
Abstract
Nuclear factor (erythroid-derived 2)-like 2 (Nrf2) modulates gene expression in response to oxidative damage in neurodegenerative diseases, including spinal cord injury (SCI). We noticed that activation of Nrf2 pathway persists for an extended time after clinically relevant contusion model of SCI. Injured Nrf2−/− mice were impaired in hindlimb function, exhibited increased atrophy, demyelination, and astrogliosis of the SC concomitant with altered expression of genes controlling apoptosis, inflammation, and neurotrophic factors suggesting the importance of Nrf2 for recovery. We used lentiviral gene transfer to increase Nrf2 expression and improve functional recovery after SCI. Although the transferred Nrf2 was expressed in neurons and astrocytes, we noticed hindlimb function impairment and elevated expression of pro-inflammatory cytokines as an adverse effect. These toxic effects were not reduced by including Nrf2 in the lentiviral vector. Augmenting the amount of delivered Nrf2 gene diminished toxic effects of the lentivirus, yet was not sufficient to improve functional recovery. Results of this study lead to the hypothesis that Nrf2 plays a crucial and multifaceted role in recovery from SCI, but even high overexpression of Nrf2 in injured SC may not offer extra benefit, providing protection only against lentivirus-induced toxicity that is manifested in the SC. Antioxid. Redox Signal. 20, 1313–1323.
Introduction
T
We observed that nuclear factor (erythroid-derived 2)-like 2 (Nrf2) regulates not only classic target genes, but also genes involved in inflammation, apoptosis, and tissue repair in the clinically most relevant model of spinal cord injury (SCI). We used a novel lentiviral gene transfer approach to increase Nrf2 expression in the injured SC, which resulted in long and stable expression of the transgene in astrocytes and neurons. However, our findings raise the question whether Nrf2 overexpression provides an extra benefit compared to its normal induction in SCI and whether a lentiviral approach is applicable for gene transfer into SC, which appeared to be sensitive to lentivirus even when uninjured.
Nuclear factor (erythroid-derived 2)-like 2 (Nrf2) is the “master regulator” of the antioxidant response, modulating the expression of antioxidant, phase II, and other cytoprotective genes (3). In nonstressed conditions, Nrf2 is constitutively ubiquitinated and thus inhibited through the Keap1/Cullin3 ubiquitin ligase complex. Upon exposure to electrophiles or OS, Keap1 is modified at its regulatory cysteine residues and inactivated. Keap1 inactivation enhances Nrf2 nuclear accumulation and antioxidant response element (ARE)-dependent transcription of genes encoding numerous protective enzymes and scavengers, including hemeoxygenase-1 (HO-1), NAD(P)H:(quinine acceptor) oxidoreductase 1 (NQO1), and glutamate cysteine ligase (GCL) (1). The Nrf2-ARE pathway has been shown to be activated in the SC after mouse compression SCI and rat contusion SCI (6, 9). Moreover, Nrf2 deficiency exacerbates the neurologic deficit and inflammation after compression SCI (6). As OS reflects an imbalance between the oxidants produced by the cells and the antioxidant gene products, a promising alternative would be to induce Nrf2 activation instead of the administration of single antioxidants to combat OS (3). Recently, sulforaphane, a natural compound with various potentially beneficial effects, including activation of Nrf2, was shown to provide protection in SCI (5, 9).
The introduction of transcription factors to the central nervous system with virus vectors enables induction or repression of genes. This is especially important in situations requiring the interplay of several factors sharing a common regulatory pathway. Therefore, in OS conditions the simultaneous induction of several antioxidant genes may be a better approach than transfer of individual antioxidant genes. Lentiviral vectors are commonly used to achieve stable gene expression in vitro and in vivo (8). Recently, we successfully used this approach to deliver human Nrf2 into the mouse hippocampus, a brain area affected in Alzheimer disease (AD). This resulted in an efficient and sustained Nrf2 expression in the hippocampus and improved cognitive functions of transgenic AD mice (4).
In this work, using the most clinically relevant mouse SCI model, we have determined whether Nrf2 in addition to combating OS participates in other pathophysiological mechanisms leading to secondary injury after SCI. Moreover, we have tested whether enhancing Nrf2 expression in the SC by gene transfer leads to improved functional recovery.
Induction of the Nrf2-ARE Pathway After Mouse Contusion SCI
There is some evidence that the Nrf2-ARE pathway becomes activated within the first 24 h after SCI (6, 9). Because it remains unknown how long this activation persists, we studied the time-course expression of the Nrf2 gene (Nfe2l2) and its main target genes Hmox1, Nqo1, Gclc, and Gclm at six different time points after injury. We detected significant elevation of Nfe2l2 expression starting as late as 3 days postinjury (dpi) (p<0.001) with a peak at 7 dpi (Fig. 1A). Hmox1 was significantly upregulated already at 12 h after SCI (p<0.001) and together with Nfe2l2 peaked at 7 dpi, at which time it was induced by 23-fold over controls. Nfe2l2 and Hmox1 elevation persisted for up to 42 dpi (Fig. 1B). Thus, we observed a delayed induction of Nfe2l2 expression peaking not earlier than 1 week after injury and persisting high at least up to 6 weeks when compared to controls. This upregulation of Nfe2l2 was associated with even earlier induction of Hmox1, which showed a sharp peak simultaneously with Nfe2l2 at 1 week, followed by clear reduction at 3 weeks but remaining elevated at least until 6 weeks. The very early induction of Hmox1 at 12 and 24 h may be explained by action of other transcription factors controlling Hmox1. For example, nuclear factor kappa-B (NF-κB) or activating protein-1 both regulate Hmox1 expression and are upregulated by a wide variety of prooxidant and proinflammatory stimuli.

The expression patterns of other Nrf2 target genes were different. Nqo1 is another important Nrf2 inducible gene and a key enzyme in the detoxication of reactive quinines. Although Nqo1 is considered to be one of the most consistently and robustly inducible genes among other cytoporotective enzymes activated through the Nrf2-ARE pathway, in our experiment it was significantly downregulated from 1 dpi (p<0.001) to 3 dpi (p<0.01), and only by 7 dpi reached the level of sham mice. Nqo1 became significantly elevated at 42 dpi (p<0.001) (Fig. 1C). Thus, in our model the pattern of Nqo1 expression was different from previous observations (9) where early upregulation of Nqo1 was demonstrated.
Gclm and Gclc, the two glutamate-cysteine ligase subunits, in contrast started with upregulation and shifted to downregulation several days after the injury. Interestingly, Gclm, but not Gclc was upregulated 24 h after SCI. Starting from 3 to 7 dpi both Gclc and Gclm were significantly downregulated until 21 and 42 dpi, respectively (Fig. 1D, E).
