
Abstract
Introduction
I
Skin Structure and Function
The skin is a complex organ that is composed of three structurally and functionally disparate tissues. While the deepest layer of the skin, the fat-rich hypodermis, may be affected by chronic exposure to UVR, it is the dermis and outer epidermis that undergo the most pronounced acute and chronic remodeling in response to UV irradiation (108, 170). The epidermis, which varies in thickness between body sites from less than 100 μm on the forearm and buttock to greater than 600 μm on the heel, functions as a barrier to both water loss and pathogen ingress (20, 86, 124). The epidermis is sub-divided into several regions, beginning with the lowest single cell layer, the stratum basale, of metabolically active stem cells and their derived keratinocytes that lie above the dermis (83). These basal keratinocytes terminally differentiate over a period of 4 weeks to form the outermost stratum corneum, which is composed of metabolically inactive and keratinized squames that are embedded in a hydrophobic lipid matrix (86, 114). In addition to keratinocytes, the epidermis also contains smaller numbers of pigment-forming melanocytes, immunologically active Langerhans' cells and sensory neurons, and Merkel cells (81, 92).
The interface between the epidermis and the dermis, the dermal–epidermal junction (DEJ), is thought to interact with both basal keratinocytes in the epidermis and oxytalan fibers (elements of the terminal elastic fiber network) on the dermal side (60, 163). Central to the DEJ is a specialized basement membrane, rich in laminin-332 and collagen IV, that is characterized by α6β4 integrin-mediated keratinocyte attachments and by collagen VII anchoring fibrils on the epidermal and dermal sides, respectively (17, 26, 142). In addition to these molecular attachments, the topology of the DEJ, which in young, photoprotected Caucasian skin is characterized by intercalating dermal and epidermal papillae, may play an important role in dermal-epidermal attachment and shear-stress resistance (78, 152). However, most of the mechanical characteristics of skin are thought to be conferred by the supporting dermis.
The dermis, which may vary in thickness from 1 mm on the forearm to 2 mm on the thigh, contains hair follicles, eccrine and apocrine sweat glands, blood and lymphatic capillaries, sensory neurons, and a diverse cellular population of resident fibroblasts and immune cells (73, 76, 82, 84, 115, 126, 133). Despite this compositional diversity with regard to sub-structures and cell types, the tissue itself is composed predominantly of highly stable (half lives of years or decades) ECM proteins that continue to function largely in the absence of the damage detection and repair mechanisms which protect short-lived proteins (half lives of hours or days) within the intracellular environment (45, 63, 129, 156). Many of these proteins are thought to perform key mechanical roles: fibrillar collagens I, III, and V, for example, resist tensile forces; while negatively charged and, hence, hydrophilic proteoglycans such as versican and the glycosaminoglycan hyaluronic acid resist compressive forces. The dermal elastic fiber system, which is composed primarily of elastin and fibrillin microfibrils, confers passive recoil (18, 64, 71). In addition to these major structural proteins, many ECM components perform crucial biochemical roles by mediating: matrix–matrix interactions and, hence, assembly (for example, the small leucine-rich proteoglycans); cell-matrix interactions (as controlled primarily by adhesive glycoproteins and soluble cytokines); and matrix homeostasis (as a consequence of matrix metalloproteinase [MMP]-driven degradation and inhibition and transforming growth factor β [TGFβ] sequestration) (3, 44, 50, 53, 87, 122). In contrast to internal organs, these molecules and the cellular components of the dermis and epidermis are required to resist exposure to potentially damaging electromagnetic radiation.
Biological and Clinical Consequences of Exposure to Ultraviolet Radiation
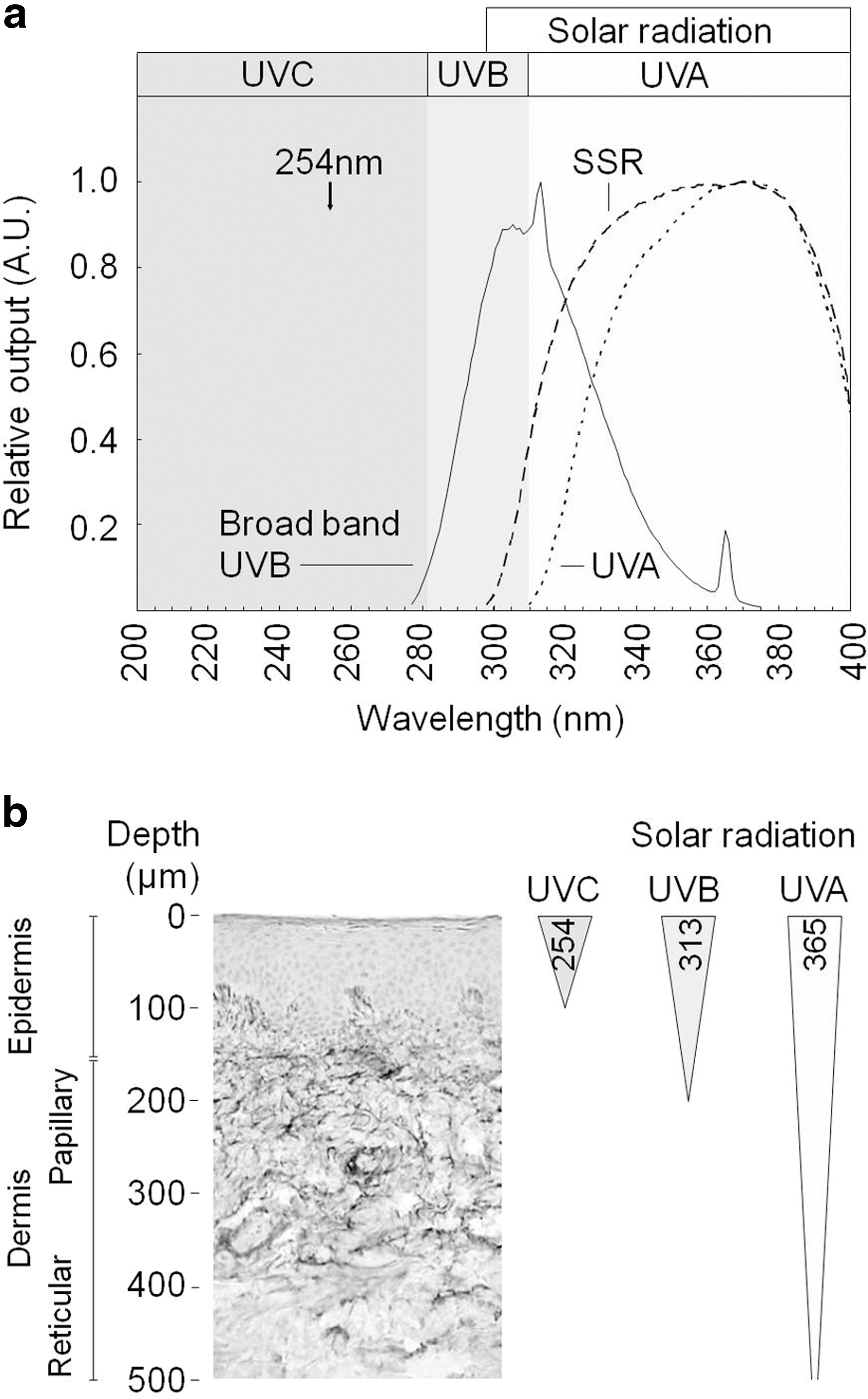
