
Abstract
Introduction
W
Since this initial discovery of bacterial tyrosine phosphorylation, a variety of substrates have been identified. For a number, an effect on function has been demonstrated, such as in Bacillus subtilis, where phosphorylation of single-stranded binding proteins results in increased binding to DNA (61). Furthermore, additional work has linked tyrosine phosphorylation to a plethora of bacterial processes, such as cell division and growth (119), stress response (48), and even biofilm formation (45, 57, 107). However, to date, the best studied role of tyrosine phosphorylation is in regard to its effect on the regulation of secreted bacterial polysaccharides.
These secreted polysaccharides are split into two major groups. Extracellular polysaccharides (EPS) retain only a minimal association with the cell surface, and are often secreted into the extracellular milieu where many contribute to the formation of the matrix of bacterial biofilms. Conversely, capsular polysaccharides (CPS) are intimately associated with the cell (Fig. 1A, B). The attachment of CPS differs between bacteria also; in gram-positive bacteria, this is attached by a covalent linkage between the peptidoglycan of the cell wall and the CPS (91), while in gram-negative bacteria, a recent study has provided evidence that the CPS are anchored to the cell surface via the outer membrane protein Wzi (11) (Fig. 1C, D).

For a variety of bacteria, including Streptococcus pneumoniae, Staphylococcus aureus, and Escherichia coli, CPS and EPS are critical virulence factors, protecting the pathogen from phagocytic uptake, as well as playing roles in the maintenance of microbial biofilms. However, secreted polysaccharides can also have important commercial and industrial applications (e.g., xanthan Gum from Xanthomonas campestris). Indeed, vaccines against a number of bacteria, including Haemophilus influenzae, Neisseria meningitidis, and S. pneumoniae, contain purified CPS as the essential component. Work has shown that in a range of bacteria, biosynthesis of secreted polysaccharides is regulated by tyrosine phosphorylation, and associated bacterial protein tyrosine phosphatases (PTPs) and kinases (BY-kinase). Given the critical role of secreted polysaccharides in virulence, these proteins represent promising targets for the development of novel antivirulence drugs (22).
With the importance of bacterial tyrosine phosphorylation finally realized, studies of proteins regulating the phosphorylation levels has increased. Indeed, a large number of BY-kinases have been identified across the bacterial kingdom [as reviewed recently by (31)]. Whereas these lack the common eukaryotic kinase domains, they possess Walker A and B motifs, a characteristic of nucleotide binding proteins, and indeed structural studies have shown that they possess much similarity to these proteins (77). Interestingly, while gram-negative bacterial BY-kinases constitute a single protein, in gram-positive bacteria, they are split in two, such as in S. pneumoniae where both CpsC and CpsD are required for the functional tyrosine kinase activity (Fig. 1E, F) (69). As well as BY-kinases, bacterial PTPs have also been the focus of much research since their original discovery in the 1990s, with much work being undertaken to investigate their role in the regulation of the biosynthesis of secreted polysaccharides, primarily through an effect on the phosphorylation levels of the BY-kinase. This review focuses on the role of PTPs in the regulation of secreted polysaccharides, comparing the different families responsible, possible mechanisms, as well as the potential of these as a target for the development of novel antimicrobials.
Bacterial protein tyrosine phosphatases
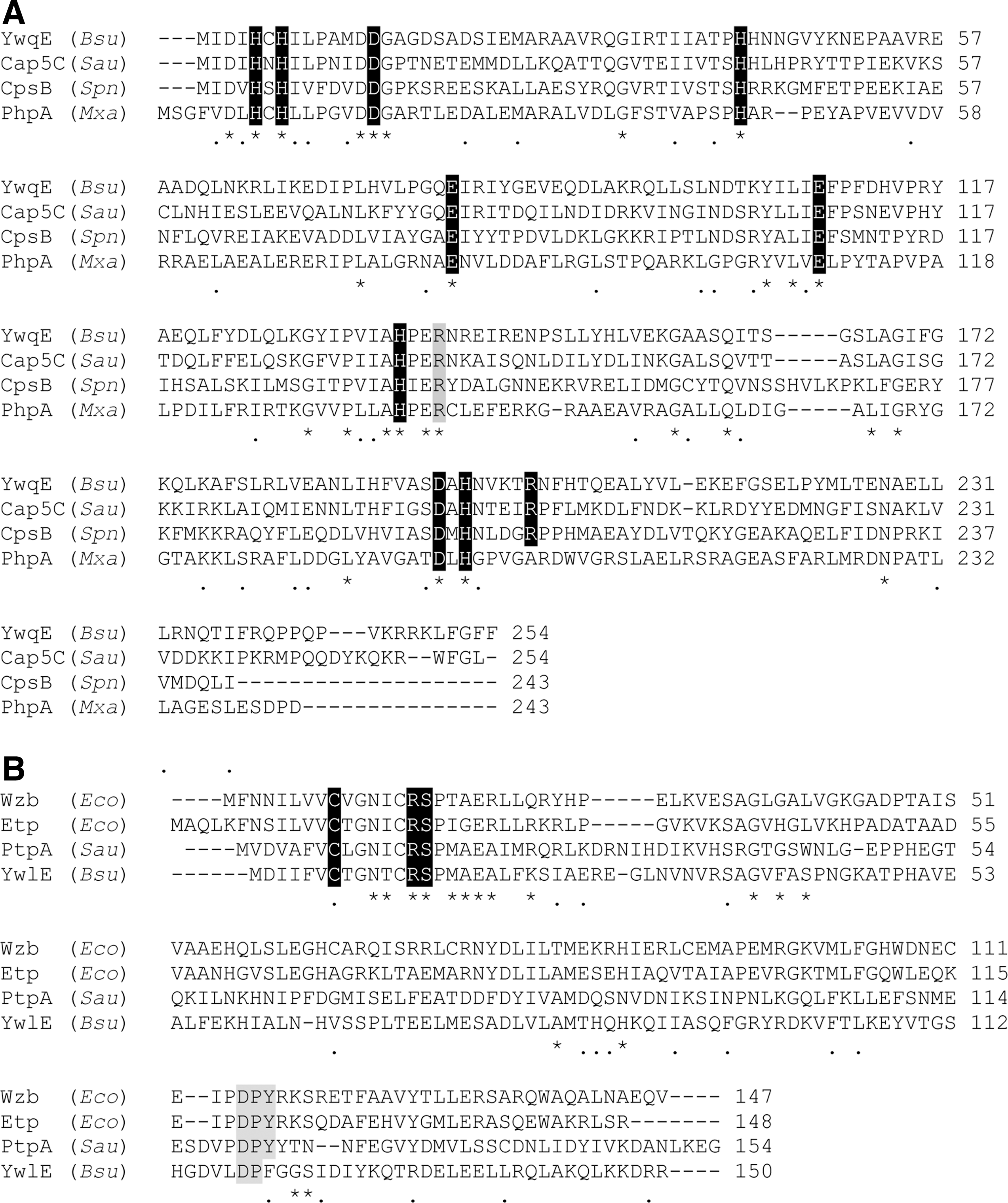
Since the discovery of the first bacterial PTP in 1992 (124), a large number of PTPs have been identified, which can be divided into three distinct groups (Table 1). These include eukaryotic-like and dual-specificity phosphatases, low-molecular-weight protein tyrosine phosphatases (LMW PTP), and the polymerase and histidinol family of phosphoesterases (PHP) (3). All are specific for phosphorylated tyrosine, apart from dual-specificity phosphatases, which additionally catalyze the dephosphorylation of substrates on phosphorylated serine and threonine residues. Whereas the first two groups are found across a spectrum of genera, both in bacteria and eukaryotes, the PHP family is found predominantly within bacteria, enhancing their potential as antimicrobial targets (3). In the past decade, numerous bacterial PTPs from the LMW PTP and PHP groups have been implicated in the regulation of the synthesis of the secreted polysaccharides EPS and CPS, suggesting that they play a major role in bacterial pathogenesis. Examples of the structures of key members of the LMW PTP and PHP families are shown in Figure 2, with key catalytic residues highlighted.