Together with previous studies, (9) our results suggest that while the immediate activation of the Nrf2-ARE pathway maybe crucial for acute protection after the SCI, a late phase activation of some target genes of the Nrf2-ARE pathway may further contribute to recovery processes. However, we cannot exclude the possibility that the activation of Nrf2-ARE pathways in cells outside of the lesion area or even in periphery plays a role in Nrf2-mediated beneficial responses. In addition, as Nrf2 regulates the expression of numerous genes, it is quite possible that target genes other than those investigated in the current study may contribute to the beneficial effects. The latter hypothesis is supported by our observation that mRNA levels of Nrf2 remained elevated at least up to 42 days postinjury.
Nrf2-Deficient Mice Exhibit Impaired Outcome After Contusion SCI
Previous studies on SCI have shown that Nrf2 deficiency is accompanied by elevated NF-κB activity, and increased production of pro-inflammatory cytokines, eventually resulting in increased neuronal death and deterioration of hindlimb function in models of mouse SC compression (5, 6). To assess the effect of Nrf2 deficiency on the recovery of motor function after contusion SCI, we monitored locomotor performance of wild-type (WT) and Nrf2-deficient mice (Nrf2−/−) for 4 weeks after injury using the Basso mouse scale (BMS) (2). All the injured mice became almost completely paraplegic on the first day after the injury and then gradually displayed partial recovery of locomotor behavior (Fig. 2). Already at 7 dpi the locomotor performance of Nrf2−/− mice was significantly impaired when compared to WT mice (p<0.05). This reduction in the performance of the Nrf2-deficient mice continued at 14, 21, and 28 days after injury. At 4 weeks after injury, the Nrf2−/− mice attained an average BMS score of 2.8±0.2, which corresponds to extensive ankle movements or plantar placement of the paw, but not plantar stepping. The WT mice obtained an average BMS score of 4.7±0.6, which corresponds to occasional or frequent plantar stepping without coordination or frequent or consistent plantar stepping with coordination, but with the paw rotation. These results suggest that Nrf2 deficiency worsens the recovery of hindlimb motor function and propose that Nrf2 is essential for functional recovery after contusion SCI.

To assess the effect of Nrf2 deficiency on the morphology of the SC, we next examined the transverse SC area at the different levels around the injury epicenter on the sections stained with Luxol fast blue (LFB). The transverse SC area was reduced in the Nrf2−/− mice 28 days after SCI (Fig. 3A). Because it is known that the functional recovery after SCI correlates with the white matter sparing in the injury epicenter (2), we assessed the degree of white matter sparing in the contused SC 28 days after injury. LFB staining revealed that the myelinated areas were less preserved rostrally to the lesion epicenter in Nrf2−/− mice (Fig. 3B, C). This suggests that contusion SCI reduces not only the SC surface area but also white matter size in Nrf2-deficient mice. As white matter sparing influences locomotor recovery after SCI (2), the detected reduction in myelin preservation is most likely a reason why Nrf2−/− mice demonstrated impaired motor function recovery. This is consistent with previous findings demonstrating that Nrf2−/− mice exhibit greater demyelination than WT mice in experimental autoimmune encephalomyelitis. Moreover, administration of sulforaphane increased white sparing after rat SCI (9). Therefore, activation of Nrf2 appears to be required for proper remyelination after SCI.

Next, we assessed whether Nrf2−/− mice exhibited aggravated astrogliosis. We used glial fibrillary acidic protein (GFAP) histochemistry to assess the degree of astrogliosis at the injury epicenter, and 1 mm caudally and 1 mm rostrally to the injury epicenter. Astrogliosis was 2.2-folds higher in Nrf2−/− mice following SCI caudally to the injury epicenter (Fig. 3D, E). As astrocytes are one of the major contributors to cytokine and chemokine synthesis following SCI, activation of Nrf2 may limit astrogliosis and therefore inflammation.
Gene expression levels of Nrf2 target genes were assessed 3 and 7 days after SCI in WT and Nrf2−/− SC. Hmox1 and microsomal glutathione S-transferase 1 (mGST1) gene (Mgst1) expression were increased 3 and 7 dpi in both WT and Nrf2−/− mice (Fig. 4A, B). Interestingly, at 7 dpi Nrf2−/− mice exhibited significantly higher Hmox1 expression (p<0.001) when compared with WT mice (Fig. 4B). Mgst1 was highly upregulated in WT when compared with Nrf2−/− both 3 and 7 dpi, but this difference was not statistically significant (Fig. 4A, B). Nqo1 levels were lower (p<0.05) in Nrf2−/− mice compared with WT mice (Fig. 4A, B). Importantly, we observed that impaired motor recovery in Nrf2−/− mice is not mediated by reduced expression of such important Nrf2-target genes as Hmox1 or Mgst1, but instead is associated with diminished expression of Nqo1, prior to and at the time of the onset of the motor impairment.
