Fig. 6A, B and Supplementary Fig. S9A–D). HMOX1 overexpression significantly inhibited NCI-H292 xenograft growth (Fig. 6C). On the contrary, miR-378 overexpression enhanced tumor growth (Fig. 6D). Measurements of luciferase activity by IVIS showed slightly diminished total flux signal in tumors overexpressing HMOX1 at 28 days after implantation (Fig. 6E) and enhanced luminescence in tumors overexpressing miR-378 starting from 14 day (Fig. 6F), confirming tumor growth estimated with caliper (Fig. 6C, D). Attenuated proliferation of cells overexpressing HMOX1 and enhanced proliferation of cells overexpressing miR-378 was demonstrated by proliferating cell nuclear antigen (PCNA) staining in tumor cryosections (Fig. 6G, H, see also Supplementary Fig. S9E, F). Similarly, plasmid-transfected cells overexpressing HMOX1 at a lower level were utilized in the xenograft NOD-SCID murine model. HMOX1 overexpression again resulted in significant inhibition of tumor growth rate (Supplementary Fig. S9G).

Since the tumor growth is highly dependent on angiogenesis, we determined the effect of transgenes in NCI-H292 cells on tissue vascularization. Nodules formed by HMOX1 overexpressing cells were smaller, and no eye-visible necrosis was detected (Supplementary Fig. S10A). On the contrary, tumors overexpressing miR-378 were bigger with traces of edema and were better vascularized (Supplementary Fig. S10B). Staining for cluster of differentiation 31 (CD31) revealed the presence of vessels inside the nodules and indicated the lower number of capillaries in tumors overexpressing HMOX1 (Fig. 7A and Supplementary Fig. S10C), but higher in tumors overexpressing miR-378 (Fig. 7B and Supplementary Fig. S10D). Concomitantly, oxygen partial pressure, estimated in vivo near the tumor surface by ruthenium fluorescence quenching measurement, was diminished in tumors overexpressing HMOX1 (Fig. 7C) and enhanced in tumors overexpressing miR-378 (Fig. 7D). Moreover, mice with xenografts overexpressing HMOX1 tended to have diminished miR-378 level in blood-circulating exosomes (Supplementary Fig. S10E), whereas this tended to increase in the case of miR-378 overexpression (Supplementary Fig. S10F), supporting the results from in vitro measurements (Fig. 5C, D). Then, we evaluated the modulation of genes regulating angiogenesis and metastasis. SDF-1 protein in tumors was down-regulated by HMOX1 (Fig. 7E) and up-regulated by miR-378 (Fig. 7F). Moreover, the high level of HMOX1 led to a decrease in MUC5AC, Ang-1, and matrix metalloproteinase 12 (MMP12) and an increase in tumor necrosis factor-alpha (TNF-α) and interleukin-1beta (IL-1β) (Fig. 7G and Supplementary Fig. S10G). Opposite effects were visible in tumors overexpressing miR-378 (Fig. 7H and Supplementary Fig. S10H).

Metastatic potential of tumor cells was measured as an amount of human ubiquitin c transcripts in different organs of the host. HMOX1 showed a tendency to diminish distal metastasis to the brain (Fig. 8A). On the contrary, miR-378 enhanced distal metastasis to the lungs, seemed to nonsignificantly favor metastasis to the brain, and did not affect the local, right-limb bone marrow (Fig. 8B). No metastasis was detected in left-limb bone marrow, liver, and adrenal glands (Fig. 8A, B). Ex-vivo bioluminescence imaging allowed for detecting the luciferase signal in the excised right inguinal lymph nodes, the closest nodes to the xenograft implantation, in all experimental groups (Supplementary Fig. S11A, B). Number of platelets, modulating tumor angiogenesis and metastasis, was decreased in mice-bearing tumors overexpressing HMOX1 (Fig. 8C) and increased in case of miR-378 overexpression (Fig. 8D) compared with mice with control tumors and healthy animals.