LMW PTP, low-molecular-weight protein tyrosine phosphatases; PHP, polymerase and histidinol family of phosphoesterases.
The catalytic mechanism of the LMW PTPs is well established, and involves the conserved signature motif C(X)5R that is the hallmark of the PTP superfamily in general (Fig. 3). Present in the phosphate-binding loop, the cysteine acts as a nucleophile and attacks the phosphorus atom of the nucleophile, while the arginine is critical for the binding of the substrate. In recent times, a novel level of regulation has come to the fore, with significant evidence that redox regulation is critical in the activity of PTPs with the C(X)5R motif. Such regulation has been reviewed in depth in recent times (103). The highly positively charged environment of the PTP-active site results in the catalytic cysteine being sensitive to oxidation. This results in a form of cysteine unable to act as a nucleophile, rendering the PTP inactive. Whereas little research has been undertaken into such an effect on bacterial PTPs, it is likely that LMW PTPs are sensitive to such regulatory effects.

The mechanism for the PHP family of PTPs, on the other hand, is less well defined as it lacks the characteristic C(X)5R motif. Indeed, unlike other PTPs they show optimal activity in basic pH and a crucial dependency on the presence of a metal ion, in particular, Mn2+ (60, 66). Recent crystal structures of representative members (CpsB and YwqE) (36, 46) (Fig. 2) have shed some light as to the mechanism of these metal-dependent phosphatases. This mechanism, as well as being dependent on metal ions, also requires an arginine residue in the active site and a nucleophilic attack by metal-bound water (36). The key conservation and highlighted differences between the PHP and LMW PTP groups are highlighted by their sequence alignments in Figure 4. Much research has been undertaken in recent times to understand the exact roles these PTPs play in bacterial physiology.
Importance of bacterial secreted polysaccharides
Over the years, the fundamental importance of both EPS and CPS has been clearly demonstrated (Fig. 5). Whereas CPS are critical for the full virulence of a wide variety of pathogens, their role is perhaps best understood and most studied in the major human pathogen, S. pneumoniae. In the pneumococcus, it is accepted as the single most important virulence factor (44), and is the serotype determining factor. Indeed, all pneumococcal clinical isolates causing invasive diseases are encapsulated, with unencapsulated bacteria only associated with superficial infections such as conjunctivitis. Furthermore, a multitude of studies have shown correlations of serotype with nasopharyngeal carriage prevalence, invasiveness, disease incidence, and even risk of death due to pneumococcal pneumonia (114). The essential requirement for CPS has also been illustrated in numerous pneumococcal animal models of infection (65, 68), suggesting that production of CPS is critical for all stages of infection, from asymptomatic colonization of the nasopharynx through to invasive disease of the lungs and blood. Similar results have been seen for other pathogens, such as S. aureus, with in vivo mouse models of infection showing that CPS are essential for the full virulence of the organism (54, 105).

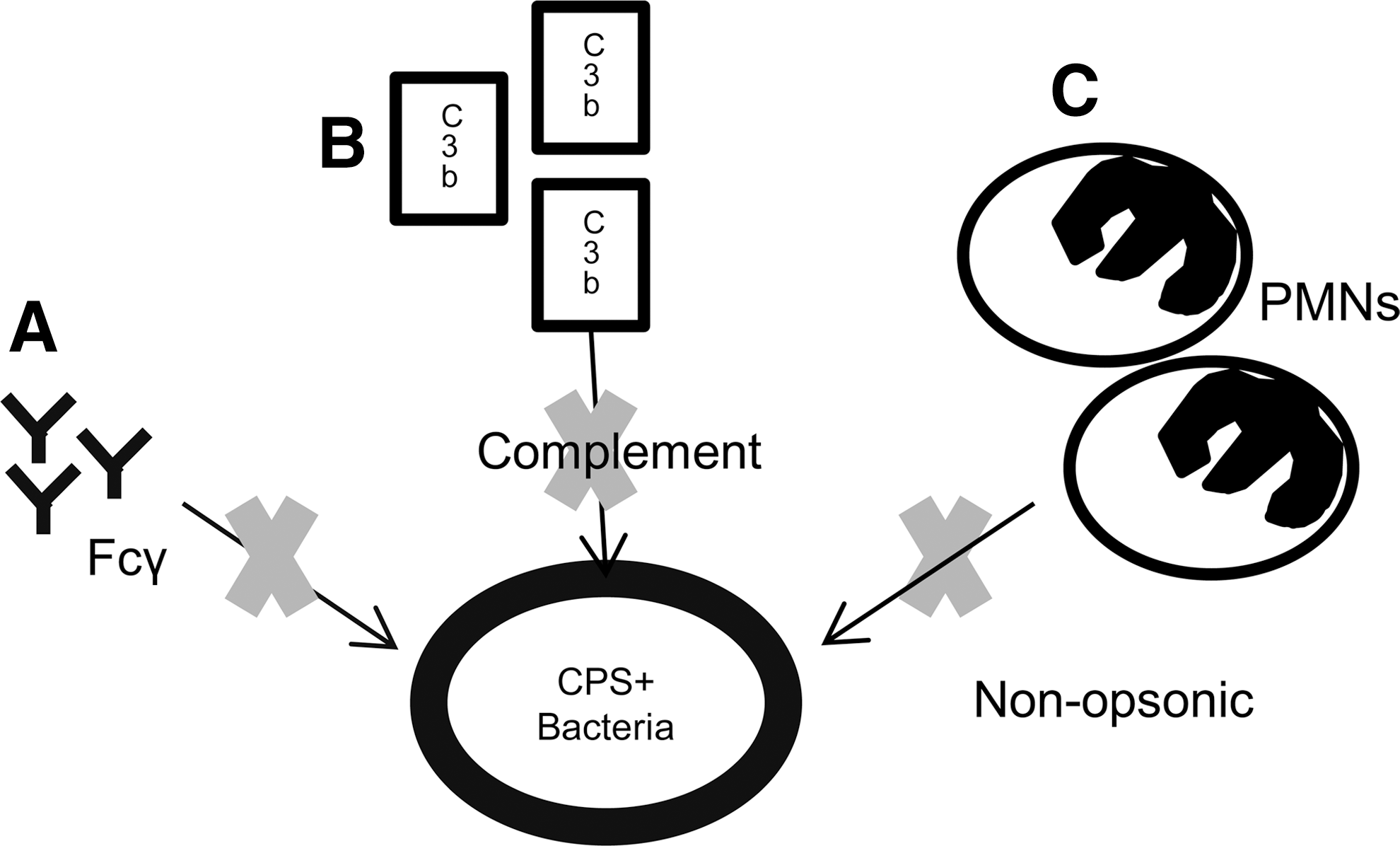
Thus, the obvious question is, why are CPS so important for pathogenicity? For many years, this has been postulated to be due to the ability of CPS to act as an antiphagocytic barrier in a variety of bacterial pathogens, including S. pneumoniae, S. aureus (113), and N. meningitidis (43) (Fig. 6). Hyams et al. (40) recently showed that unencapsulated pneumococci were more heavily opsonized by C3b/iC3b, thus resulting in increased phagocytosis by neutrophils, cells that are critical for the clearance of S. pneumoniae (97). Interestingly, the same work suggested that the CPS also prevented phagocytosis mediated directly by an antibody as well as by nonopsonic receptors. Thus, this data illustrated the multiple mechanisms that the CPS employ to protect the pneumococcus from phagocytosis.