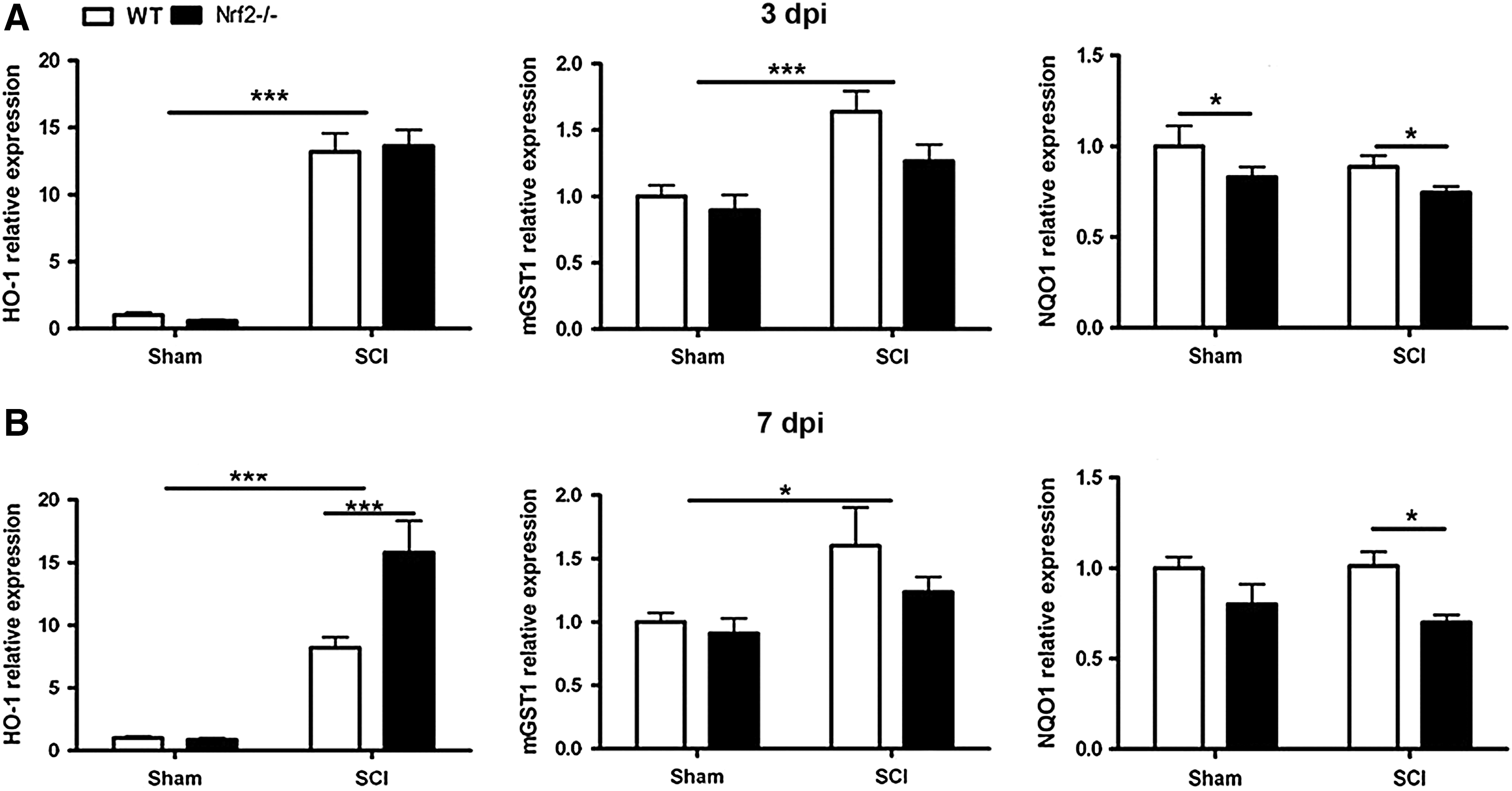
Apoptosis is the main mechanism of cell death following SCI and the role of OS in promoting apoptosis is well established. To our knowledge, the role of Nrf2 in regulating apoptosis in vivo has not been studied. We observed that expression of pro-apoptotic caspase 3 gene (Casp3) was induced at 3 dpi (p<0.05) (data not shown) and 7 dpi (p<0.001) in both studied genotypes (Fig. 5A). In contrast, the anti-apoptotic B-cell lymphoma 2 (BCL2) gene (Bcl2) was significantly downregulated at 7 dpi in both WT and Nrf2−/− mice (p<0.05) (data not shown). Importantly, the expression of the anti-apoptotic BCL2-extra large (BCL2-xl) gene (Bcl2l1) was significantly reduced at 7 dpi (p<0.05) only in Nrf2−/−mice (Fig. 5B). As induction of Bcl2l1, an anti-apoptotic gene, was prevented in Nrf2−/−, it is well possible that the aggravated damage after contusion SCI in Nrf2-deficient mice is also, at least in part, due to increased apoptotic death of SC cells, including oligodendrocytes.
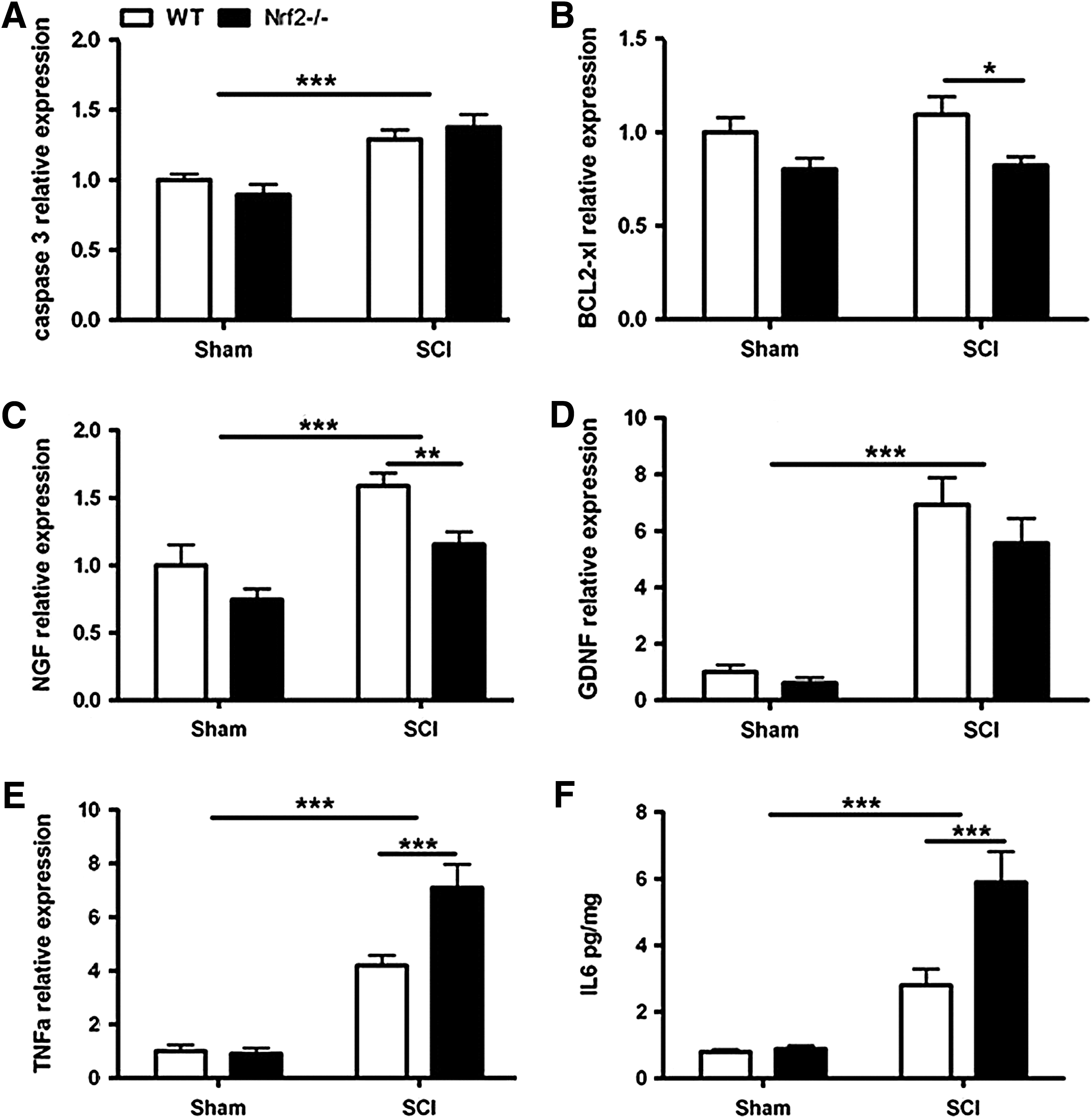
Growth factors modulate neuronal survival, neurite outgrowth, synaptic plasticity, and neurotransmission. Exogenous administration of growth factors has been proposed as a potential therapy for SCI, but whether Nrf2 regulates endogenous growth factor expression has not been described. We observed an induction of nerve growth factor (NGF) gene (Ngf) and glial cell-derived neurotrophic factor (GDNF) gene (Gdnf) at 3 dpi (Fig. 5C, D) and 7 dpi in both WT and Nrf2−/− (p<0.001) mice. The upregulation of Ngf at 3 dpi was significantly delayed in Nrf2−/− mice compared with WT (p<0.01) (Fig. 5C). In contrast to Ngf and Gdnf, the expression of brain-derived neurotropic factor (BDNF) gene (Bdnf) decreased at 3 dpi (p<0.001) and 7 dpi (p<0.05) in both genotypes (data not shown). So, the third group of genes with altered expression in Nrf2-deficient mice after contusion SCI was growth factors, as upregulation of Ngf was reduced in Nrf2−/− mice when compared with WT. At the same time we detected a clear trend toward delayed Gdnf upregulation in Nrf2-deficient mice.