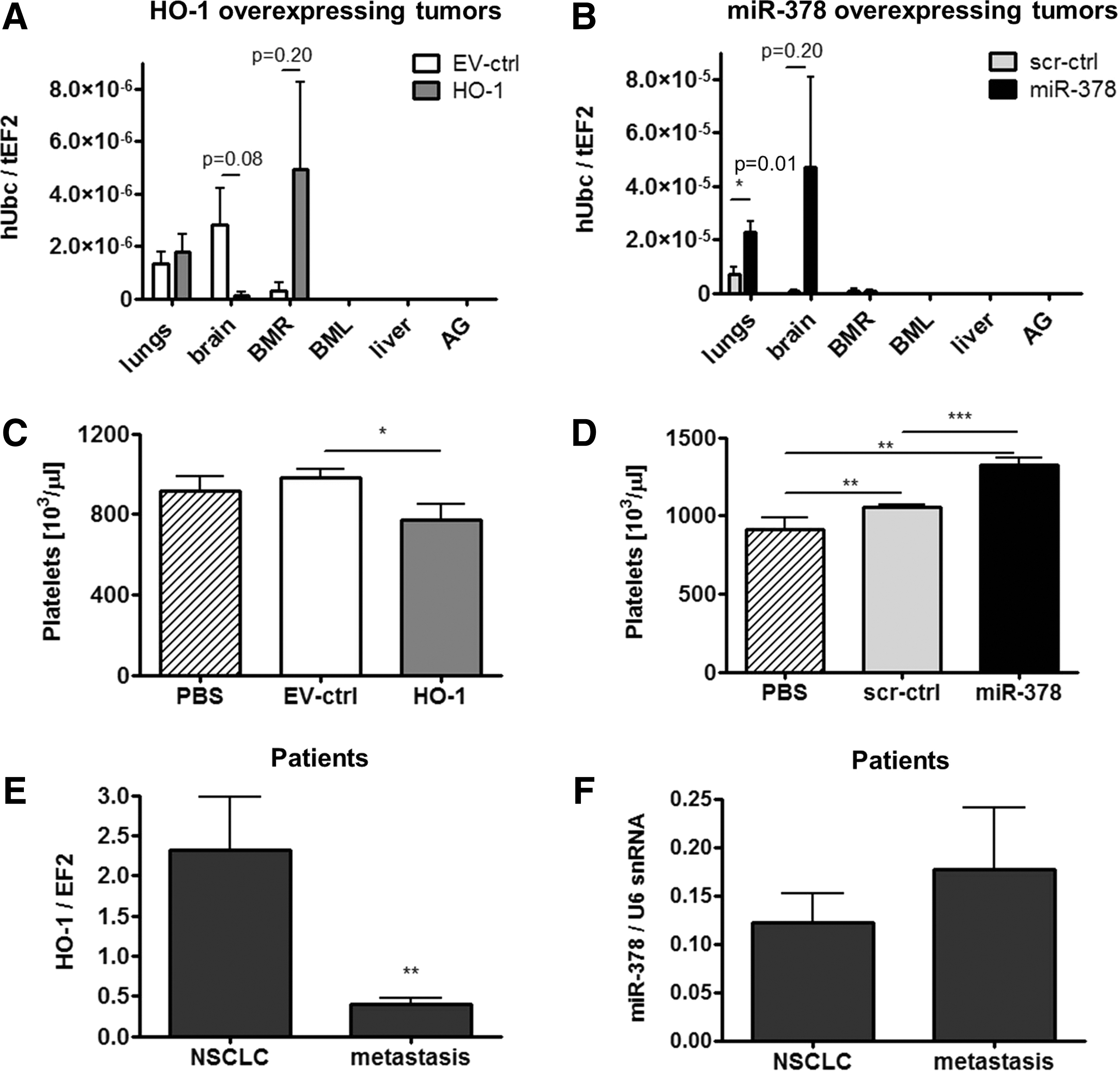
To extrapolate results from mice to patients suffering from NSCLC, we evaluated the expression of HMOX1 and miR-378 in primary NSCLC tumors and in metastases to lymph nodes. Quantitative real-time PCR revealed a higher HMOX1 levels in primary NSCLC than in metastases (Fig. 8E), whereas miR-378 expression seemed to be the opposite (Fig. 8F).
