
Abstract
In their letter, Pagano et al. appreciate the development of the Nox1, Nox2, and Nox4 triple (3N−/−) knockout mouse. They also agree on the view that chemiluminescence assays in general have severe limitations. However, they criticize the fact that the membrane assays in the particular study were restricted to chemiluminescence techniques. Moreover, Pagano et al. got the impression that statements concerning membrane assays of Nox activity in general were made. In addition to a lack of some technical details, Pagano et al. also found the characterization of the 3N−/− incomplete and some of the results to be incomprehensible. Although we are grateful for the interest of Pagano et al. in our work, we realized that basically each observation of our study was questioned. This is certainly an excessive rejection of the study in total and fails to appreciate the clear chain of evidences presented. Our work focused on chemiluminescence, and thus, any conclusions are restricted to this technique. Moreover, the 3N−/− mice were never developed to study the physiology of Nox enzymes, but rather to validate Nox specificity of NADPH-stimulated chemiluminescence assays. We are convinced that our findings are a valid demonstration that chemiluminescence-based assays in membrane preparations stimulated with NADPH do not measure Nox activity. This conclusion is based on both overexpression studies as well as genetic deficient mouse models. The criticisms of Pagano et al. thus might be justified in some aspects; they, however, cannot disprove the conclusions of our work. Antioxid. Redox Signal. 23, 1247–1249.
W
Imposed by the particular News and Views format of the journal, certain brevity had to be applied to our article. This is reflected by the absence of a discussion and the insufficient coverage with references. We are grateful that the letter by Pagano et al. provides a basis to enter into a discussion on how Nox activity should be measured.
The focus of our study was on the use of enhanced chemiluminescence in membrane assays stimulated with NADPH. In that particular context, we found little evidence that Nox NADPH oxidases contribute to the signal obtained. This conclusion was based on two different experimental results: genetic deletion of Nox1, Nox2, and Nox4 all together had no impact for most of the assays on the signal obtained and overexpression of Nox4 and Nox5 in cultured cells did increase the signal in intact cells, but not in the NADPH-stimulated membrane