Finally, we studied the expression and protein levels of number of pro- and anti-inflammatory cytokines. The expression levels of tumor necrosis factor-alpha (TNFα) gene (Tnf), interleukin-1 beta (IL1β) gene (Il1b), interleukin-6 (IL6) gene (Il6), and interleukin-10 (IL10) gene (Il10) were increased at 3 dpi in both WT and Nrf2−/− mice when compared with sham mice (p<0.001) (data not shown). At 7 dpi the level of Il1b and Il6 gene expression in animals with SCI reached the level of sham mice, while Tnf and Il10 remained significantly upregulated (p<0.001). At this time point the expression of Tnf was higher in Nrf2−/− mice (p<0.001) when compared with WT mice (Fig. 5E). The Cytometric Bead Assay revealed a significant elevation in IL6 (p<0.001) and decline in IL2, IL17α (p<0.001) protein levels in the SCs of both genotypes in comparison with sham mice. Of this, Nrf2−/− mice exhibited a significant increase in IL6 level (p<0.001) in comparison with WT mice (Fig. 5F). We noticed that from the investigated cytokines the expression of pro-inflammatory Tnf and Il6 was strongly upregulated in Nrf2-deficient mice at the time when the impaired recovery of these mice from SCI became evident. It is well known that upregulation of pro-inflammatory cytokines substantially contributes to secondary injury following SCI and it is believed that Nrf2 attenuates pro-inflammatory cytokines by inactivation of NF-κB. Indeed, the absence of Nrf2 resulted in greatly increased NF-κB activity in the acute phase of SCI. Moreover, sulforaphane treatment blocked the expression of pro-inflammatory cytokines via influencing the NF-κB signaling pathway (5, 9). As astrogliosis was apparently starting to increase in the SCI tissue during that time, eventually resulting in twofold expression of GFAP, it is possible that the increased inflammatory response in Nrf2-deficient mice is secondary to activation of astroglia, the major cell type contributing to inflammation after CNS injury and expressing Nrf2.
Collectively, these results suggest new aspects of Nrf2 action and indicate that Nrf2 is essential for the normal recovery from contusion SCI. Therefore, we hypothesized that enhancing Nrf2 expression by gene transfer of Nrf2 gene directly into the injured SC may be a promising strategy in SCI treatment.
Stable and Efficient Lentivirus-Mediated Gene Delivery to SC Neurons and Astrocytes
To investigate the efficiency of lentivirus-mediated gene transfer, a lentiviral vector carrying green fluorescent protein (LV-GFP) was injected into SCs of laminectomized mice. The pattern of GFP fluorescence was assessed 7 days later. GFP fluorescence was mainly localized to the grey matter (Fig. 6A).