HMOX1 diminishes while miR-378 does not modulate resistance of cells to chemotherapeutics and oxidative stress
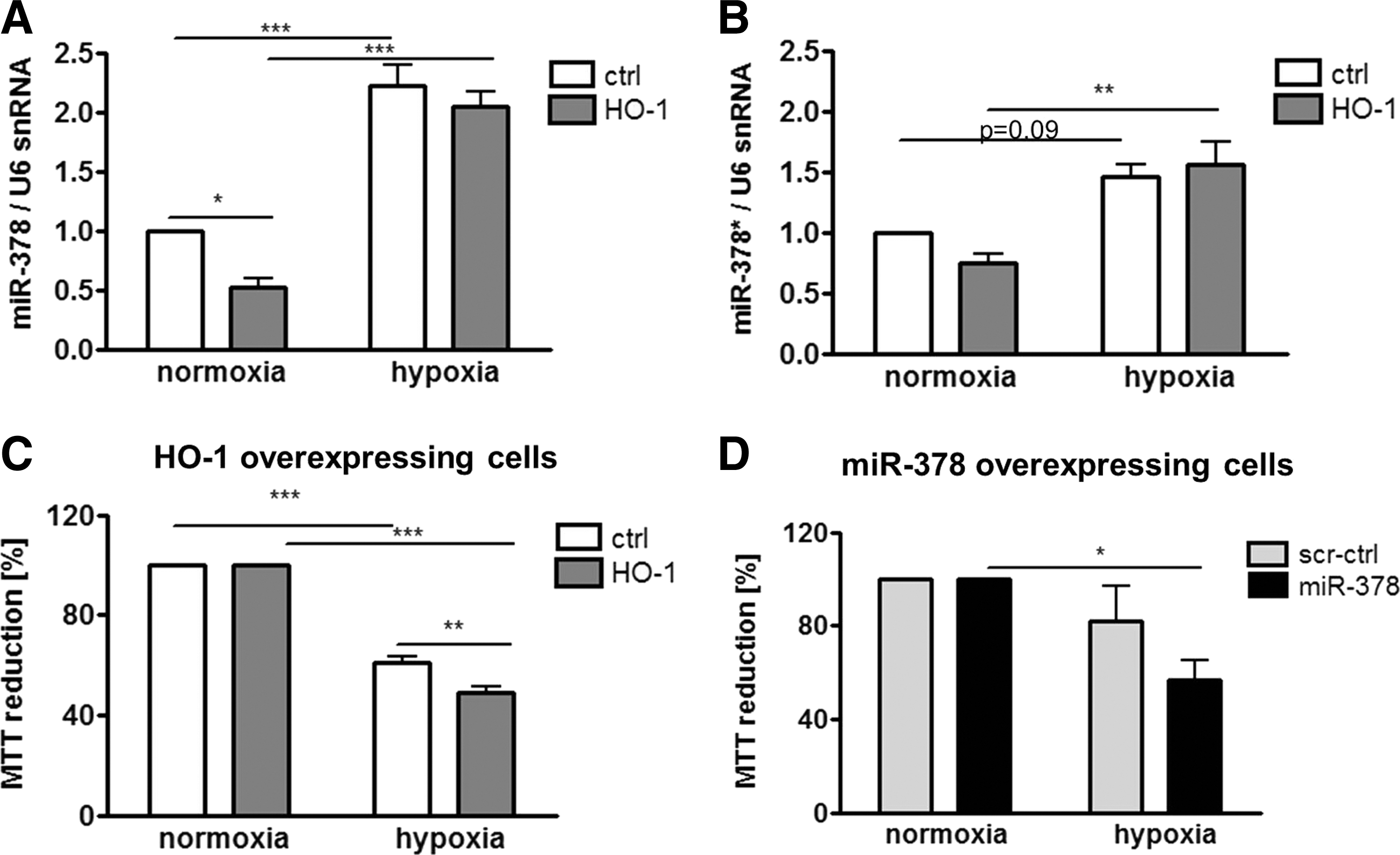
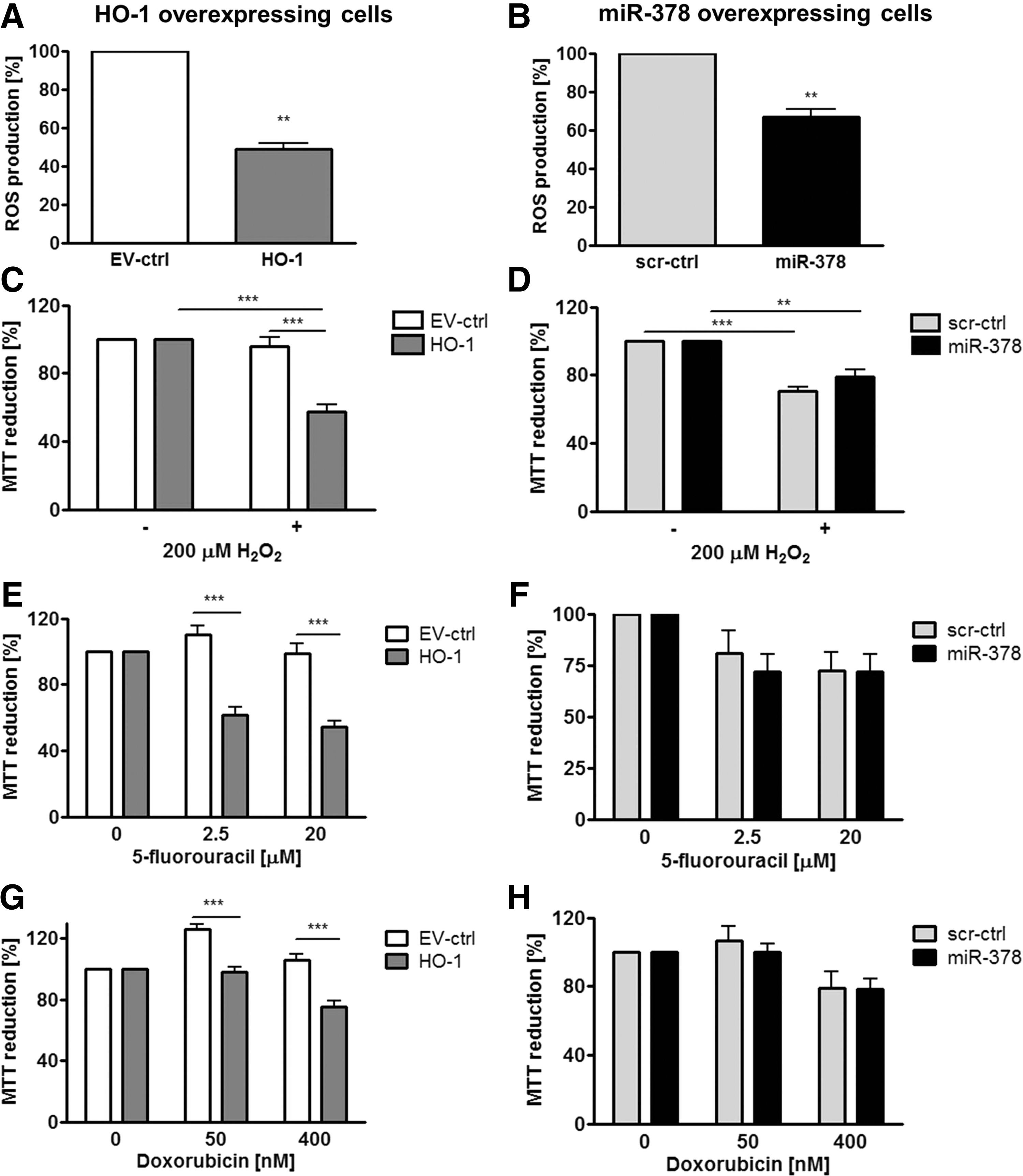
In many studies, HMOX1 has been considered a cytoprotective enzyme, which is up-regulated in stressful conditions and protects cells against oxidative stress injury (53). Therefore, we evaluated the role of HMOX1 and miR-378 in the resistance of cancer cells to stress. First, we have shown that hypoxia up-regulated miR-378 and miR-378* in control and HMOX1 overexpressing cell lines (Fig. 9A, B). Nevertheless, in hypoxia, both HMOX1 and miR-378 overexpressing cells displayed reduced viability (Fig. 9C, D). Interestingly, both HMOX1 and miR-378 displayed diminished production of ROS (Fig. 10A, B). However, viability of HMOX1 overexpressing cells was diminished after treatment with H2O2, doxorubicin, and 5-fluorouracil; whereas miR-378 overexpression did not modify it significantly (Fig. 10C–H).