Although critical for virulence, however, complete CPS expression, is not always advantageous for the pathogen. Numerous studies have found that CPS inhibit the ability of the pneumococcus to bind host epithelial cells (37, 102), due to the shielding of adherence factors (88). The pneumococcus also undergoes spontaneous phase variation between two phenotypes, opaque and transparent (115), with CPS, just one factor that varies. The opaque phase possesses increased CPS levels and is prevalent in invasive disease, while the transparent phenotype, with reduced CPS and thus a greater ability to adhere to epithelial cells, is favored during the asymptomatic colonization of the nasopharynx (117).
Secreted polysaccharides are also important for a wide variety of other bacteria. A recent study showed that secreted anionic CPS from S. pneumoniae, Klebsiella pneumoniae, and Pseudomonas aeruginosa, can sequester cationic antimicrobial peptides protecting the pathogens from this crucial host defense (53). Indeed, CPS in gram-negative pathogens are also crucial virulence determinants as has been shown for E. coli during urinary tract infections (29), K. pneumoniae, and even the emerging pathogen Acinetobacter baumanni (87). The CPS are important through similar mechanisms to Gram-positive bacteria via evasion of complement and inhibition of opsonophagocytosis (14) as well as contributions to biofilm formation (29) and by promoting resistance to complement-mediated lysis (13). In Vibrio cholerae, a major component of the biofilm matrix is Vibrio polysaccharide (VPS), which is essential for the formation of mature biofilm surfaces (122). In addition, synthesis of a slime polysaccharide in the gram-negative soil bacterium, Myxococcus xanthus, is essential for S-motility, fruiting body formation, and the formation of mature spores (47).
A wide range of bacteria also produce EPS, which have significant industrial applications, such as the hydrocarbon-degrading organism Acinetobacter lwoffii RAG-1, which produces a polymeric bioemulsifier emulsan (35). This is originally a minicapsule before being released in the medium as a protein–polysaccharide complex. Thus, production of EPS and CPS is critical for a wide variety of functions, including the virulence of a number of pathogens. Therefore, the mechanisms of regulation of polysaccharide biosynthesis are of fundamental importance and could be significant as targets for the development of novel antivirulence drugs, as well as to improve production yields and EPS size distribution.
Regulation of polysaccharide biosynthesis
The polysaccharides that make up both EPS and CPS vary greatly. This diversity occurs not only between bacteria, but also significantly within single bacterial species such as S. pneumoniae and E. coli. To date, greater than 90 serotypes in the pneumococcus have been identified (123), while in E. coli, more than 80 different capsular types or K-antigens have been characterized (78). These serotypes differ both in the many particular sugar moieties and also in the way these sugars are linked by varied glycosidic bonds. Thus, this results in greater variation of both branched and unbranched polysaccharides. There are three different pathways for polysaccharide biosynthesis: Wzy-dependent, the synthase-dependent, and the ABC transporter-dependent pathways (17, 123). Whereas the ABC transporter-dependent pathway is solely present in gram-negative bacteria, the Wzy- and synthase-dependent pathways occur in gram-positive bacteria also. The Wzy-dependent system is of particular interest to this review as this comprised a PTP and a BY kinase, which act in concert to regulate the biosynthesis of EPS and CPS.
Wzy-dependent capsule synthesis
In S. pneumoniae, the Wzy-dependent pathway is part of the CPS biosynthesis (cps) locus, which is located between the dexB and aliA genes on the S. pneumoniae chromosome (Fig. 7). In general, organization of the cps locus is conserved across all serotypes, apart from serotypes 3 and 37, which produce CPS via a synthase-dependent mechanism (123). At the 5′ end of locus, there is a group of four regulatory genes; cpsA, cpsB, cpsC, and cpsD (also known as wzg, wzh, wzd, wze). These highly conserved genes are then followed by cpsE (or wchA), the initiating glucose phosphate transferase, and then wzy and wzx, which encode the polysaccharide polymerase and flippase, respectively. The conserved order then ends and the distal part of the operon is considered serotype specific. The ensuing genes encode glycosyl and acetyl transferases, which are responsible for nucleotide glycosidic linkages and modification, and enzymes for synthesis of nucleotide-activated sugar precursors. Similar gene arrangements are found in other bacteria that encode Wzy-dependent CPS and EPS (Fig. 7). There are, however, some intriguing differences. For instance, S. aureus possesses two copies of the genes encoding the capsule regulatory genes; CapA, CapB, and CapC (92). These copies share significant amino acid similarity (CapA1 & CapA2 [48% identity]; CapB1 & CapB2 [53% identity]; and the PTP CapC1 & CapC2 [56% identity]), although to date, the importance of these multiple copies is not known.