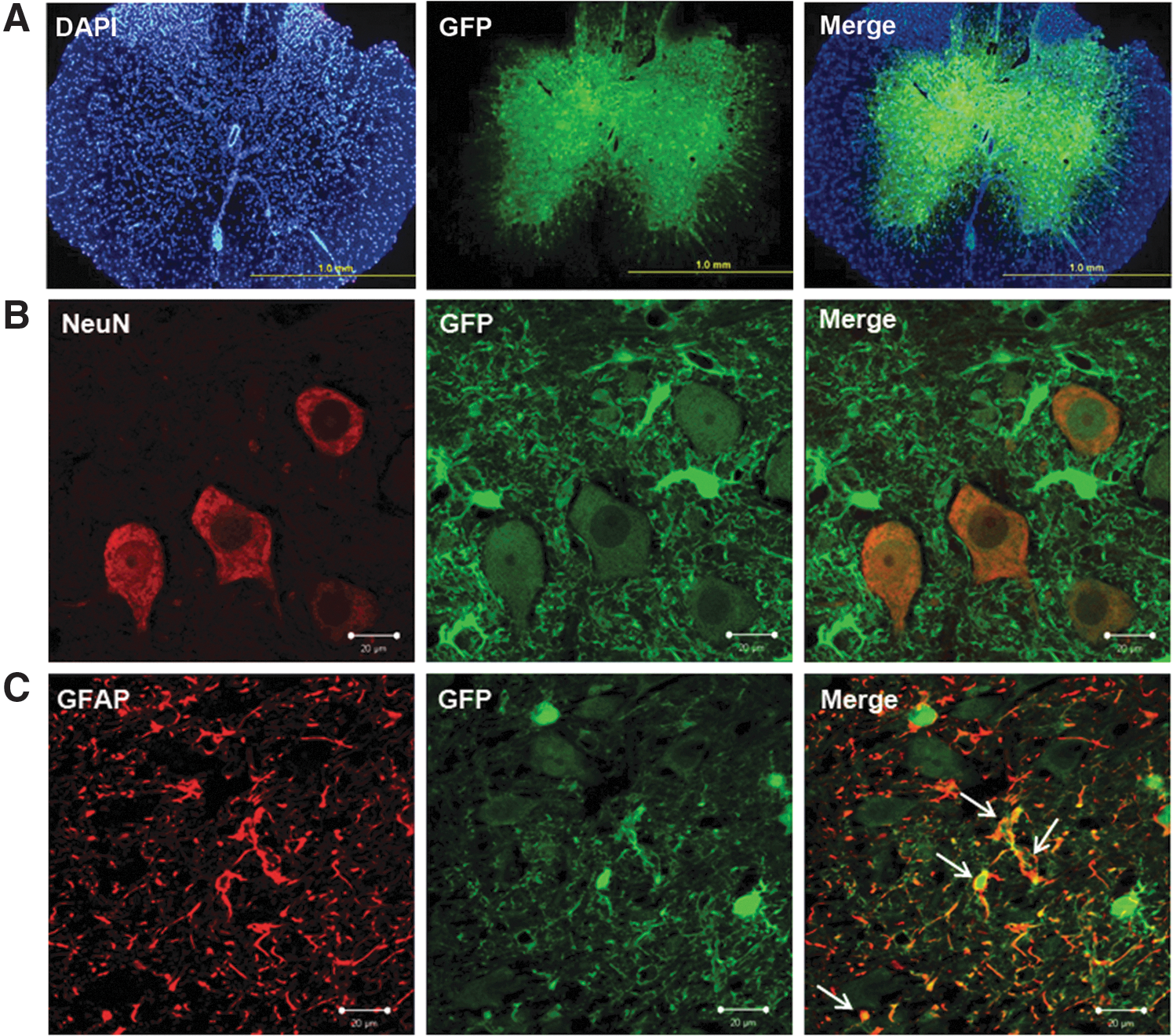
Staining of SC sections from GFP-transduced mice with neuronal marker NeuN and astrocyte marker GFAP revealed that both neurons and astrocytes were transduced by the viral vector (Fig. 6B, C). There were no GFP-positive microglia, as assessed by immunostaining for CD45 (data not shown).
To investigate the stability of lentivirus-mediated transfer, the expression of the transgene was assessed in SCs of mice 28 days after virus delivery. Intraspinal delivery of the lentiviral vector carrying human Nrf2 (LV-Nrf2) resulted in stable long-term expression; 28 days after LV-Nrf2 injection the expression of human Nrf2 was 350-fold higher when compared with background signal in LV-GFP group (data not shown).
Nrf2 Overexpression in Contused SC Protects from Lentivirus Toxicity and Improves Behavioral Recovery Compared with Lenti-GFP, but not Vehicle
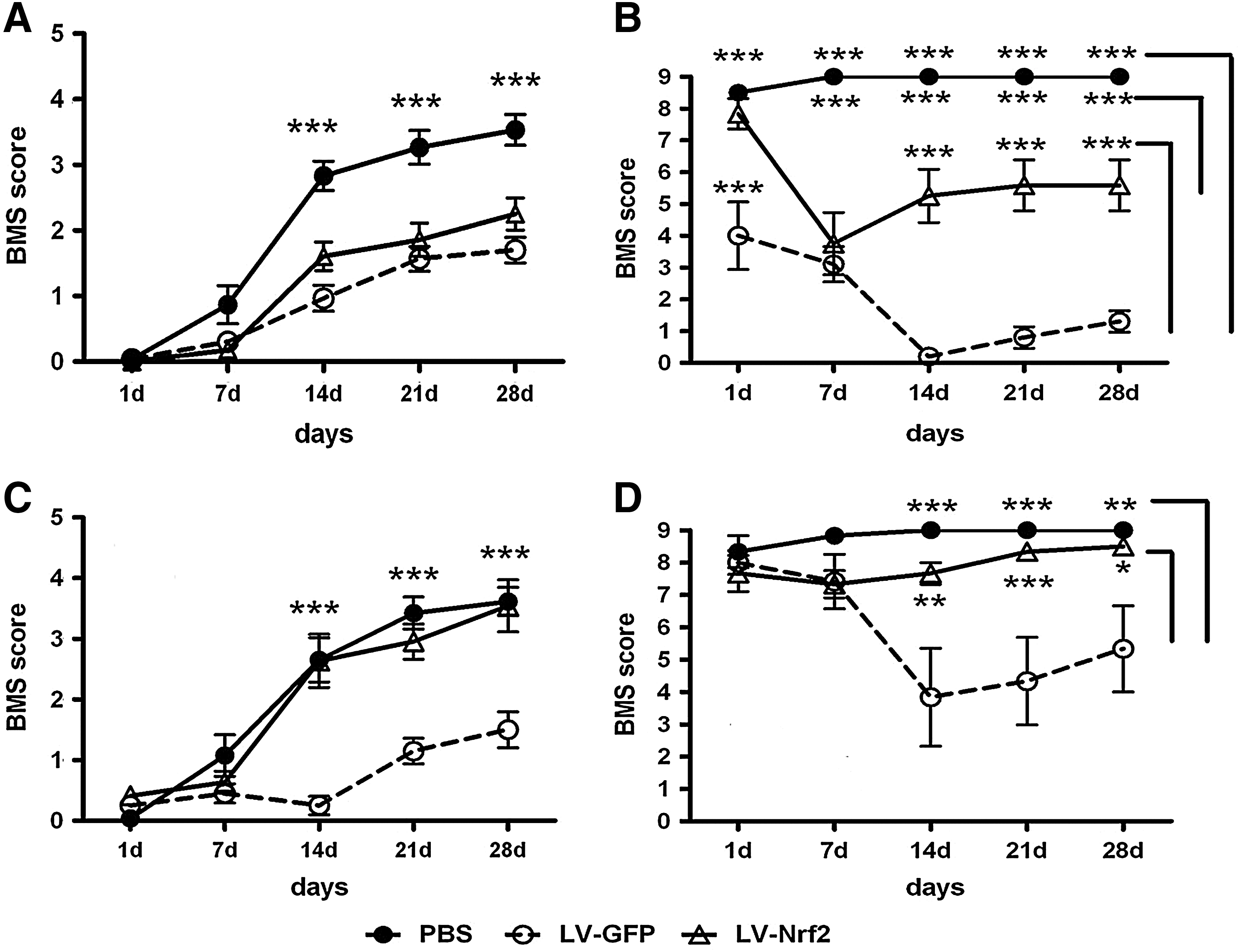
To assess the effect of lentivirus-mediated delivery of Nrf2 on behavioral recovery we next compared the BMS scores of injured mice injected with LV-Nrf2 with BMS scores of mice injected with either LV-GFP or the vehicle, phosphate-buffered saline (PBS) (Fig. 7A). Starting from 14 days after the injection of viral vector, the locomotor performance of mice injected with either LV-Nrf2 or LV-GFP was significantly worse when compared with mice injected with PBS (p<0.001). At 28 dpi mice in the control group treated with PBS in average exhibited plantar placement of the paw, dorsal stepping, or occasional plantar stepping. At the same time we did not observe a difference between BMS scores of mice injected with LV-GFP and mice injected with LV-Nrf2. In contrast to PBS-treated mice, mice treated with any lentiviral vector on average demonstrated only extensive ankle movements.