UVR, which forms only a small component of solar radiation, is conventionally split into high-energy UVC (wavelength 100–280 nm) and lower-energy UVB (280–315 nm) and UVA (315–400 nm) wavebands (Fig. 1a). However, while being biologically highly damaging, UVC radiation is absorbed by stratospheric ozone and is not, therefore, a component of terrestrial solar radiation (52). As a consequence, human skin at the Earth's surface is exposed to less energetic UVB and UVA radiation. UVB radiation, which comprises only 5% of terrestrial UVR, penetrates no further than the papillary dermis whilst UVA radiation, which comprises the remaining 95%, may penetrate the whole dermis to reach the sub-cutaneous fat (4, 31) (Fig. 1b). This exposure has both short- and long-term impacts on human health.
Positive effects of UVR
Humans, in common with other primates, obtain vitamin D either through dietary sources or through the UVB/heat-induced conversion of cutaneous 7-dehydrocholesterol to pre-vitamin D3 and then vitamin D3 in the skin (58). However, lack of sunlight exposure, particularly at higher latitudes, when compounded by a diet that is poor in vitamin D, may contribute to longer-term deficiencies which not only impact on the structure and function of the growing and mature skeleton but have also been linked to the development of disparate forms of cancer, multiple sclerosis, and cardiovascular disease (51, 58, 175). In addition to the beneficial effects of UVB-mediated vitamin D synthesis, UVA radiation may liberate bound nitric oxide, which, in turn, may exert a powerful cardioprotective effect (37, 113). Remarkably, when other known risk and protective factors are accounted for, mortality within the United Kingdom correlates linearly with latitude (79). These beneficial effects, both well established and putative, should be balanced against the profound negative impacts of excessive UVR exposure.
Negative effects of UVR
In the short term, UVR may induce acute clinical effects, including skin inflammation (in which vasodilation contributes to erythema or skin reddening) and immunosuppression [for in-depth reviews, see Refs. (22, 88)]. Chronically, UVR-induced epidermal deoxyribonucleic acid (DNA) damage and consequent mutation of tumor suppressors and oncogenes initiates nonmelanoma skin cancers and is associated with melanoma (15, 59, 104, 174). As a consequence of the adverse effects on health of both low (vitamin D deficiency and reduced NO-mediated cardio-protection) and high (principally skin cancer) UVR exposure, there is ongoing debate on the risk-benefit relationship of UVR exposure, in relation to the quantification of optimal UVR exposure for cutaneous vitamin D synthesis and public health advice on the use of topical UV filters (sunscreens) (48, 49, 89, 173). Regardless of the outcomes of this debate, there is a consensus that chronic exposure to UVR (photoexposure) induces profound changes in skin structure, which, in turn, manifest as apparent skin aging (photoaging) (105, 170). Establishing the main causative mechanisms of photoaging would impact not only commercially important cosmetic concerns but also attempts to understand the link between photoaging and skin cancer (170). In addition, chronically UVR-exposed skin provides an important human tissue in which the role that oxidative damage may play in driving systemic pan-species aging is studied (45).
Skin aging and photoaging
Although there is some commonality between the appearance of chronological (also known as intrinsic) and UVR-induced (extrinsic) skin aging, both the severity and speed of onset of outward manifestations such as wrinkles and tissue laxity and the nature of the underlying structural remodeling differ (14, 35, 129). Specifically, intrinsically aged skin, which is typically evident only at an older age (from the eighth decade onward), remains unblemished but is characterized macroscopically and functionally by the development of fine wrinkles and by reduced mechanical compliance and resilience (ability to recoil) (35, 36, 99). Microscopically, both the epidermis and dermis undergo atrophy, the DEJ becomes less convoluted, Langerhans' cells and fibroblast populations are depleted, and the dermal ECM loses structural oligosaccharides (proteoglycans and hyaluronic acid) and fibrillar collagens (35, 38, 46, 47, 68, 109, 121, 152). Where present, the effects of extrinsic aging are superimposed on this background of intrinsic aging.