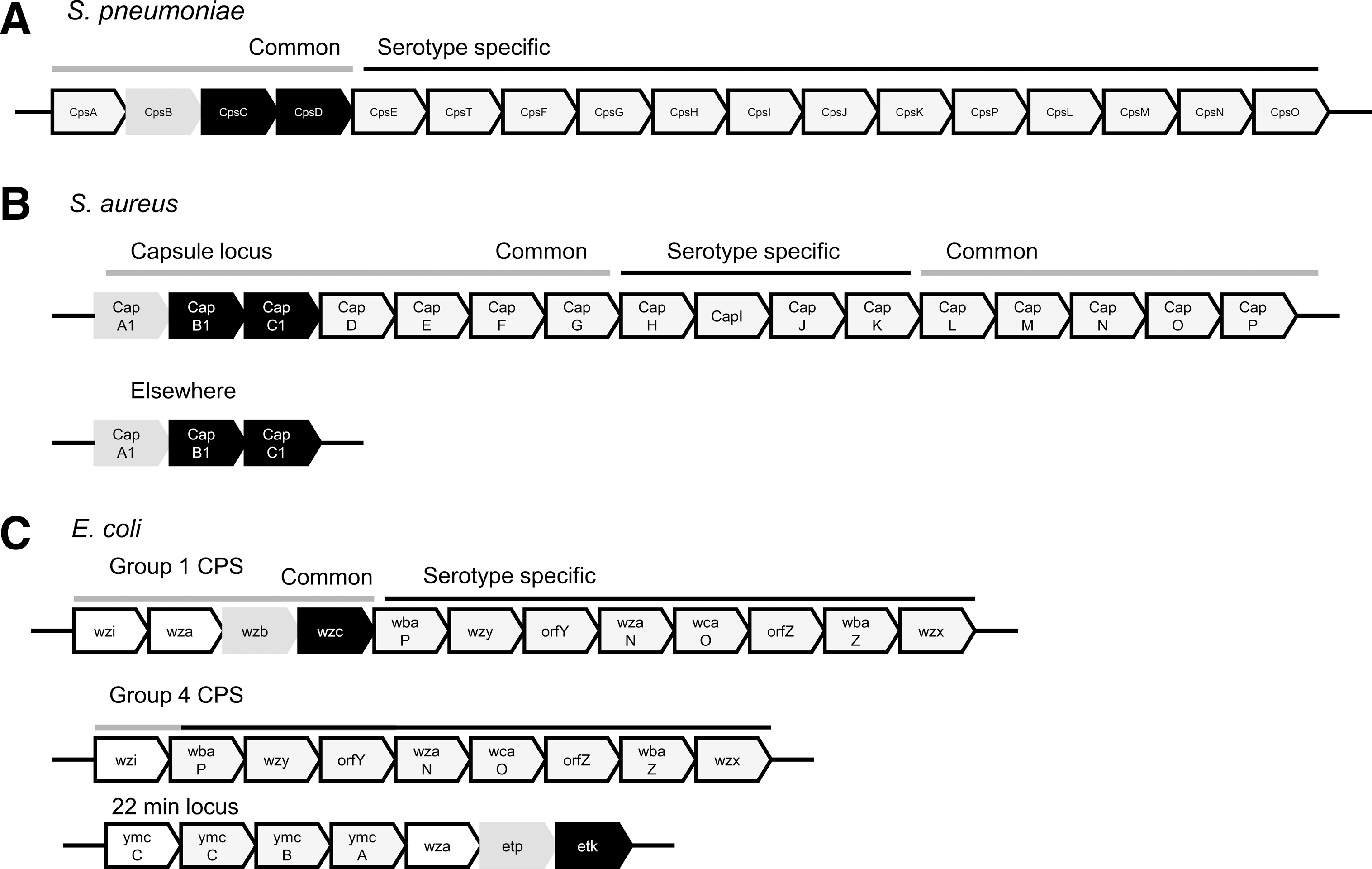
In E. coli, two of the four capsule groups are produced by a Wzy-dependent biosynthesis pathway. The genes for both are extremely similar (Fig. 7). The major difference is that Type 1 genes encoding capsule translocon Wza, PTP Wzb, and BY-kinase Wzc are present in the CPS locus, while for Type 4 CPS biosynthesis, the homologous genes encoding Wza, PTP Etp, and BY-kinase ETK are present on a distinct site of the chromosome known as the “22-min locus,” with all these genes essential for production of CPS (79) (Fig. 7).
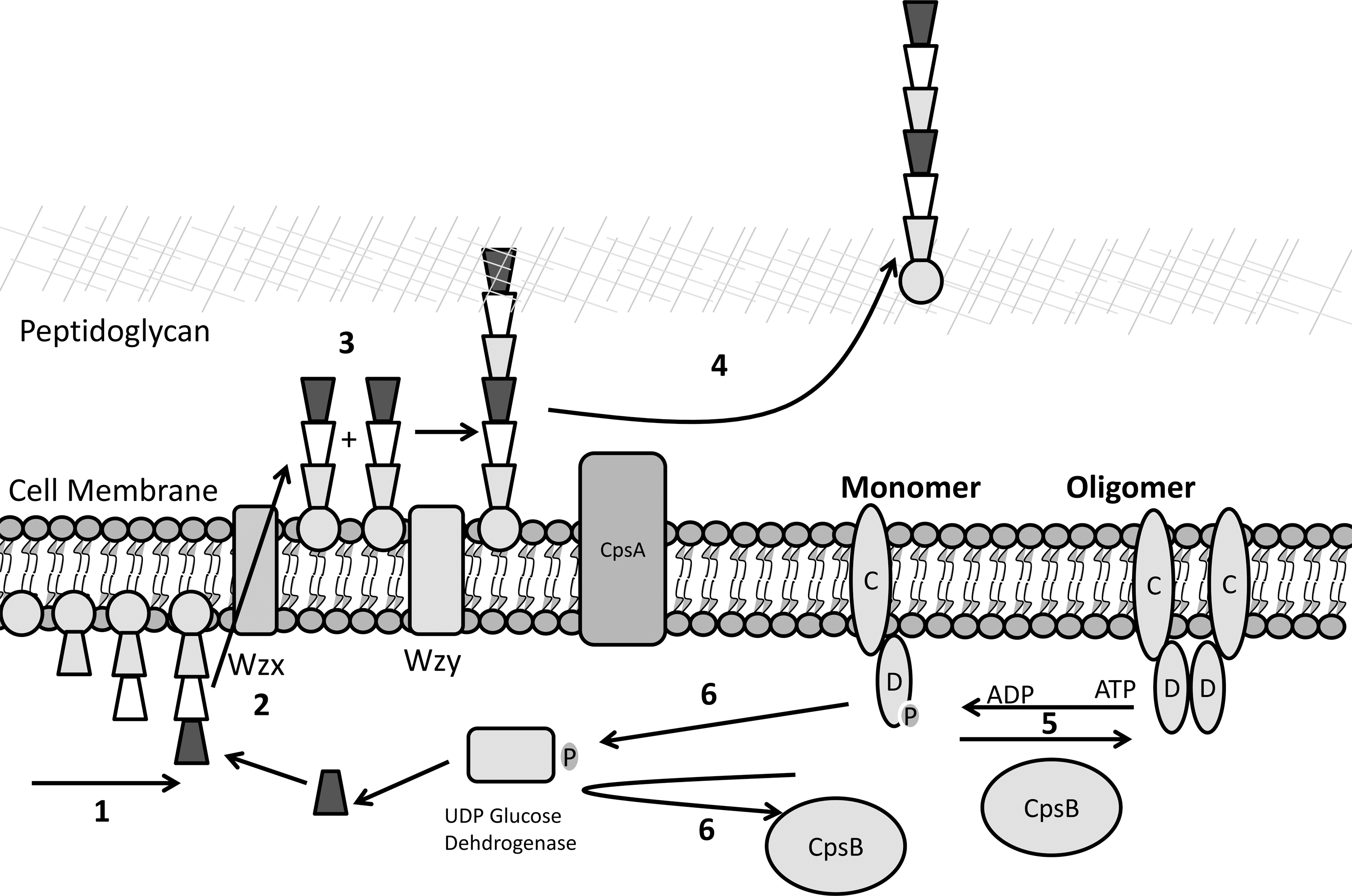
Whereas the model for Wzy-dependent synthesis of CPS is incomplete, much is known about this intricate process (summarized in Fig. 8). In S. pneumoniae, CPS synthesis starts in the cytoplasm with catalysis by a glycosyl transferase (CpsE), resulting in the transfer of a sugar phosphate to undecaprenol-phosphate (Und-P) via a nucleotide-activated sugar. This results in the formation of monosaccharide-PP-Und, the precursor molecule for oligosaccharide repeat unit synthesis. The particular sugar moiety, and thus initiating glycosyltransferase, varies depending on the particular serotype. Additional sugar moieties, either identical or different, are then added by glycosyl transferases until a complete oligosaccharide-PP-Und has been synthesized. This so-called repeat unit is then flipped to the external side of the cytoplasmic membrane by the Wzx flippase (52). The Wzy polymerase then extends these polysaccharides in a nonprocessive manner, adding single oligosaccharide-PP-Und repeat units to the reducing end of the chains. Once complete, the chain then needs to be attached to the cell. Most CPS of gram-positive bacteria are covalently attached to the cell, either to peptidoglycan or other components in the cell membrane (70, 123). In the pneumococcus, this ligation has recently been shown to be completed by CpsA (21) (Fig. 8). In gram-negative bacteria, this is obviously a more complicated process, due to the need to transport the polysaccharides from the inner membrane across the periplasm to the outer membrane where the CPS are attached by Wzi (11) (Fig. 1D). This transport is thought to be undertaken by members of the polysaccharide copolymerase (PCP) proteins (such as the E. coli BY-kinase Wzc) along with a member of the outer membrane auxiliary protein family (18, 70).
Regulation of capsule synthesis—contribution of BY-kinase and PTP
With the varied need for polysaccharide production in different environmental niches, regulation of its biosynthesis is critical. In the pneumococcus, a range of factors contribute to regulation, including oxygen levels (116), with suggestions that the activity of CpsB and CpsD varies in high and low oxygen levels (123). The catabolite control protein CcpA or RegM has also been implicated to play a role in the regulation of capsule synthesis (28). However, for those bacteria that synthesize their polysaccharide via a Wzy-dependent system, this regulation is mediated primarily via a phosphoregulatory system, consisting of the aforementioned BY-kinases and their cognate PTPs (Fig. 8). The genes encoding these proteins are highlighted in Figure 7, and for the most part are cotranscribed with the genes responsible for the biosynthesis of CPS and EPS. Whereas other genes vary between different capsular serotypes, these are highly conserved.
BY-kinases undergo intermolecular autophosphorylation, with oligomerization resulting in the insertion of the tyrosine-rich region of one monomer into the active site of another (6, 77). The PTPs are primarily thought to act on the BY-kinase dephosphorylating these tyrosines. Recent research has suggested that these BY-kinases and PTPs may also have additional targets (62), as discussed in detail below. To date, the best studied regulatory systems are found in E. coli (gram-negative), and S. pneumoniae and S. aureus (gram-positive).