Discussion
The salient finding of the present study is the demonstration of mutual interactions between HMOX1 and miR-378 in NSCLC cells in which a high level of HMOX1 correlates with a low expression of mirR-378. Investigating the mechanisms of that interplay in a model of NCI-H292 cell line, with low basal HMOX1 expression, we showed that HMOX1 overexpressing NCI-H292 cells displayed reduced proliferation and migration. Furthermore, growth of the HMOX1-overexpressing cells in Swiss nude mice was impaired, tumors were less vascularized and revealed decreased expression of SDF-1 and Ang-1. Distal metastasis to the brain tended to be attenuated by HMOX1 overexpression and was accompanied by down-regulation of MMP12 and MUC5AC and up-regulation of TNF-α and IL-1β. Interestingly, HMOX1 overexpression in NCI-H292 cells increased total pool of miRNAs with some suppressor miRNAs up-regulated and oncogenic ones decreased. The most strongly inhibited was miR-378, a known oncomir, with pro-angiogenic properties. Further studies revealed that carbon monoxide, one of HMOX1 by-products, significantly decreased miR-378 expression, and the effect was mimicked by NAC, indicating the redox-dependent regulation. On the other hand, when miR-378 was overexpressed by lentiviral vectors transduction in NCI-H292 cells, it inhibited the expression of its known target genes, such as Sufu, Fus-1, and CYP2E1 and also targeted mRNA of HMOX1. Overexpression of miR-378 affected NSCLC properties in an opposite manner to HMOX1: proliferation and migration of tumor cells were increased, tumor growth was accelerated, with augmented tumor vascularization and oxygenation, distal metastasis to lungs appeared to be increased, and angiogenic or inflammatory gene expressions were regulated by miR-378 inversely than in HMOX1 overexpressing cells.
Both this and our other recent study (20) indicate that HMOX1 is a potent regulator of miRNAs biogenesis and function, acting in a cell-type specific manner. Indeed, certain miRNAs that are up-regulated by HMOX1 in NCI-H292 cells were previously described as tumor suppressors inhibiting either proliferation or migration of cancer cells. This group consists of miR-181a—tumor suppressor in NSCLC (11); miR-193b—inhibitor of invasion of breast cancer (29); miR-424, which arrests cancer cells in G1 phase (43) and inhibits angiogenesis (3); miR-22—inhibitor of invasion of ovarian cancer (28); miR-24—inhibitor of cancer cell proliferation (38); miR-129-3p whose low expression was associated with poor clinicopathological features of primary gastric cancer patients (48); miR-130a—inhibitor of migration of NSCLC cells (1); and miR-1246, which can induce apoptosis (57).
Accordingly, the expression of known oncomirs was down-regulated by HMOX1. For example, miR-378 promoted glioma growth (24) and NSCLC invasion (4). Members of miR-17–92 cluster, such as miR-17*, −18a, −18b, −19b, and −20a, were previously associated with carcinogenesis and were up-regulated in tumors, including lung cancer (17). miR-210 and miR-135a also augmented tumor growth and metastasis (31, 41), while miR-20b displayed an oncogenic role in normoxia (27). Similarly, angiomirs, miR-17–92 cluster, miR-378, miR-210, and miR-135, were down-regulated (7, 24, 30, 35).