When rigidly interpreted, the term extrinsic aging refers to tissue remodeling that may be induced by multiple external factors, including both UVR exposure and smoking; in practice, however, the terms photoaged and extrinsically aged skin are often used synonymously (102, 105). Clinically, severely photoaged skin appears deeply wrinkled and unevenly pigmented and is both less compliant and less resilient than photoprotected skin (1, 36, 77, 160, 170). This lack of resilience is particularly striking given that the photoaged dermis is characterized not only by the loss of fibrillar collagens and collagen VII-containing anchoring fibrils at the DEJ but also by the gain of oligosaccharides (proteoglycans and hyaluronic acid) and nominally resilient components of the elastic fiber system (12, 13, 26, 35, 97, 147). Crucially, however, the normal hierarchy of this latter system (elastin-rich elastic and elaunin fibers in the reticular dermis and fibrillin-rich oxytalan fibers in the papillary dermis) is lost in photoaged skin (105). The process begins with the specific loss of oxytalan fibers (and their associated components such as fibrillin and fibrulin-5) from the papillary dermis (65, 162, 163) and ends with the apparent deposition of elastin and fibrillin-containing material (termed solar elastosis) in the reticular dermis (12, 97, 105). In contrast to intrinsic aging, which is characterized by the gradual loss of ECM components from the dermis, any putative photoaging mechanisms should account for the differential spatiotemporal remodeling of specific ECM components.
Molecular Targets of UVR
An understanding of photochemistry is based on the Grotthuss—Draper law which states that “photon energy must be absorbed in order to have a subsequent reaction” (166). This absorption may directly affect the structure of the target molecule or generate reactive oxygen species (ROS) (via photosensitization), which, in turn, may damage the same target molecule or other molecular components (111). Alternatively, ROS may affect tissue homeostasis via their influence on cell signaling and phenotype (119). In order, therefore, to understand the mechanisms that drive photoaging of human skin, it is necessary to identify which molecules act as UVR absorbers (chromophores), where these molecules are located (in cellular or intracellular compartments), and what the downstream molecular consequences of this absorption are.
UVR chromophores in human skin
By definition, the term chromophore refers solely to the radiation-absorbing region of a molecule; in practice, however, UVR-absorbing biological polymers such as DNA are often referred to as UV chromophores (172). In the case of DNA, the absorption of UVR produces highly mutagenic photolesions, such as cyclopyrimidine dimers, in tumor suppressors or oncogenes and results in the initiation of skin cancer (30). Protection against such events is afforded by melanin, which is most abundant and most readily inducible in skin phototypes V and VI (127, 172). Other small molecules that act as UVR chromophores (and in some cases, as photosensitizers) include urocanic acid, the porphyrins and flavins, vitamin K and B6 derivatives, bilirubin, NAD(P)H, and advanced glycation end products. The reader is referred to excellent reviews by Young and by Wondrak et al. for detailed discussions of the identity of these skin chromophores and their complex absorption characteristics and resultant photochemistry (166, 172). However, while the most abundant molecules in human skin are the structural proteins of the dermis and it is these same components that undergo profound remodeling in UVR-exposed tissue, the role of ECM proteins as UVR chromophores and sensitizers remains less well defined.
Proteins as UVR chromophores
After absorbing photon energy, chromophores within proteins may enter a short-lived, singlet excited state, resulting in direct perturbations to molecular structure (111). Alternatively, this singlet state can undergo intersystem, crossing to a longer lived triplet state that has the potential to act as an intra-molecular photosensitizer. Such photosensitizers can, in turn, undergo either type I (electron transfer) or type II (energy transfer) reactions to form radical species (e.g., superoxide radical anion) or singlet O2 (1O2) (29, 42). Compelling experimental evidence for a link between photodamage and protein oxidation was first presented by Sander et al., who identified a dose-dependent accumulation of oxidation-induced protein carbonyls in the acutely UVR-exposed papillary dermis (125). However, the susceptibility of specific ECM dermal proteins to UVR-mediated degradation will be determined, in part, by their chromophore content that may be nonprogrammed (due to post-translational modification with other chromophores, including photosensitizers such as porphyrins, riboflavin, and glycation-derived cross-links) or programmed (as a consequence of amino-acid composition and macro-molecular structure) (111, 150, 166).
There is increasing experimental and theoretical support for the role played by post-translationally modified proteins in mediating UVR-induced DNA damage and, hence, cellular phenotype (165, 167 –169). In addition to these intracellular effects, post-translational modification of ECM proteins such as fibrillar collagens and elastin (via targeted lysyl oxidase [LOX]-driven cross-links or age and diabetes-induced glycation) may influence the mechanical behavior of tissues, including skin, lungs, and blood vessels and, hence, cellular phenotypes (via mechanotransduction pathways) (6, 62, 80, 143, 144, 164). The major structural proteins such as fibrillar collagens and elastin are not, however, the only targets of glycation. Less abundant, yet still biologically important ECM components, including adhesive glycoproteins, elastic fiber-associated proteins, and basement membrane components, may also react with glucose (5, 66, 148). The relative susceptibility of different ECM proteins to glycation (and, hence, to UVR-mediated damage via photosensitization) is likely to depend on: (i) the availability of free Lys and Arg residues after enzymatic cross-linking and (ii) cumulative absorbed UVR dose, as determined by protein architecture within the dermis, exposure to both glucose and UVR (dependent, in part, on molecular longevity), and the functional form of the protein. For example, while both collagen I and tropoelastin are long lived in human skin, and hence have ample opportunity to become glycated, their functional macromolecular structures in mature tissues (monomers assembled into large-diameter dense fibrils and fibers) ensure that in most cases the internal constituent proteins of collagen fibrils and elastic fibers will be protected from interstitial glucose and hence from glycation (71, 120, 140, 155).