The precise mechanism by which tyrosine phosphorylation of the BY-kinases affects polysaccharide synthesis is still unknown. In crystallographic studies, the nonphosphorylated form of Wzc (BY-kinase domain) forms an octomer that is disrupted following phosphorylation (6). Similarly, the structure of the BY-kinase from S. aureus (CapAB; a hybrid protein containing the cytoplasmic C-terminus of CapA along with CapB) shows that the kinase forms a ring-shaped octomer that disassociates upon phosphorylation (77). In addition, numerous studies using mutations in PTPs and BY-kinases have produced divergent results, with some suggesting that the phosphorylated form of the BY-kinase promotes synthesis (7, 120), while others see the opposite (67, 73, 76). These results have led to the hypothesis that both forms are important and that a cycling between the phosphorylated (monomer) and nonphosphorylated states (octomer) of the BY-kinase is critical for complete synthesis to occur.
As discussed previously, S. aureus possesses two copies of CapA, CapB, and CapC; the first two, a transmembrane adaptor and BY-kinase, while CapC is a PHP- type PTP. One copy of each is cotranscribed with the CPS loci, while another is present at a distant site on the chromosome (Fig. 7). Surprisingly, it has been shown for serotype five strains that CapB1 shows no tyrosine kinase activity in vitro, in the presence of either transmembrane activator proteins, CapA1 or CapA2 (92). However, CapB2 showed significant activity, particularly in the presence of CapA1 (92), which is transcribed at a distant site on the chromosome. There has been a recent suggestion that CapB1 may in fact be a pseudokinase; a protein that while containing a kinase-like domain is actually catalytically inactive (34), and may perform other auxillary roles as has been seen for the pseudokinase, MviN of Mycobacterium tuberculosis (27). The role of these genes within the bacterial cell is yet to be investigated. However, while one would expect the genes within the capsule locus to be of primary importance, not all regulatory genes in other Wzy systems are cotranscribed and present within the CPS locus, as evident in the production of Type 4 CPS in E. coli (Fig. 7).
Role of PTPs in the regulation of CPS and EPS synthesis
PTPs that play a role in the regulation of polysaccharide biosynthesis are primarily from either the LMW PTP or the PHP family of PTPs (Table 1). Significant research has been undertaken to investigate the role of these PTPs in the regulation of CPS and EPS synthesis (Table 2). For the majority, the phosphatase catalytic activity has been confirmed via purification of the particular PTP and investigation of activity against either a general substrate such as p-nitrophenyl phosphate or a specific phospho-tyrosine (PY) substrate (summarized in Table 2). CpsB is the representative PTP from S. pneumoniae and is one member of the PHP family of PTPs (3). Interestingly, results concerning its role in the regulation of CPS have varied (7, 66). Whereas our group has seen significantly less capsule in strains with otherwise isogenic in-frame deletion mutations in cpsB (68, 69), Bender et al. (2003) saw a slight increase (7). These differences may, however, be explained by the method of quantitation, as more recent research suggests that while CpsB mutants produce significantly lower levels of CPS compared with the wild type, they attach significantly more CPS to the cell wall (68). However, it is agreed that mutation of this PTP renders the pathogen unable to cause invasive disease to the same level as the wild type (7, 65), illustrating the overall importance of CpsB in the pathogenesis of this major human pathogen.
Phosphatase activity confirmed by activity against either pNPP or PY substrate.