Despite the significant reduction in BMS score at 14 dpi between injured mice treated with PBS and mice treated with LV-GFP or LV-Nrf2 the size of the SC lesion was not affected, as measured by magnetic resonance imaging (MRI) (data not shown).
There were no significant differences between the BMS scores of noninjected sham mice or uninjured mice injected with PBS (data not shown). Uninjured mice injected with LV-Nrf2 exhibited significantly impaired locomotor performance (p<0.001) starting from 7 dpi when compared with noninjected shams and shams injected with PBS (Fig. 7B). The BMS scores of sham mice injected with LV-GFP were significantly lower than in all other studied groups.
Further, we increased the amount of viral vector used for injection by 33%. In contrast to the previous experiment, already at 14 dpi the locomotor performance of the injured mice treated either with PBS or LV-Nrf2 was significantly better in comparison with injured mice treated with LV-GFP (p<0.001) (Fig. 7C). However, we did not observe a significant difference between BMS scores of injured mice injected with PBS and injured mice injected with LV-Nrf2 during the whole observation period.
Similar to BMS scores obtained from injured mice, we did not detect differences between the BMS scores of uninjured mice injected with PBS or LV-Nrf2, whereas sham mice injected with LV-GFP exhibited significantly impaired locomotor performance starting from 14 dpi (Fig. 7D).
We suppose that both LV-GFP and LV-Nrf2 delivered to the SC in current titers are toxic and therefore worsen hindlimb recovery after contusion SCI. Increase in the amount of introduced viral vector encoding Nrf2, although apparently leading to the protection against lentiviral toxicity, may not be sufficient to improve hindlimb recovery compared to the vehicle.
Differential Gene Expression Profile Following Gene Transfer to the Injured SC
To understand the mechanisms that underlie the functional changes after the introduction of Nrf2 or GFP into the SC after SCI, we compared the expression of Nrf2 target genes and apoptosis-related genes, and pro- and anti-inflammatory cytokines and growth factors 3 days after transduction. Human Nfe2l2 mRNA expression was 1300-fold higher in LV-Nrf2-injected animals when compared with background signal in other studied groups that do not express human Nfe2l2 (p<0.001) (data not shown). Mouse Nfe2l2 and Hmox1 mRNA were upregulated in all injured groups, not depending on treatment (p<0.001) (Fig. 8A, B), whereas Nqo1 was higher in LV-Nrf2-treated group in comparison with other treated groups (p<0.05 for LV-GFP; p<0.01 for PBS) (Fig. 8C). In contrast, Mgst1 was significantly upregulated in PBS- (p<0.05) and LV-GFP (p<0.001)- treated groups in comparison with noninjured mice (Fig. 8D).
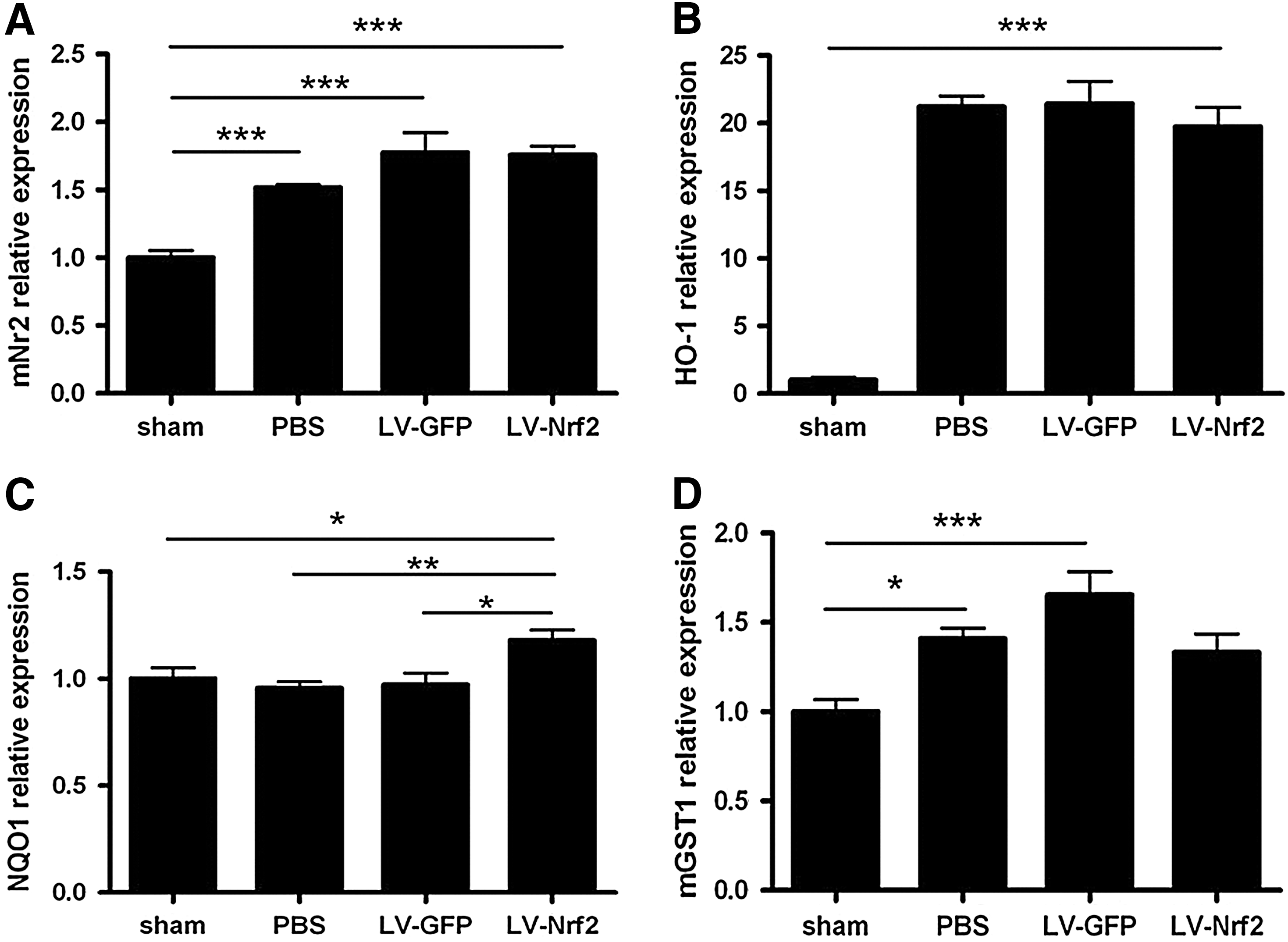
Interestingly, the expression of pro-inflammatory cytokines genes such as Tnf, Il1b, and Il6 was greatly increased in both LV-GFP- and LV-Nrf2-treated groups in comparison with uninjured mice and mice treated with PBS (Fig. 9A–C). We did not observe difference between cytokine expression in LV-Nrf2 and LV-GFP injected groups.