Differential glycation of protein species may result from differences in their free Lys composition. Both the α1 chain of collagen I (Swiss Prot Accession: Q9UML6) and the precursor of elastin (tropoelastin: Q6P0L4), for example, contain similar numbers of Lys residues (38/and 44/1000 residues, respectively) but after LOX-driven cross-linking α1(I), collagen still contains 25–26 free Lys residues per 1000 compared with only 5–6 for tropoelastin (6). Finally, ECM proteins are unevenly distributed within the dermis and as a consequence, fibrillin-microfibril-associated proteins, which form arborizing (and, hence, low density) oxytalan fibers in the papillary dermis, could be key targets of glycation-mediated photosensitization (163).
Although post-translational modifications, macro-molecular structure, longevity, and architecture may play important roles in mediating protein susceptibility to UVR, a neglected area of study is the influence of relative amino-acid composition on the ability of individual protein species to act as chromophores.
Amino-acid composition as a determinant of protein chromophore load
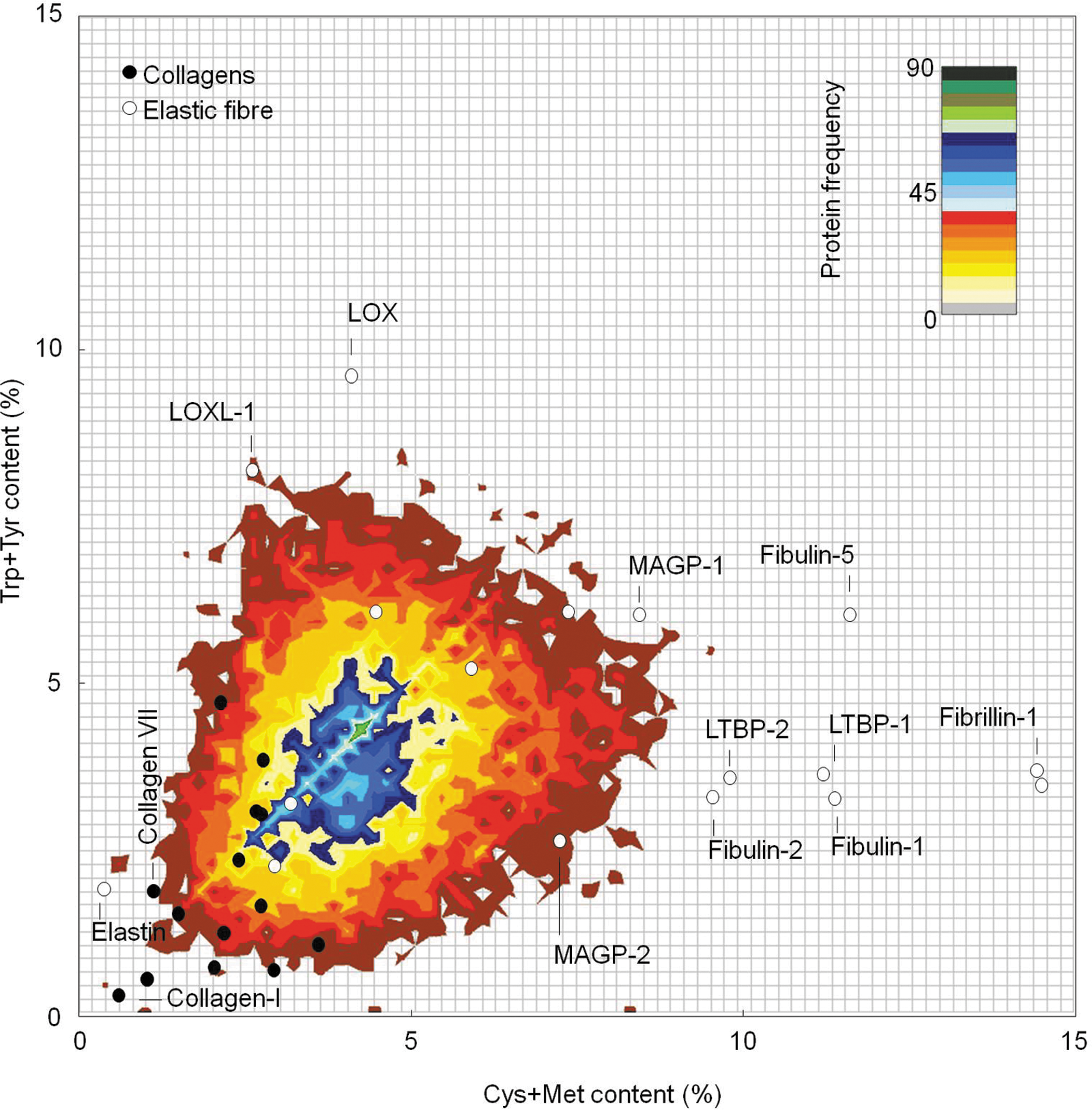
Individual proteins differ not only in their primary structures (amino-acid sequences) but also in their relative amino-acid compositions. Of the 20 amino-acid residues that comprise human proteins, Leu is the most common and Trp is the least common (∼10% and 1%, respectively, of the 19,889 characterized human proteins documented in the Swiss Prot database: Fig. 2). Furthermore, not all residues absorb the UVR wavelengths that are present in sunlight; UVB radiation is absorbed by Cys, Trp, Tyr, and His residues only (10, 34, 111). Of these residues, the susceptibility of Cys, Trp, and Tyr to photochemical modification is well established and from their absorption spectra, we can conclude that Tyr, and to a much greater extent Trp and Cystine (disulfide bonded Cys), are likely to be the key mediators of photochemical UVA-mediated damage to dermal proteins in vivo (27). Therefore, proteins that are rich in these residues and in Met may be key targets of photodegradation and indirect 1O2-mediated photo-oxidation [see Pattison et al. (111) for an in-depth review]. However, these amino-acid residues are under-represented in human proteins (Fig. 2), and we have previously proposed that relative amino-acid composition may be used to identify dermal ECM proteins or protein families which are either UVR/ROS resistant (primarily collagens) or labile (elastic fiber-associated proteins—with the exception of elastin itself ) (131, 149). By plotting the concentration of sulfur-containing (Cys and Met) against aromatic (Trp and Tyr) amino-acid residues, we can establish that this predicted differential UVR/ROS susceptibility is also evident when these groups (collagens and elastic fiber components) are compared with the entire human proteome (Fig. 3).