In humans, HMOX1 promoter displays (GT)n repeat polymorphism, and short S allele leads to a higher basal HMOX1 level (45). Longer (GT)n repeats may be associated with the development of lung adenocarcinoma in Japanese male smokers (19). This suggests that lower HMOX1 expression may favor lung cancer development, which fits our results. Similarly, HMOX1 also inhibited the development of breast cancer (14) and tongue SCC (55), but it promoted the growth of melanoma (52), pancreatic cancer (44), and sarcoma (34).
We next hypothesized that miR-378, as the oncomir and angiomir being most potently down-regulated by HMOX1 in comparison to other miRNAs, might mediate HMOX1 effects in NCI-H292. Transient restoration of its level did not reverse the effects of HMOX1 overexpression in NCI-H292 cells, probably because HMOX1 regulates tumor growth through many miRNAs. However, we observed that both temporal and stable miR-378 overexpression stimulates proliferation and migration in NCI-H292 cells. Similarly, miR-378 promoted cell survival and tumor growth of glioblastoma by targeting Sufu and Fus-1 (24); facilitated the c-Myc-mediated transformation of breast cancer (9); and favored the NSCLC invasion to the brain (4). Regulation of Sufu and Fus-1, as well as a tendency to augmented brain metastasis, was also observed in our studies. Moreover, miR-378* was identified as a molecular switch involved in the orchestration of the Warburg effect in human breast cancer (8).
HMOX1 was not predicted to be a typical miR-378 target with bioinformatics tools. However, in our analysis, one of the methods called RNA22 (37) predicted such probability, what was supported by experiments, where HMOX1 was down-regulated in response to miR-378 overexpression. Alternatively miR-378 may regulate HMOX1 indirectly. For example, CYP2E1, a direct target of miR-378 (39), may influence HMOX1 expression (46). In our study, CYP2E1 was also down-regulated in miR-378 overexpressing cells.
Importantly, miR-378 overexpressing cells with down-regulated HMOX1 behaved oppositely to HMOX1 overexpressing cells with down-regulated miR-378. Enhanced proliferation of NCI-miR-378 cells and diminished proliferation of NCI-HO-1 cells in vitro may translate for mode of tumor progression in vivo. Tumor growth rate may be also explained by transition of cell cycle from G1 to S and G2/M phase or partial growth arrest in G1 phase in NCI-miR-378 or NCI-HO-1, respectively. Furthermore, HMOX1 silencing with siRNA and inhibition of miR-378 with anti-miR reversed the effect of HMOX1 and miR-378 on proliferation of cancer cells. Similarly, A549, another adenocarcinoma cell line, exhibited growth arrest and increased cells' number in G0/G1 phase when overexpressing HMOX1 (26); whereas A549 cells overexpressing miR-378 proliferated faster (4). Diminished proliferation of cancer cells with elevated HMOX1 expression was also demonstrated for breast (13) and prostate cancer (14).
In our hands, HMOX1 overexpression tended to attenuate distal metastasis, while miR-378 enhanced lung metastasis. These observations are supported by an analysis of NSCLC clinical samples in which HMOX1 expression was lower in lymph node metastasis than in primary tumors, while miR-378 tended to be nonsignificantly higher in metastasis. Other studies also demonstrated the association of miR-378 with brain metastasis and enhanced migratory capabilities of A549 adenocarcinoma cells overexpressing miR-378 (4). In regulation of metastatic properties, the MMP12 or MUC5AC, differently regulated by HMOX1 and miR-378 in vivo, may be involved. MMP12 expression correlated with local recurrence and metastatic disease in NSCLC patients (15), and MUC5 overexpression was associated with early postoperative metastasis in NSCLC (56). Platelets may enhance NSCLC metastasis (21) and here, tumors overexpressing miR-378 were also associated with increased number of platelets in blood, whereas HMOX1 exerted the opposite effect.