Casp3 expression was increased in all treated groups in comparison with uninjured mice (p<0.01 for PBS; p<0.001 for LV-Nrf2 and LV-GFP) (Fig. 9D). The expression of anti-apoptotic Bcl2l1 and Bcl2 was not altered (data not shown). In addition, the induction of Ngf and Gdnf was observed in the LV-GFP and LV-Nrf2 treated groups in comparison with uninjured mice and mice treated with PBS (Fig. 9E, F). There was no difference in Ngf expression in SCs transduced with LV-Nrf2 and LV-GFP, whereas Gdnf expression in the group treated with LV-Nrf2 was significantly higher (p<0.05).
Concluding Remarks and Future Directions
Exploiting the hypothesis of enhanced Nrf2 expression in injured SC we were able to achieve strong long-term overexpression of transgene throughout the grey matter, including both neurons and astrocytes. Moreover, we observed that Nrf2 overexpression induced the expression of Nqo1 and Gdnf. However, it did not result in significant improvement of functional recovery. In Nrf2−/−mice the expression of Nrf2 is absent since the first days of embryonic development in all SC cell types, whereas the overexpression of Nrf2 was introduced only right after the contusion, and it was limited only to neurons and astrocytes, excluding microglia. Since we did not study transduction of oligodendrocytes or other cells in the periphery, it is possible that the lack of Nrf2 overexpression on the outcome of SCI is due to late timing or cell type-specific restriction of expression. Assuming that in the previous studies sulforaphane targeted the Nrf2 pathway (5, 9) and as a small molecule reached this target in all cell types present in the SC, our hypotheses are in line with previous reports on the beneficial effects of this drug in the rat model of SCI. In future studies more thorough attention and effort should be placed on transduction of oligodendrocytes with the Nrf2 gene. Also, the role of those beneficial effects of sulforaphane that are unrelated to Nrf2 would be helpful to identify.
As an important side observation, we observed that the lentivirus used for overexpression studies in current titers is toxic to the SC, possibly by increasing inflammation (TNFα, IL6, and IL1β), but without extending the volume of injured tissue or changing the balance between pro-apoptotic and anti-apoptotic genes. It is of particular interest that lentivirus-mediated gene transfer, even at same titers that was previously successfully used in the hippocampus of a mouse model of Alzheimer's disease (4), resulted in severe paralysis of intact, healthy mice. Delivery of an increased volume (by 33%) of lenti-Nrf2 into the SC of naïve or contused mice significantly reduced the toxicity of viral vector, but still was not sufficient to improve hindlimb recovery compared to vehicle. Therefore, our results suggest that the SC may be exceptionally sensitive to lentivirus or any virus-mediated gene transfer, by so far unidentified mechanism. The results also underline the importance of the optimal titer of the virus vector used to gain functional improvement to avoid the toxicity associated with the gene transfer approach.
Notes
Animals
Animal experiments were conducted according to the national regulation of the usage and welfare of laboratory animals and approved by the Animal Experiment Committee in State Provincial Office of Southern Finland. For Experiment 1 (Nrf2-ARE time-course expression study) and Experiments 3 and 4 (Lenti-Nrf2 overexpression studies) C57BL/6J female mice between the ages of 2 and 4 months were used. For Experiment 2 (Nrf2-deficiency study) age- and weight-matched female Nrf2-knockout (Nrf2−/−) and their WT littermate controls on a C57BL/6J background of the same age were used. The genotypes of the mice were determined by polymerase chain reaction (PCR).
Spinal cord contusion injury
Nrf2−/− mice and their WT littermates were randomly assigned to experimental groups subjected to SCI as described below or only T10 laminectomy. Five percent and 1%–1.5% isoflurane in 30% O2/70% N2O were used for the induction and maintenance of anesthesia, respectively. After midline incision of skin and muscles laminas of T9-T11 were exposed. The T10 lamina was removed and moderate contusion injury was made using the NYU Impactor (W.M. Keck Center for Collaborative Neuroscience) by dropping the rod from a distance of 6.25 cm on exposed dura in Experiment 1 or Infinite Horizons Spinal Cord Impactor with the force of 60 kdyn (Precision Scientific Instrumentation) in Experiments 2–4. The muscles were sutured and skin was closed with wound clips. After surgery, the mice were allowed to recover overnight in cages placed on heating pads at 37°C. Thirty minutes before surgery, the mice received the first dose 0.1 mg/kg of buprenorfine s.c. (0.3 mg/ml, Temgesic®; Schering-Plough) followed by 0.1 mg/kg every 12 h for 3 days. Bladders were manually expressed until the normalization of their function.
Lentivirus-mediated gene transfer
Lentiviral vectors
Enhanced GFP- and human Nrf2-expressing lentiviruses under the control of human phosphoglycerate kinase promoter were cloned as described in Experiment 4. Lentiviruses were produced using standard methods. The viral titer was assessed by HIV p24 enzyme-linked immunosorbent assay (ELISA) (Perkin Elmer).