Experimental evidence for the degradation of collagens and elastin by UVR
The predicted resistance of fibrillar collagens to UVR can be tested experimentally. Fibrillar collagens (and in particular, collagen I) are the most abundant components of human skin and as a consequence, their susceptibility to in vitro UVR has been extensively studied. From an initial reading of the literature, it appears that fibrillar collagens are, in fact, UVR-labile. Reported UVR-induced structural changes include modifications to Tyr and Phe residues, altered electrophoretic mobility, and fragmentation of collagen monomers (23, 57, 61, 67, 94, 96, 98, 116). These structural changes may have functional consequences impacting collagen mechanical properties, resistance to protease digestion, thermal stability, triple helix formation, and fibrillogenesis (24, 25, 28, 43, 91, 93, 95, 98, 116, 135, 136). However, each of these studies employed UVR sources that emitted some short wavelength UVC radiation (Table 1). Such high energy sources can induce skin reddening (minimal erythemal dose [MED]) at doses of just 0.01 J/cm2 (32). In contrast, the MEDs for lower-energy UVB and UVA radiation that are present in sunlight are much higher: 0.05 and 56.5 J/cm2, respectively (39, 56). In order to compare the potential biological impact of UVR doses emitted from the disparate sources used by researchers over a time span of nearly five decades, we have, where possible, expressed these UVR doses as MEDs. (55, 74, 85, 131, 159). From this analysis, it is clear that nonphysiological UVR wavelengths, and in many cases doses (approximately four orders of magnitude greater than the MED), are capable of inducing profound structural and functional remodeling of fibrillar collagens in vitro. However, when physiologically attainable wavelengths and doses are used, the evidence for UVR-induced collagen denaturation is much less compelling (Table 2). UVA/UVB-rich radiation sources have a minimal effect on collagen structure and function even when the doses employed are more than two orders of magnitude greater than the MEDs of the respective wavebands (67, 90, 91, 131).
The structure and function (both mechanical and biochemical) of purified fibrillar collagen (predominantly collagen I) is profoundly disrupted by exposure to UVC-containing radiation sources (emitting primarily 254 nm) and delivering doses many orders of magnitude higher than the predicted MED for UVC-exposed human skin. UVR wavebands: UVC 100–280 nm (predominantly 254 nm); UVB 280–315 nm; UVA 315–400 nm.
MED, minimal erythemal dose; UVR, ultraviolet radiation; ND, not determined.
UVR, which is low dose and/or devoid of nonphysiological wavelengths, has no (or very limited) effect on fibrillar collagen structure, fibrillogenesis, or protease susceptibility. UVR doses are expressed relative to approximate MED equivalents as calculated for the various UV sources using published skin phototest data (55, 74, 85, 131, 159). All collagens are assumed to be primarily fibrillar (collagen I, III, or V) unless otherwise stated. UVR wavebands: UVC 100–280 nm (predominantly 254 nm); UVB 280–315 nm; UVA 315–400 nm; SSR 280–400 nm.
SSR, solar-simulated radiation.
In addition to the fibrillar collagens that confer tensile strength, the mechanical properties of the dermis are also determined by a complex network of elastin-rich elastic fibers. Although otherwise biochemically and structurally dissimilar to the collagen triple helix, the repeating domain structure of elastin is also largely devoid of UVA-absorbing and ROS-sensitive Cys, Trp, Tyr, and Met residues (69). When compared with the extensive literature on UVR/collagen interactions, the effects of UVR on the structure of elastin are poorly defined, although it has been demonstrated that elastin-derived peptides undergo structural modification after exposure to very high-dose UVC radiation (300 MED) (137, 138). Therefore, from the experimental evidence, it appears that the major long-lived structural proteins of the ECM are likely to be protected from the action of UVR and ROS by their amino-acid composition.