Effect of mutation on biosynthesis of CPS or EPS. Deletion of PTP results in reduction (↓), increase (↑), no change (-), and not determined (N.D.).
Other roles for designated PTPs. –represents no other roles found to date.
PTPs, protein tyrosine phosphatases; CPS, capsular polysaccharides; EPS, extracellular polysaccharides.
The majority of S. aureus strains causing disease belong to CPS serotypes 5 and 8, and encode two PHP PTPs, CapC1 and CapC2. In Type 5 capsular strains, CapC1 does not appear important (90), while in Type 8 strains, CapC1 may play some role (112), although both these studies did not use defined mutations but were based on complementation studies of various CPS locus mutants. Furthermore, no work has been undertaken to investigate the role of the second copy of CapC that is present in the S. aureus genome in both type 5 and type 8 serotype strains (Fig. 7). Strains where CapC1 and CapC2 are deleted may be required to investigate the actual role of these PTPs in the production of CPS.
S. aureus also possesses two other PTPs belonging to the LMW PTP family of proteins (93, 109). Furthermore, a protein showing high homology to the family of LMW PTP is transcribed on the pneumococcal genome outside of the CPS locus (D39; Spd 1837). What role these likely PTPs may play in the regulation of CPS biosynthesis is yet to be determined in either pathogen. However, work has illustrated in vitro that CpsB and the LMW PTP from E. coli Wzb are interchangeable, with CpsB able to dephosphorylate the E. coli BY-kinase Wzc (36). Furthermore, we have also shown that an inhibitor of CpsB can also inhibit the in vitro phosphatase activity of Wzb (96), suggesting that there is some conservation of the active site. Thus, further research is required to investigate the possible role of these additional PTPs in these regulatory processes, and whether they can affect the phosphorylation levels of the BY-kinases.
In E. coli, two separate LMW PTPs, Etp and Wzb, are involved in the regulation of CPS synthesis (Table 2). Wzb plays an important role in Type 1 CPS biosynthesis with deletion resulting in significantly lower levels of CPS in E. coli K30 (120). Additionally, Wzb plays a role in the expression of the EPS colanic acid in E. coli K12, with deletion of wzb, but not etp resulting in loss of colanic acid synthesis (111). The PTP Etp, on the other hand, is responsible for Type 4 CPS biosynthesis (also known as O-antigen capsule synthesis) (72, 79), with deletion resulting in unencapsulated bacteria. Mutation of either PTP leads to increased phosphorylation of their cognate BY-kinase. While in vitro Etp can dephosphorylate Wzc and Wzb can dephosphorylate Etk, in vivo it appears that they act with specificity and only act on their cognate BY-kinase.
Additional LMW PTPs are present in a range of other bacteria, which appear likely to play a role in the regulation of polysaccharide biosynthesis. In Klebsiella pneumoniae, an increasingly important human pathogen for which CPS are essential for evasion of the immune response (13), a LMW PTP is present in the corresponding cps locus (80), although to date there is no data concerning its precise role (2). Interestingly, there seems to be some significant differences in the role of PTPs, as deletion of PTPs does not alter the production of CPS and EPS in all bacteria investigated.
In V. cholerae, VPS is essential for the formation of biofilms. The LMW PTP homolog VpsU has been shown to play an important role, with deletion of the PTP resulting in altered colony morphology, pellicle structure, biofilm formation, and VPS production (25). However, as yet, no investigation of the likely BY-kinase VspO (identified on basis of homology to E. coli Wzc) has been undertaken. Another LMW PTP, AmsI from plant pathogen Erwinia amylovora, is involved in the regulation of EPS, an essential prerequisite for E. amylovora pathogenicitiy. Both overexpression as well as deletion of AmsI result in decreased levels of EPS (9, 10). This lends more support to the proposal discussed above that the cyclic phosphorylation/dephosphorylation of the associated BY-kinase by a PTP is essential for the complete production of secreted polysaccharides. This is further supported with research into regulation of emulsan in A. lwoffii, an extracellular, polymeric, galactosamine-containing lipoheteropolysaccharide bioemulsifier, which has significant industrial applications. In this study, deletion of the LMW PTP Wzb results in a strain unable to produce polysaccharides. Deletion of either Wzb or cognate BY-kinase Wzc resulted in a similar reduction in biosynthesis of emulsan of ∼85% (73).
In Burkholderia cepacia, deletion of the LMW PTP BceD does not affect the production of EPS (cepacian) (24). However, this strain forms biofilms at a much lower level than the otherwise isogenic wild type. Indeed, a number of PTPs play a role in the formation of biofilms, with this associated with effects on EPS biosynthesis, which is essential for the formation of numerous biofilms. Interestingly, in P. aeruginosa, a dual-specificity phosphatase (TbpA) (107) contributes to the production of Pel EPS as well as that of extracellular DNA (81). However, the mechanism of this is significantly different from that of the other PTPs described here. TbpA effects the expression of the second messenger 3,5-cyclic diguanylic acid (c-di-GMP), essential for the production of the Pel EPS in this important human pathogen. The recent solution structure of TbpA has illustrated that the phosphatase shows significant structural differences from eukaryotic dual-specificity phosphatases, suggesting that it may be an attractive drug target (49).
An effect on biofilm formation by PTPs is not limited to TbpA and BceD. For the dental pathogen, Porphyromonas gingivalis, EPS production is essential for complete biofilm formation. P. gingivalis possesses a LMW PTP (Ltp1) with demonstrated phosphatase activity, as well as a protein showing homology to a BY-kinase, although they are present at distant sites on the chromosome. Studies of the role of Ltp1 show that the PTP contributes to the control of both EPS and biofilm formation, with mutants showing increased levels of both of these (57). Interestingly, this study showed that Ltp1 was also able to regulate transcriptional activity of the global regulator LuxS, suggesting that this PTP may be important for more global effects.
There has also been substantial investigation of additional PTPs of the PHP family as to their effect on the regulation of the biosynthesis of secreted polysaccharides. Deletion of PhpA from the M. xanthus results in increased levels of EPS (64), with the enzyme shown to act on BY-kinase BtkA (47), which is required for the formation of mature spores. In the Streptococcus thermophilus, mutation of the PHP PTP EpsB did not produce a demonstrable effect on EPS production, whereas mutation of EpsD, the cognate BY-kinase, did (63, 99). Interestingly, EpsB and EpsD modulated the enzymatic activity of phosphogalactosyltransferase (EspE), the initial glycosyl transferase responsible for the first step of EPS biosynthesis. Deletion of epsB increased the EspE activity, suggesting it was able to play a role in the suppression of EPS, perhaps in some conditions (63). Similarly, in Lactococcus lactis, deletion of the PHP member PTP epsC had only a minimal effect on the production of EPS, while deletion of the BY-kinase (epsB) resulted in a loss of EPS (75). However, the same study suggested that the nonphosphorylated form of the BY-kinase drove biosynthesis of EPS, which indicates that the PTP should play a significant role (75).