fraction. Nevertheless, our observations indeed are restricted to chemiluminescence assays and the particular experimental setup. This is also reflected by the title of our work in which, specifically, the chemiluminescence technique is mentioned. We cannot and did not make statements about the signal of membrane preparations measured by ESR or cytochrome C. Nevertheless, such specialized techniques to measure Nox activity have never gained broad utility. Moreover, whether the word gold standard should be applied to those assays if they are used in the presence of NADPH can also be argued.
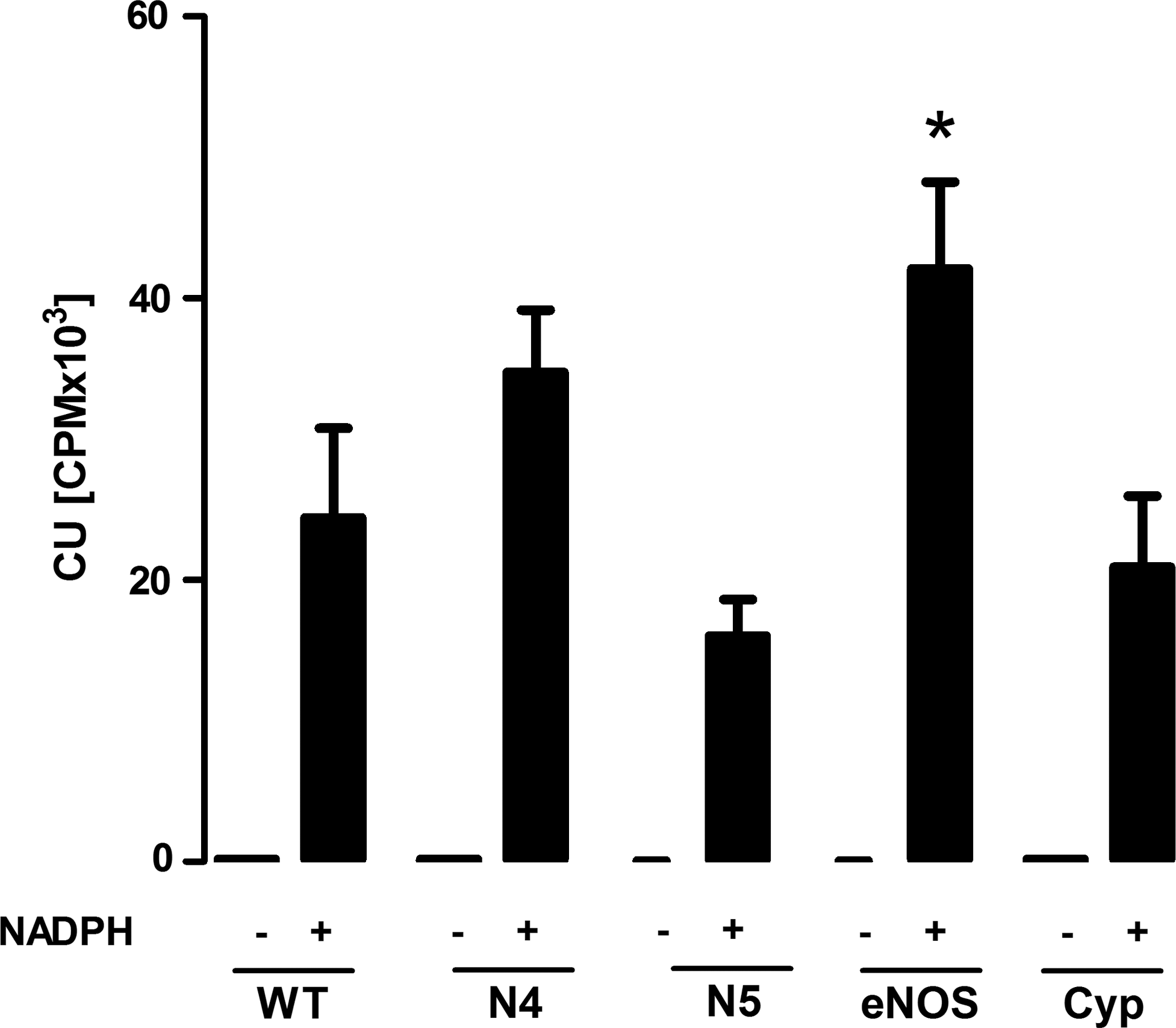
Regarding the criticisms applied to the assays performed in homogenates of HEK293 cells, Pagano et al. ask for the basal values of membrane preparations; these values are now exemplified for the lucigenin measured in Figure 1 of this letter. In the absence of NADPH, the signal was basically below the detection limit of our system in all assays and, therefore, not shown in the original article. Moreover, Pagano et al. criticize that the basal level of Nox and eNOS was not provided for HEK293 cells. These data were not included as we are unable to detect Nox or eNOS protein in HEK293 cells by Western blot. We cannot follow the argumentation that omitting this information in the original article disqualifies our observations, that is, HEK293 cells once overexpressing Nox enzymes generate a Nox-dependent chemiluminescence signal, which is not present in NADPH-stimulated membrane preparations of the very same cells. Moreover, the argument that HEK293 cells present an invalid heterologous system ignores that we supported those findings in cells and tissue data with native Nox4 expression: kidneys and fibroblasts (Figs. 4 and 5 of the original article). Finally, it is simply wrong to state that we attributed the eNOS-derived chemiluminescence in the presence of NADPH to superoxide formation. This is by no word stated in the article.
Pagano et al. criticized the use of the serine phosphatase inhibitor okadaic acid in our article by quoting an irrelevant study demonstrating that pretreatment of neutrophils with excessive concentrations of that compound prevents zymosan-induced Nox activation in leukocytes (2). In our current work, okadaic acid was present only during cell lysis to prevent the loss of p47phox serine phosphorylation. This was used instead of phenylmethylsulfonyl fluoride (PMSF), which is similar to AEBSF and may therefore inhibit Nox activity (6).