UVR-mediated degradation of ECM glycoproteins
In addition to an extensive network of collagen fibrils, elastic fibers, and hydrophilic proteoglycans, the ECM contains abundant structural glycoproteins that play dynamic roles in organizing and maintaining matrix structure (75). Foremost among these glycoproteins is fibronectin, a ubiquitous ECM component, which is present as both high molecular mass dimers in plasma and functional fibrils in the matrix (101). These fibrils contain binding sites for matrix components, including collagens, heparin, and fibronectin itself and crucially also for cell-membrane bound integrins (134). It is via these RGD (Arg-Gly-Asp) site/integrin interactions that fibronectin exerts influence over cell migration, morphology, and oncogenic transformation (75, 151). Compared with both collagen I and tropoelastin, fibronectin is relatively rich in the UVA chromophores Cys, Trp, and Tyr (0.3%, 2.2%, and 8.5% respectively). Hence, fibronectin may be a key target of UVR-mediated degradation in vivo. However, while there is evidence that human dermal fibroblasts respond to UVA exposure (both in vitro and in vivo) by synthesizing fibronectin, descriptions of the consequences of UVR exposure on fibronectin structure are, to our knowledge, limited to a study which we published in 2010 (11, 128, 131). We demonstrated using both polyacrylamide gel electrophoresis and atomic force microscopy (AFM) that fibronectin could be induced to aggregate by exposure to relatively low doses of broadband UVB radiation. In addition, we showed by using AFM that exposure to broadband UVB radiation altered the molecular dimensions of discrete fibronectin dimers. Such structural remodeling (with its attendant risk of cytotoxic amyloid formation) is a common response of diverse polypeptide chains to oxidation and/or UV irradiation both in vitro and in tissues such as the lens (100, 141, 158).
Since even small changes in fibronectin structure, induced by adsorption to hydrophilic or hydrophobic surfaces, for example, can affect cell adhesion, UVR-mediated fibronectin denaturation in vivo has, therefore, the potential to profoundly affect cell phenotype and hence tissue homeostasis (72, 130, 132). We should note, however, that these broadband UVB radiation-induced changes in fibronectin only became evident at multiple MEDs (4–10 times) but that this was not the case for key components of the elastic fiber system such as fibrillin microfibrils. These assemblies are both UV-chromophore rich (fibrillin-1 16.4% Cys, Trp, and Tyr content) and highly UVR susceptible.
Elastic fiber-associated proteins as key targets of UVR
The potential role of cell-derived ECM proteases in mediating dermal remodeling in photo-exposed skin is well established. UVR exposure not only up-regulates the expression of MMPs-1, -2, -3, -7, -9, and -12 (21, 39, 41, 123) but also promotes a pro-oxidative environment in human skin. In turn, this protease and ROS-rich environment may cause the activation of newly synthesized and sequestered MMPs by enzymatic or oxidative pathways, both of which operate on a common cysteine switch (8, 19, 117). Collectively, however, these enzymes are capable of degrading most dermal ECM components, including fibrillar collagens, elastic fiber constituents, proteoglycans, adhesive glycoproteins, and DEJ basement membrane components (3, 19, 110). Therefore, it is difficult to reconcile the concept of cell-derived ECM proteases as the sole mediators of matrix degradation with the complex spatial, compositional, and temporal ECM remodeling that characterizes chronically UV-exposed skin. Hence, while the suggestion that ECM components may be targets of UVR is not new, we have additionally proposed that elastic fiber-associated components in particular, by virtue of their high UVR-chromophore content, may be important mediators of dermal remodeling in photoaged tissue (28, 90, 131, 149, 166).
We have demonstrated experimentally that even sub-MED doses of broadband UVB radiation (20 mJ/cm2: 0.4 MED) are capable of inducing profound changes in fibrillin microfibril morphology (131). Similar changes in molecular structure, when arising as a consequence of inherited mutations in the fibrillin-1 gene, are associated with the life-threatening vascular pathologies that characterize the heritable connective tissue disorder Marfan Syndrome (70, 118). Hence, the susceptibility of these disulfide-bonded microfibrils (and potentially of the structurally related fibulins and latent transforming growth factor β binding proteins [LTBPs]) to environmentally relevant doses of UVR provides a potentially selective mechanism for (i) the early photochemical degradation of oxytalan fibers (fibrillin-rich microfibril bundles) in the papillary dermis and (ii) the subsequent remodeling of the elastic fiber system in the reticular dermis as a consequence of microfibril exposure to penetrating UVA radiation. In this latter case, photochemical degradation of fibrillin microfibrils is likely to be the triggering event that leads to aberrant TGFβ signaling, the up-regulation of both MMP and tropoelastin synthesis, the subsequent dysregulation of elastogenesis, and, hence, the deposition of elastotic material (solar elastosis) (16, 33, 106, 107, 171).
Mechanisms of Photoaging
The accumulating theoretical and experimental evidence for the role played by specific protein families in mediating downstream tissue remodeling leads us to propose a new model of photoaging that combines and extends the two existing models (direct hit and bystander) as defined by Wondrak et al. (166) (Fig. 4).