In vivo injections of viral vectors
T10 laminectomy and SCI were performed on C57BL/6J female mice as described above, and immediately after contusion injections of LV-GFP or LV-Nrf2 or PBS (Experiments 3, 4) were made 5 mm from midline at both the rostral and caudal ends of the laminectomy site. Two injections were stereotaxically performed for each mouse using a 5-μl Hamilton syringe and injecting minipump (Nanomite Injector Syringe Pump; Harvard Apparatus). A 33-gauge needle was lowered 1.3 mm beneath the dorsal surface of the SC, and 2 μl (Experiment 3) or 3 μl (Experiment 4) of PBS, LV-GFP, or LV-Nrf2 was injected at the rate of 0.5 μl/min. At 7 min after injection the needle was slowly raised, and the wound was sutured as described above. For all in vivo transduction studies virus vector titer was 1.88×106TU/μl.
Behavioral assessment
Hindlimb motor function was evaluated 24 h following SCI in Experiment 2, or SCI and viral vector injection in Experiments 3 and 4 and then weekly for 28 days in the open field using locomotor rating of the BMS (2) by two raters who were blinded to the experimental groups. Each mouse was observed for 4 min in each session and the hindlimb motor performance was evaluated with a nine-point scale. Mice with a BMS score greater than 1 at first day after injury were excluded from the further study.
Quantitative real-time reverse transcription-PCR
Mice were terminally anesthetized with tribromoethanol (Avertin; Sigma-Aldrich) and transcardially perfused with heparinized saline (2500 IU/L). SCs were dissected and ∼5-mm SC segments caudal and rostral to the injury epicenter were frozen at−80 C for future processing.
Total RNA was extracted from frozen SCs by TRIzol reagent (Life Technologies) according to the manufacturer's instructions. RNA concentration and purity was measured with Nanodrop 1000 spectrophotometer (Thermo Fisher Scientific). cDNA was synthesized from 500 ng of total RNA using random hexamer primers (Promega) as a template and Maxima reverse transcriptase (Thermo Fisher Scientific). The relative expression levels of mRNA encoding genes of interest were measured according to manufacturer's protocol by quantitative RT-PCR (StepOnePlus; Life Technologies) by using specific assays-on-demand (Life Technologies) target mixes. The expression levels were normalized to ribosomal RNA and presented as fold change in the expression±SEM.
Cytometric bead array
Frozen SCs were homogenized in buffer containing 10 mM Tris-HCl (pH 8.0), 5 mM EDTA, 150 mM NaCl, 10% glycerol, 0.1% Nonidet P-40, and protease inhibitor cocktail (Complete; Roche Applied Science). The level of IL2, IL4, IL6, IFNγ, TNF, IL17a, and IL10 protein in the SC homogenates was measured by using CBATh1/Th2/Th17 kit (BD Biosciences) according to manufacturer's instructions. The samples were run on FACSCalibur flow cytometer (BD Biosciences) and analyzed by using FCAP Array software (Soft Flow, Inc.).
Histology and immunohistochemistry
Animals were terminally anesthetized with tribromoethanol and transcardially perfused with heparinized saline (2500 IU/L), followed by 4% paraformaldehyde (PFA) solution. SCs were dissected and postfixed by immersion in 4% PFA for 21 h at 4°C. After cryoprotection in 10% sucrose for 24 h followed by 30% sucrose for 48 h, a 10-mm length of the SC containing the lesion site were embedded in Tissue-Tek O.C.T. Compound (SakuraFinetek), frozen on dry ice, and stored at −80 C. Serial crossections were cut at 20 μm thickness on a cryostat (Leica Microsystems GmbH) and stored at −80°C until processed for histological analysis.
Total SC area and white matter sparing
Total SC area, myelinated areas, and degree of myelin sparing were assessed by LFB staining on transversal sections with an interval of 200 μm. The selected slides were incubated in 0.1% LFB (Sigma-Aldrich) in acidified 95% ethanol overnight at 60°C. Differentiation was carried out with 0.05% lithium carbonate (Sigma-Aldrich). The epicenter of each injury was determined based on the section with the least percentage of spared white matter.
GFAP immunoreactivity
Selected sections were immunostained using rabbit polyclonal GFAP antibody (1:200; DAKO) followed by secondary antibody (goat anti-rabbit Alexa Fluor conjugated 568, 1:200 (Life Technologies). The sections were imaged with an Olympus AX70 microscope equipped with a digital camera (Color View 12 or F-view; SoftImaging Systems). The immunoreactive area was quantified by using ImagePro Plus software (Media Cybernetics). Astrogliosis was calculated as the area of GFAP immunoreactivity (area per total SC area) at the injury epicenter, and 1 mm caudal and rostral to the injury epicenter.
Distribution of LV infection with LV-GFP
Seven days after LV-GFP injection three mice were transcardially perfused with heparinized saline followed by 4% PFA. SCs were dissected, postfixed, and cryoprotected as described above. GFP expression was assessed on sections separated by 200 μm. Selected sections were immunostained using GFAP antibody, rat monoclonal CD45 antibody (1:100; Serotec), and mouse monoclonal NeuN antibody (1:200; EMD Millipore). The secondary antibodies were as follows: goat anti-rabbit Alexa Fluor conjugated 568, 1:200 (Life Technologies) for GFAP, goat anti-mouse Alexa Fluor conjugated 568, 1:200 (Life Technologies) for NeuN, and goat anti-rat Alexa Fluor conjugated 568, 1:200 (Life Technologies) for CD45. Finally, sections were mounted with Vectashield mounting media with Dapi (Vector Laboratories). The images were acquired with confocal Zeiss LSM 700 microscope (Carl Zeiss AG).
Magnetic resonance imaging
A 9.4 Tesla Varian scanner (Varian, Inc.) interfaced to a Varian DirectDrive console was used. A volume coil (diameter 35 mm) served as transmitter and receiver. Lesion volumes were determined from T2-prepared fast spin echo images (minimum repetition time 1.5 s, echo spacing 7.02 ms, four echoes, TE for preparation 30 ms, 16 averages, field-of-view 16×16 mm2, slice thickness 0.5 mm, data matrix 128×128, 8 contiguous sagittal slices). Data postprocessing was performed by an observer blinded to the treatment using in-house-built Matlab software (Aedes;
Statistical analysis
All data are given as mean±SEM. For comparing means of two treatment groups unpaired t-test was employed. Behavioral data were analyzed using repeated measures two-way ANOVA followed by Bonferroni's post hoc test. For multiple comparisons one-way ANOVA followed by Tukey's post hoc test or two-way ANOVA followed by Bonferroni's post hoc test were used. All statistical analysis were performed by GraphPad Prism version 5.03 for Windows (GraphPad Software;
Footnotes
Acknowledgments
The work was supported by the Academy of Finland. We thank Sara Wojciechowski and Riitta Kauppinen for technical assistance with behavioral assessment and Dr. Merja Jaronen for technical help with confocal imaging.