PTPs phosphorylate UDP-glucose dehydrogenases; possible mechanism for an effect on biosynthesis?
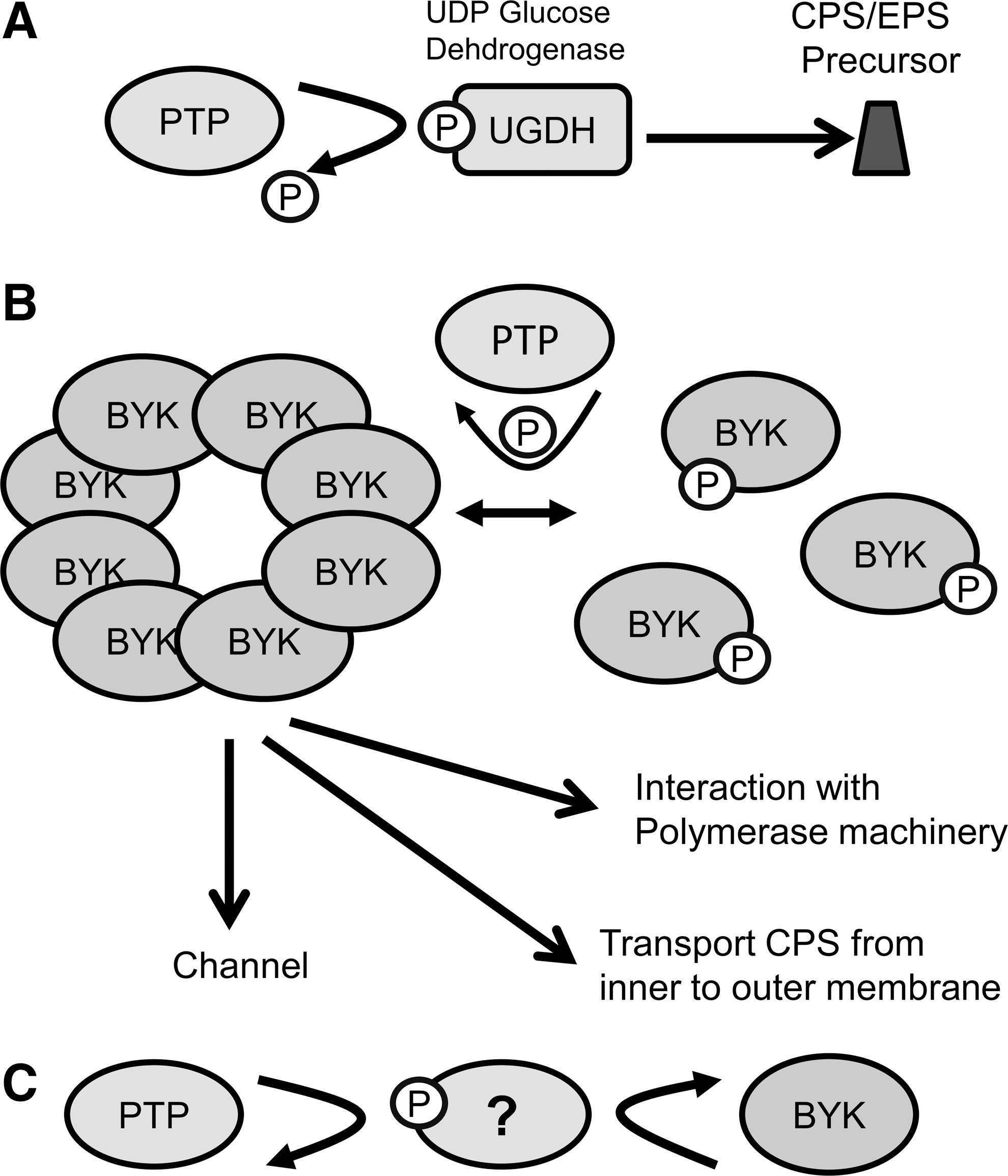
Whereas there is undoubtedly a link between bacterial PTPs and the regulation of CPS and EPS, the big question is how (summarized in Fig. 9)? YwqE from B. subtillus is another well-studied PHP PTP, which is linked to the regulation of a number of facets of bacterial physiology (46, 60 –62). YwqE dephosphorylates BY-kinase YwqD as well as a number of other substrates (61, 62). These include single-stranded DNA-binding proteins, SssA and SssB, which result in significant increases in binding of these proteins to DNA (61). Furthermore, YwqE and YwdD target two UDP-glucose dehydrogenases (UGDs) (YwqF and TuaD) (62), which are thought to play a role in the production of B. subtilis EPS (Figs. 8 and 9A). Interestingly, UGD from E. coli is phosphorylated by Wzc and dephosphorylated by Wzb (32), with this phosphorylation contributing to both polysaccharide synthesis (colonic acid) as well as polymixin resistance (50). Additionally, the same enzyme in S. aureus (CapO) is also tyrosine phosphorylated, although the consequences of this are not understood (33). It would be of considerable interest to investigate if such phosphorylation occurs in other bacteria, such as in S. pneumoniae, which possess a homologous protein as well as the conserved phosphorylated tyrosine from E. coli and B. subtilis UGD. However, it is worth noting that a recent study has called into question whether phosphorylation of UGD in E. coli K-12 effects its activity (58). Furthermore, not all bacteria or even all serotypes within a single bacterial species such as S. pneumoniae possess a UGD, suggesting that this mechanism is not universal.
Role of Wzc in transporting CPS across membranes
Whereas one particular role of PTPs is to control the phosphorylation and oligomerization of their cognate BY-kinase, what affect this has on the polymerization of the secreted polysaccharide is in question. To date, a number of alternative hypotheses have been proposed, such as phosphorylation of additional factors as discussed (Fig. 9). Structural studies have suggested that BY-kinases may form channels that could allow the polysaccharide polymer to cross the membrane (6). Furthermore, it has been shown that in gram-negative bacteria, the BY-kinase Wzc interacts with the outer membrane pore protein Wza to transport the growing polysaccharide chain from the inner to the outer membrane (15, 19), with this thought to be related to the phosphorylation state of Wzc and its ability to form higher order oligomers (Fig. 9B). However, in gram-positives, such a contribution of the BY-kinase is obviously not required due to the lack of an outer membrane. An alternative hypothesis is that this switch of the structure of the BY-kinases from an octomer to a monomer may influence the interaction of the BY-kinase, with components of the polysaccharide assembly complex thus influencing the synthesis of the CPS (18). Whereas Wzc and the transmembrane adaptor protein in gram-positive bacteria, such as CpsC and CapA, show homology to PCP proteins, a group involved in chain length regulation of LPS O-antigen (70), an effect on chain length is difficult to envisage for CPS due to the fact that deletions of the respective gene in general result in unencapsulated bacteria. Thus, identification of a complete mechanism for the effect of the PTP in the regulation of biosynthesis of CPS and EPS is still some time off.
LMW PTP Etp is tyrosine phosphorylated—what about other PTPs?