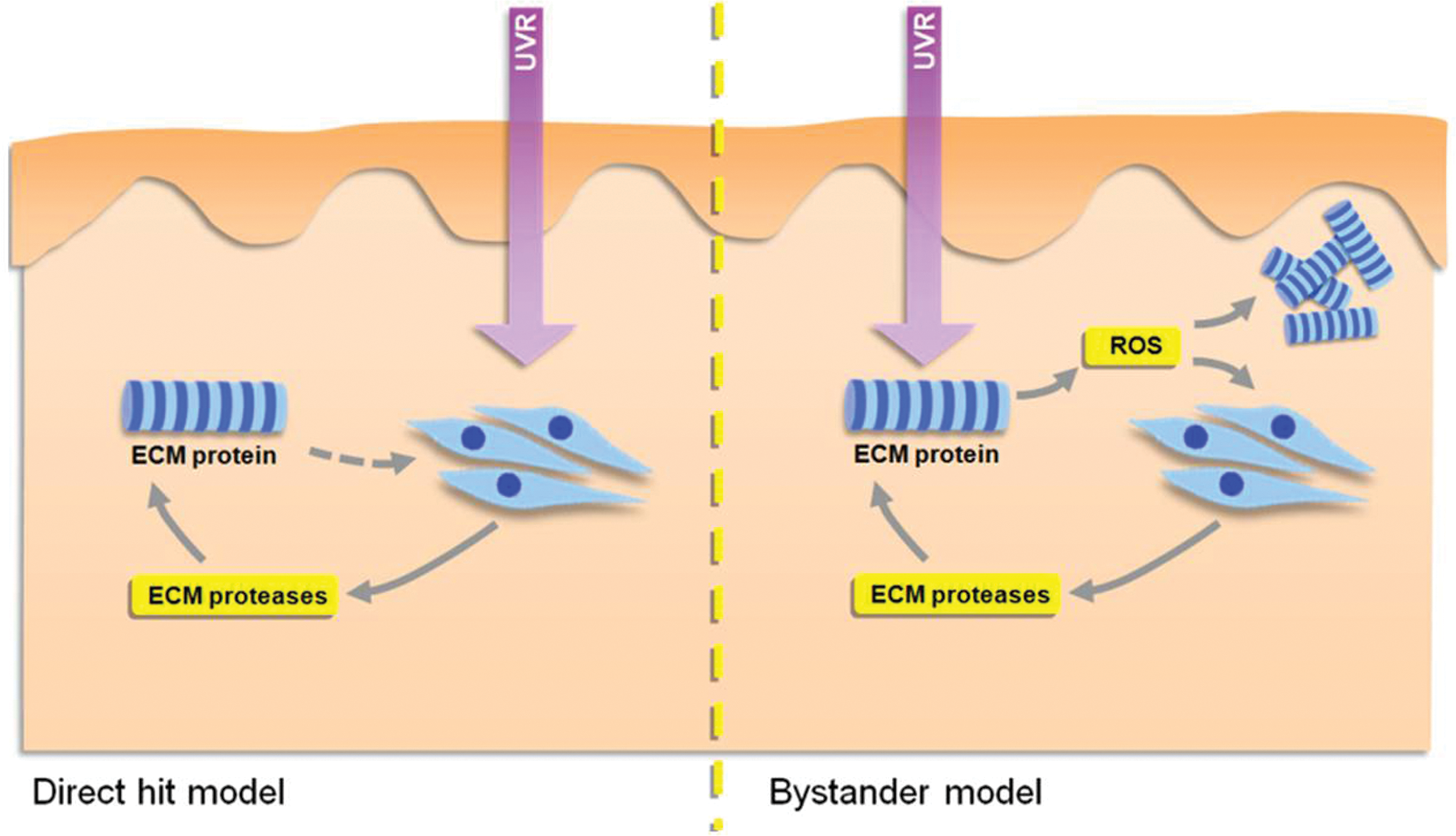
Direct hit and bystander models of photoaging
Although ECM remodeling is the key structural change in photoaged skin, existing models of photoaging have focused primarily on the influence of UVR on cell behavior. In the direct hit model, UVR is absorbed by intracellular photosensitizers to produce H2O2 or singlet oxygen, which act via cellular signaling pathways involving the inhibition of protein-tyrosine phosphatase-κ, activation of epidermal growth factor receptor, stimulation of mitogen-activated and c-Jun amino terminal kinases, and subsequent transcription of nuclear transcription complex AP-1 or the expression of interleukins-1α, -1β, and -6 to up-regulate the expression of ECM-degrading MMPs [for detailed reviews, see Refs. (119, 153, 170)]. By contrast, in the bystander model, the photosensitizer (which may be a glycated protein) is located in the ECM and liberates ROS, which both degrades ECM proteins directly and promotes MMP expression and activity (166).
Since these models are not mutually exclusive and in their present form cannot explain the specific degradation of key elastic fiber components (fibrillin-1 and fibulin-5) that characterize early photoaging, we propose a new combined model of photoaging: the selective multi-hit model, in which UVR-chromophore-rich cellular and extracellular photosensitizers mediate both direct and bystander effects (Fig. 5) (65, 131, 149, 163).
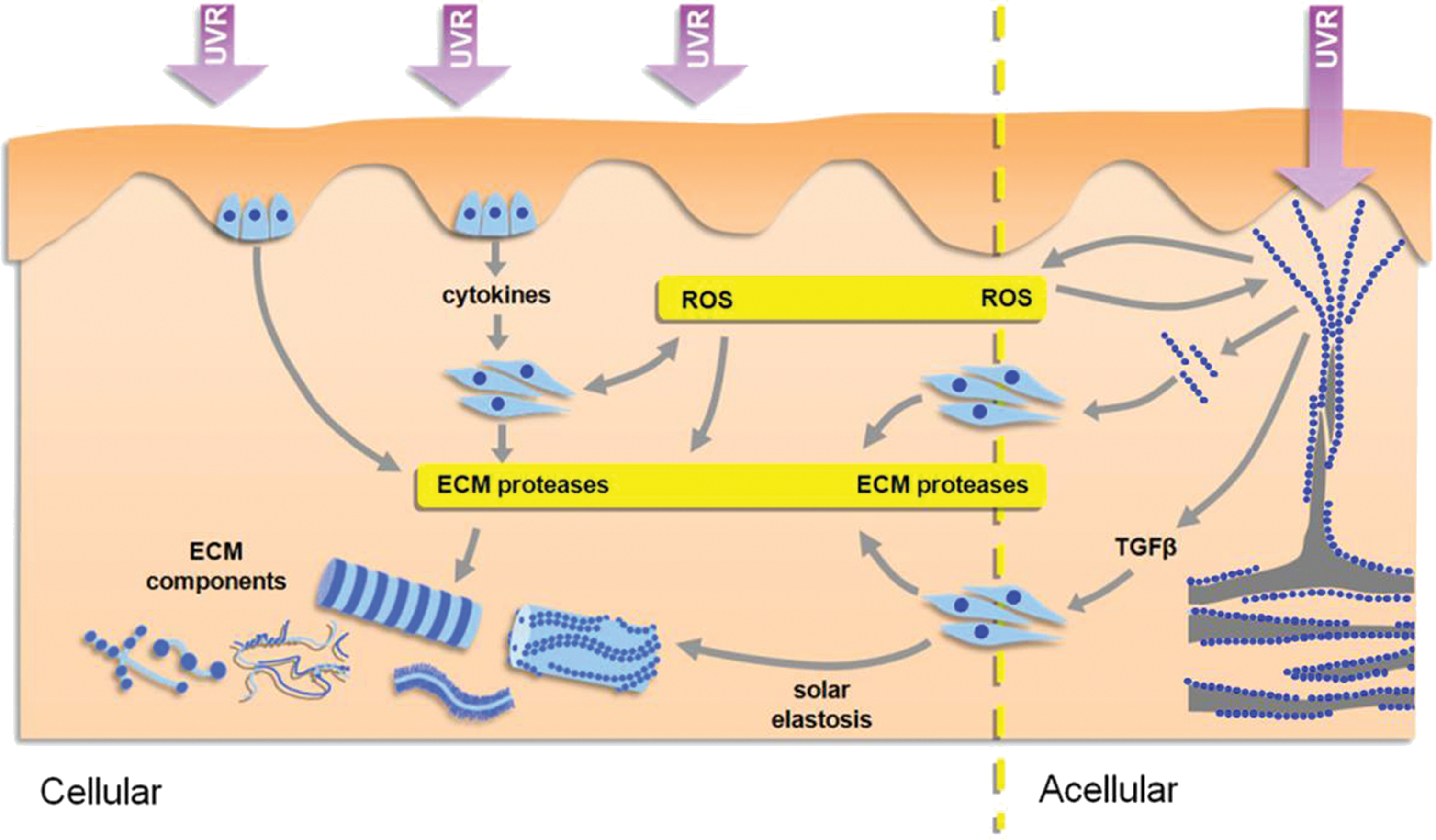
Selective multi-hit model
Identifying the key intracellular proteins that act as photosensitizers will require a combination of detailed bioinformatic analysis and experimental investigation. We can, however, begin to classify extracellular proteins as potential photosensitizers according to their amino-acid composition, susceptibility to glycation, location in the dermis, and macro-molecular structure. Hence, fibrillar collagens that are Cys-, Trp-, and Tyr-poor, resistant to glycation in their fibrillar form, diffusely distributed throughout the dermis, and present as dense fibrils and fibril bundles are unlikely to be targets for UVR (139, 157). Instead, UVR-upregulated MMPs (-1, -2, -3, and -9) may be the main mediators of fibrillar collagen (I, III, and V) degradation (19, 39, 41). Nonfibrillar collagens such as microfibrillar collagen VI are also UV-chromophore poor and their predicted lack of UVR susceptibility, combined with the absence of known proteases for the assembled form, may underlie the resistance of collagen VI to photoaging in vivo (103, 154, 161).