As another second aspect, Pagano et al., criticized an, in their understanding, insufficient characterization of the triple knockout mice. By ignoring the vascular function data and the results on heart to bodyweight ratio, which clearly showed an effect of angiotensin II, they specifically criticize that vascular morphometry was not carried out. As also mentioned in the article, we never intended to perform an in-depth physiological characterization of the triple knockout mice. Rather, we felt that it was sufficient to demonstrate some, but not all, of the changes established in the single knockouts, that is, lack of phorbol myristate resonance (PMA) induced Reactive oxygen species (ROS) production, altered endothelial function, and changed cardiac hypertrophy. A triple-knockout mouse is a highly artificial model which, at least in our opinion, should rather not be used to study physiology. For the present study, it was only a tool for our analysis of the ex vivo chemiluminescence assays.
The criticism that we failed to obtain differences in basal lucigenin-enhanced chemiluminescence between wild-type mice treated with and without AngII in the absence of NADPH is valid. Since the introduction of the low concentration of lucigenin (5 μmol/L) in the standard protocol, we are basically unable to detect this difference in normal mice with our machine. The signal is too low and the high extracellular superoxide dismutase (ecSOD) activity in mice (6) (other than in rats) is one of the causes for this. The statement of Pagano et al. that numerous groups have succeeded in showing increased chemiluminescence in mouse aortic rings after in vivo treatment with angiotensin II using 5 μmol/L lucigenin cannot be derived from the current literature.
Pagano et al. speculate that a potential induction of Nox3 might compensate for the lack of the other Nox enzymes. Indeed, in the presence of AngII, we cannot exclude this possibility; however, as Nox3 was not changed under basal conditions, we feel that this is somewhat unlikely, in particular, as a great impact of Nox3 to vascular ROS production even in pathology still needs to be established (3). Potentially, global p22phox knockout mice should be studied to ultimately address this aspect.
As clearly stated in the introduction of our work, the focus of our study was NADPH-stimulated chemiluminescence assays. On this basis, we selected what we felt was the appropriate methodology for the current work. We do not understand why the lack of inclusion of mice knockout for each individual Nox enzyme is a paramount concern. In the chemiluminescence assays in homogenates, control plasmid (WT) and 3N−/− were basically indistinguishable. We disagree that such a finding necessitates studying all individual knockout mice, of which the 3N−/− was generated.
Our work is certainly not discrediting the vascular Nox field. It is, however, a reflection of the prevalent difficulties researchers have when trying to assess Nox activity in membrane assays. Given the wide availability of microplate readers and the strong signal generated by chemiluminescence enhancers, these assays are the first to be performed when groups enter the field. Those groups are not aware of the fact that the signers of the letter and Prof. Pagano have moved on to use a far more advanced technology to assay the Nox activity in membranes (4). In fact, we have been contacted by numerous people from academia and the industry complaining about their inability to measure the Nox activity by the chemiluminescence technique. Also Prof. Pagano and the other opinion leaders cosigning his letter have to acknowledge this problem.
The new groups entering the field are not aware of the fact that poorly understood signal amplification either by NADPH or even more surprisingly NADH, high concentration of the enhancer or high pH like in the Krebs buffer (1), was needed by some undefined chemistry to eventually result in a Nox signal in membranes. That this signal is largely diphenylene iodomium (DPI) and superoxide dismutase (SOD) insensitive and not quantitative has been widely recognized and ignored. We are not at all denying that those data, to which our group also contributed, have been instrumental in establishing fundamental observations in the Nox field. Nevertheless, it is also true that at least the authors of the so heavily criticized article never understood how the Nox NADPH oxidases generate that amplified chemiluminescence signal precisely.
We cannot deny that we are frustrated that there is no robust and easy assay to measure Nox-dependent ROS production in membranes. We are sure that Pagano et al. also share this view and that they agree that electron paramagnetic resonance (EPR) and cytochrome C in membranes stimulated with NADPH are also rather difficult assays restricted to specialized laboratories. Nevertheless, we felt that it was important to express our concerns regarding the use of chemiluminescence in an interesting and stimulating set of data. We agree with Pagano et al. that our study has limitations. We are, however, convinced that the essential conclusion of our work, that is, the signal obtained with NADPH-stimulated chemiluminescence in membrane preparations is not primarily Nox derived and cannot be disproved by the concerns raised by Pagano et al.