A recent publication on the impact of Etp and Etk from E. coli Type 4 on CPS synthesis provided the interesting observation that the PTP Etp is itself tyrosine phosphorylated (72). Interestingly, the authors illustrated that this phosphorylation occurred in the highly conserved DPY motif, similar to that seen in the DPYY motif in eukaryotic PTPs (84), and suggesting that this phosphorylation may occur across the family of LMW PTPs. Furthermore, a recent study of the interaction between Wzc and Wzb has shown that the DPY motif, and indeed this tyrosine, is critical for the interaction between the BY-kinase and PTP (104). Whereas in the case of Etp, PY did impede Etp phosphatase activity, mutation of the amino acid rendered the bacterium unencapsulated. A surprising result was that neither of the two E. coli BY-kinases identified to date (Etk or Wzc) was responsible for Etp phosphorylation, as even in the absence of both, Etp was phosphorylated. Thus, it seems likely an additional unidentified BY-kinase plays a role in the biosynthesis of E. coli Group 4 CPS, via its effect on the phosphorylation of Etp (72). As there are no other proteins showing significant homology to Wzc and Etk, it seems likely that this constitutes a novel BY-kinase. This work now promotes interest in whether other PTPs involved in the biosynthesis of polysaccharides are also tyrosine phosphorylated. One unanswered question is the topic of additional targets of the PTPs. Whereas a number have been identified and discussed above, it is likely that there are a large number of additional targets. In recent times, phosphoproteomic studies carried out across a range of bacterial species have led to the discovery of a range of proteins phosphorylated on serine, threonine, and tyrosine (56, 59, 94, 101). Whereas those phosphorylated on tyrosine constituted the smallest group, this is due, in part, to the fact that the methods used in these studies favor identification of more heavily phosphorylated amino acids, with tyrosine normally phosphorylated at low percentages.
It is likely that additional targeted studies will lead to the identification of a substantial number of proteins with PY, using methods such as those described by Condina et al. (2010) (16). Indeed, increased identification of PY has already been evident in investigations of E. coli. Whereas a phosphoproteomic analysis of the whole cell led to the identification of only seven tyrosines phosphorylated (55), a more defined study of only ribosomal-binding proteins found that 11 of these proteins were tyrosine phosphorylated (94). Furthermore, a recent study utilizing methods to solely investigate the tyrosine phosphoproteome of E. coli found a total of 342 tyrosine phosphorylated proteins (38). Thus, it seems likely that we are only on the very first cusp of the identification of the full spectrum of tyrosine phosphorylated proteins in bacteria. With PTPs and BY-kinases described in this review, the only ones described to date, it would seem likely that these would play a role in the phosphorylation levels of these additional proteins in vivo. This in turn would suggest that regulation of CPS and EPS biosynthesis is an incredibly complex regulatory system, comprising a large number of proteins. Thus, significantly more research is required before we completely understand the role of PTPs beyond their role in the regulation of the secreted polysaccharide synthesis. Nevertheless, their known effect on regulation suggests that they are attractive targets for the development of novel antimicrobials.
PTPs as a target for the development of novel antivirulence drugs
With their critical role in the biosynthesis of CPS and EPS, the described bacterial PTPs represent a novel target for the development of antivirulence drugs [as we have discussed in detail recently (22)]. Such drugs would be expected to interrupt biosynthesis of this crucial virulence determinant, rendering the pathogen susceptible to the host's immune system (22). It is thought that bacteria will be less likely to develop resistance against such antivirulence drugs compared with conventional antibiotics, as antivirulence drugs do not directly kill the pathogen (83). Of these PTPs described within this review, those belonging to the PHP family are perhaps the most attractive to target, due to their predominant presence in bacteria. Thus, it would seem more possible to design and/or discover compounds that are specific and do not show off target affects against eukaryotic PTPs. LMW PTPs, such as Wzb, share many similarities with eukaryotic-like phosphatases (36), including the catalytic mechanism, and thus may present difficulties to design compounds that specifically target the PTP of choice.
Our work has involved a high-throughput screen to identify inhibitors of the PHP family (Pfam PF02811) member PTP CpsB from S. pneumoniae (96). This study led to the identification of a compound (fascioquinol E; FQE) that inhibited CpsB PTPase activity in vitro, and also in vivo inhibited phosphorylation of the cognate BY-kinase CpsD and pneumococcal CPS biosynthesis. Interestingly, FQE also inhibited the growth of S. pneumoniae as well as another gram-positive human pathogen S. aureus. This led to our hypothesis that the compound was a general PHP family inhibitor (3). This family comprises a multitude of subfamilies, including the DNA polymerase PolC from S. pneumoniae. Interestingly, it has been found that the PHP domain from PolC is essential in S. aureus (42). Recently, we showed that FQE was able to inhibit the catalytic activity of the PHP domain of PolC in vitro (95), providing further evidence for this hypothesis. Other notable PHP family proteins include the other DNA polymerase PolX and DnaE as well as histidinol phosphatase, which provides a critical step in the histidine biosynthesis pathway. In a number of DNA polymerases (PolX and PolC) (4, 98), the PHP domain has been shown to possess exonuclease activity, although this is not always the case (23) with some postulation that it serves nonenzymatic roles also, such as maintenance of the polymerase structure (5). We are currently investigating if FQE can inhibit the functions of additional PHP domain proteins, amid promise that the domain is a novel target for the development of a new breed of antibiotics, which also have the potential to also act as antivirulence drugs.
Interestingly, if specific inhibition of these PTPs was possible, leading to unencapsulated bacterium, it is possible that such treatment could lead to protection from subsequent infections. For S. pneumoniae, it has been shown that unencapsulated bacteria linger in the nasopharynx of mice for some time (74). Interestingly, the use of such strains as live-attenuated vaccines has provided significant protection against subsequent infection, both from the same and different capsular serotypes (85). Thus, it is interesting to hypothesize that any drug that inhibits these PTPs could also lead to protection against subsequent infection.
Conclusion
With the ever-increasing rates of antibiotic resistance, we are creeping dangerously close to returning to a preantibiotic era of untreatable bacterial infections. Thus, the search for novel antimicrobial targets is of the utmost importance. CPS and EPS are critically important virulence factors for a range of major bacterial pathogens. For a number, bacterial PTPs are critically important enzymes in the regulation of their biosynthesis, and as such are attractive targets for the development of novel antimicrobials. However, the precise role that these PTPs play, both in the regulation of this biosynthesis as well as in wider bacterial physiology is only just beginning to be uncovered. Indeed, it is likely that in the coming years, a much broader range of substrates for these PTPs will be identified, as more research into the role of bacterial tyrosine phosphorylation is conducted. Such work is critical, such that we can gain a greater insight into the regulation undertaken by these PTPs, not just regulation of CPS and EPS biosynthesis, but also of further features of bacterial physiology.
Footnotes
Acknowledgments
This work was supported by grants from the NHMRC (Program grant (565526) and Project grant (1048749)), and also a grant from the Channel 7 Children's Research Foundation.