In contrast to the UVR-resistant collagens, the proteoglycan fibromodulin (which modulates collagen fibrillogenesis), the adhesive glycoproteins thrombospondin-1 and -2 and vitronectin, the basement membrane components (laminin-332, laminin-311) and perlecan (which will be exposed to relatively high UVB doses at the DEJ), and crucially, the microfibril-associated components of the elastic fiber system are potential UVR chromophores and photosensitizers (4, 9, 54, 131). These otherwise biochemically dissimilar elastic fiber components (fibrillin-1/2, fibulin-1/5, LTBP-1/2, MAGP-1 LOX, and LOXL-1/2/3) are Cys-, Trp-, and Tyr-rich, located primarily in the papillary dermis as arborizing microfibrils or on the periphery of elastic fibers, and, in the case of fibrillin-1, are susceptible to glycation (5, 71). We suggest, therefore, that UVR will interact with multiple UVR-chromophore-rich proteins in both the extracellular and intracellular environments to induce direct, oxidative, and enzymatic matrix remodeling.
Conclusions
Although the skin undergoes profound architectural and functional remodeling with chronic UVR exposure, the causative mechanisms remain poorly defined. Given the increasing evidence for the role of both intra- and extra-cellular photosensitizers in mediating the production of ROS and hence of oxidative damage and aberrant cell signaling, we propose a new selective hit model of photoaging in which proteins that are either readily glycated or which are rich in Cys, Trp, and Tyr act as cutaneous chromophores. Human skin, and in particular UVR-exposed skin, is an ideal model system in which to study aging of less accessible connective tissues, including blood vessels and lungs. The age-related pathological remodeling that is associated with the accumulation of damage by long-lived cellular components (DNA) and ECM proteins in these tissues is associated with the action of both glucose and ROS. Hence, identifying the key targets of age-related modifications (such as elastic fiber components) is an important step in understanding and hence ameliorating and repairing the effects of aging.
Footnotes
Acknowledgments
This work was supported, in part, by a program grant awarded to the authors by Alliance Boots, Nottingham, United Kingdom. In addition M.J.S., R.E.B.W. and C.E.M.G. are in receipt of Medical Research Council UK funding and C.E.M.G. is an NIHR Senior Investigator. We have also carried out independent commercial studies funded by BioMinerals NV; Croda Chemicals Europe Limited; Degussa AG; Kao Corporation; L'Oréal Recherche; Oriflame GTC Limited; Proctor & Gamble Technical Centers; and Unilever R&D Colworth. However, no commercial organization exerted editorial control over the contents of this article.