The metastatic potential is affected by tumor vascularization. In many studies, HMOX1 has been shown as a pro-angiogenic enzyme up-regulating VEGF (18). There is only one study demonstrating the anti-angiogenic effect of HMOX1 in prostate cancer (10). Here, we demonstrated diminished angiogenic potential of lung cancer cells overexpressing HMOX1. However, no direct relation with VEGF or IL-8 level was established, although Ang-1 and SDF-1 were inhibited by HMOX1. In contrast, miR-378 overexpression increased VEGF, IL-8, Ang-1, and SDF-1, indicating the role of those factors in miR-378-enhanced angiogenesis in NSCLC. miR-378 can promote VEGF expression by competing with miR-125a for the same seed region in the VEGF 3′-UTR (16) or through Sufu–sonic hedgehog homolog (Shh) pathway (51).
Importantly, enhancement of tumor growth by miR-378 and its inhibition by HMOX1 could also partially result from the regulation of tumor suppressors, such as p53 (22). Finally, we demonstrated that HMOX1 overexpressing cells were more sensitive to hypoxia, oxidative stress caused by H2O2, and chemotherapeutics: 5-fluorouracil or doxorubicin. The potentially puzzling higher sensitivity of HMOX1 overexpressing cells can be explained by other studies indicating that higher levels of HMOX1 can be associated with significant oxygen cytotoxicity, due to accumulation of reactive iron (2).
To conclude, we demonstrated for the first time that HMOX1 overexpression inhibits tumorigenic and angiogenic capabilities of human NCI-H292 NSCLC cells. Mechanistically, this involves the regulation of miRNAs. Of particular significance, there is an interplay between HMOX1 and miR-378, which is reflected by reciprocal regulation of their expression in several NSCLC cell lines and by the opposite effects on lung cancer growth, vascularization, oxygenation, and progression (Supplementary Fig. S12). Importantly, the described cross-talk may be of clinical significance, as evidenced by the lower expression of HMOX1 and the tendency of higher miR-378 in lymph node metastasis in NSCLC patients.
Materials and Methods
Cell line
NSCLC cell lines NCI-H292, NCI-H460, A549, and SK-MES-1 were purchased from the American Type Culture Collection (ATCC). The NCI-H292 cell line purity was authenticated by a short tandem repeat analysis at LGC Standards Ltd company by PowerPlex® 16HS 16 Loci Service. NCI-H292 cells were grown in RPMI 1640 medium (PAA or Lonza) with 10% FBS (PAA), penicillin (100 U/ml), and streptomycin (0.1 mg/ml; Invitrogen). Other cell lines were cultured as described in Supplementary Materials and Methods.
Generation of stable cell line overexpressing HMOX1 and miR-378/378*
Retroviral vectors were produced using Phoenix-Ampho HEK293 packaging cell line with MSCV luciferase PGK-hygro (Addgene), pLNCX2, or pLNCX2-HO-1 plasmids and then, NCI-H292 cells were transduced, as previously described (54), and subsequently selected with antibiotics: G418 (800 μg/ml; Cytogen) for pLNCX2 (NCI-EV-ctrl cells) or pLNCX2-HO-1 (NCI-HO-1 cells) backbones, and hygromycin (Invitrogen; 600 μg/ml) for MSCV luciferase PGK-hygro backbone (NCI-Luc, NCI-Luc-EV-ctrl, and NCI-Luc-HO-1 cells for in vivo experiments).
Lentiviral vectors particles (pEZX-MR01, FIV backbone, or pEZX-MR03, HIV backbone), harboring miR-378 precursor or control scrambled sequence, were purchased from Gene Copoeia. Since in vitro results were analogous for both vector backbones, for subsequent in vivo experiments, only cells transduced with pEZX-MR03 backbone were used, as they maintained higher luciferase activity.
Characterization of cell lines
Details relevant to characterization of cell lines and angiogenic assays are provided in Supplementary Materials and Methods.
Microarray profiling
MicroRNA Array Profiling was performed by Exiqon Company using miRCURY™ LNA array version 11.0. Results of at least a 1.15-fold difference and p≤0.05, estimated in paired t test using MeV program—TM4 Microarray Software Suite (42)—were considered significant. Microarray data are deposited in Gene Expression Omnibus (GEO) under accession number GSE37779.
Animal studies with NCI-H292 cells transduced with viral vectors
All animal work was approved by the Local Ethical Committee for Animal Research in Orleans in France. All manipulations on animals were performed under anesthesia with inhaled isoflurane. 6-week-old Swiss nude females were injected subcutaneously with 5×106 empty vector control (EV-ctrl) or HMOX1 overexpressing cells or scrambled vector control (scr-ctrl) or miR-378/378* overexpressing NCI-H292-Luc cells in PBS. Tumor size was monitored weekly with caliper (V=D×d 2×0.5; V, tumor volume; D, the biggest dimension; d, the smallest dimension) and with IVIS® Lumina II Imaging System (Caliper Life Science) after intraperitoneal injections of luciferin (150 μl, 5 mg/ml). Tumor tissue pO2 was estimated with fiber-optic oxygen-sensing device (OxyLite™; Oxford Optronix). For hematological analysis of blood, 150 μl of noncoagulated blood was taken from the eye using Pasteur pipette to a tube with EDTA. It was analyzed with a digital computer analyzer of hematology MS9-5 V (Melet Schloesing Laboratoires). The animals were sacrificed 5 weeks after xenografting, and tumor, lungs, brain, bone marrow, liver, adrenal glands samples, and blood from the heart were collected. Tissue specimens for staining were frozen in TissueTEK® O.C.T. compound. Cryosections were stained for CD31 and PCNA as previously described (12, 54).
Human studies
Human studies were approved by the Local Ethical Committee of the Collegium Medicum of the Jagiellonian University in Krakow, Poland. Biopsies of primary lung adenocarcinoma and lymph nodes metastases were analyzed.
Statistical analysis
Data are presented as means±SEM of at least three independent experiments. Data were analyzed with unpaired or paired Student's t-test or one-way ANOVA with Tukey's post-test or two-way ANOVA with Bonferroni post-test; p<0.05 was considered significant. Statistical analysis was performed using GraphPad Prism 5 software (trial version).
Innovation
Here, we have shown for the first time that in contrast to its known pro-tumorigenic effect in many cancers, HMOX1 in human NSCLC NCI-H292 cells attenuates cell proliferation, angiogenesis, and metastasis, significantly affecting miRNAs. An oncogenic miR-378 is the most strongly inhibited by HMOX1 and reciprocally, miR-378 overexpression decreases HMOX1 but increases pro-angiogenic genes enhancing tumor growth. In vitro and in vivo data indicate that the interplay between HMOX1 and miR-378 significantly modulates NSCLC progression and angiogenesis.
Footnotes
Acknowledgments
The authors thank Alain Le Pape, Stephanie Lerondel, Marilyne Le Mee, Stephanie Retif, and Julien Sobilo (Center for Small Animals Imaging, CIPA TAAM UPS44, CNRS, Orleans, France) for help with IVIS imaging; David Gosset (Cytometry Platform of CBM UPR4301, CNRS Orleans, France) for help with the flow cytometer; and Jacek Stepniewski from UJ for the amplification of pSL plasmids. This work was supported by grants 347/N-INCA/2008 and N301 144336 from the National Science Center and CNRS-INCA-MSHE Polish-French conv. 2009–2011 and by COST Action TD901 HypoxiaNet. Klaudia Skrzypek and Magdalena Tertil were supported by the Conseil Regional du Centre, stipends for co-tutorial PhD theses. Alicja Jozkowicz was an International Senior Research Fellow from Wellcome Trust. Agnieszka Loboda is supported by the Foundation for Polish Science–Parent-Bridge Program that is co-financed by the European Union within European Regional Development Fund (POMOST/2010-2/8), and she is the recipient of the Ministry of Science and Higher Education Scholarship. The Faculty of Biochemistry, Biophysics, and Biotechnology, Jagiellonian University, received structural funds POIG.02.01.00-12-064/08, 02.02.00-00-014/08, 01.01.02-00-109/09, and 01.01.02-00-069/09.
Author Disclosure Statement
All authors declare that no competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.