
Abstract
Introduction
T
The importance of the lymphatic vasculature in human diseases is being increasingly recognized. For example, lymphatic vessels frequently serve as a route for tumor metastasis [reviewed in Stacker et al. (109)]. Moreover, they actively regulate inflammation through both the transport of antigens and their presentation [reviewed in Card et al. (22)]. Impaired function of the lymphatic vessels leads to lymphedema, a debilitating tissue swelling, which develops most frequently either after filarial parasitic infection, common in developing countries [reviewed in Nutman (87)], or following iatrogenic tissue damage, affecting a significant proportion of cancer patients after lymph node resection or irradiation (82).
In addition, mutations in genes important for lymphatic vascular development and maintenance underlie different subtypes of hereditary lymphedema, for example, VEGFR-3 in Milroy disease or FOXC2 in lymphedema-distichiasis (LD) [reviewed in Mortimer and Rockson (81)].
Importantly, recent studies suggested that lymphatic vascular dysfunction contributes to a significantly broader array of human diseases than previously thought and may include such prevalent conditions as obesity, hypertension, and atherosclerosis [reviewed in Coso et al. and Mortimer and Rockson (29, 81)].
Lymphatic Vascular Development in Mammals
During mouse embryonic development, the majority of lymphatic endothelial cells (LECs) are produced by budding of the venous endothelium [reviewed in Schulte-Merker et al. (105)]. In parallel, other vascular beds or even nonendothelial cell types can contribute to the developing lymphatic vasculature (11, 61, 76, 110).
A leading event that triggers LEC commitment is the upregulation of prospero-related homeodomain transcription factor (Prox1), which is essential for LEC programming and further maintenance of LEC fate (47, 52, 93, 114). Prox1 induces the expression of vascular endothelial growth factor receptor −3 (Vegfr-3), which is important for LEC survival, migration, and proliferation (47, 93, 107). LEC progenitors then migrate out from the cardinal vein following a gradient of Vegfr-3 ligand, Vegf-c, and assemble into primordial lymphatic vessels, which remain connected to the cardinal vein to provide a return path for the lymph to the blood circulation [reviewed in Schulte-Merker et al. (105)]. To avoid blood flow into the lymphatic network, the connections between blood and lymphatic vasculatures are secured by lymphovenous valves (108).
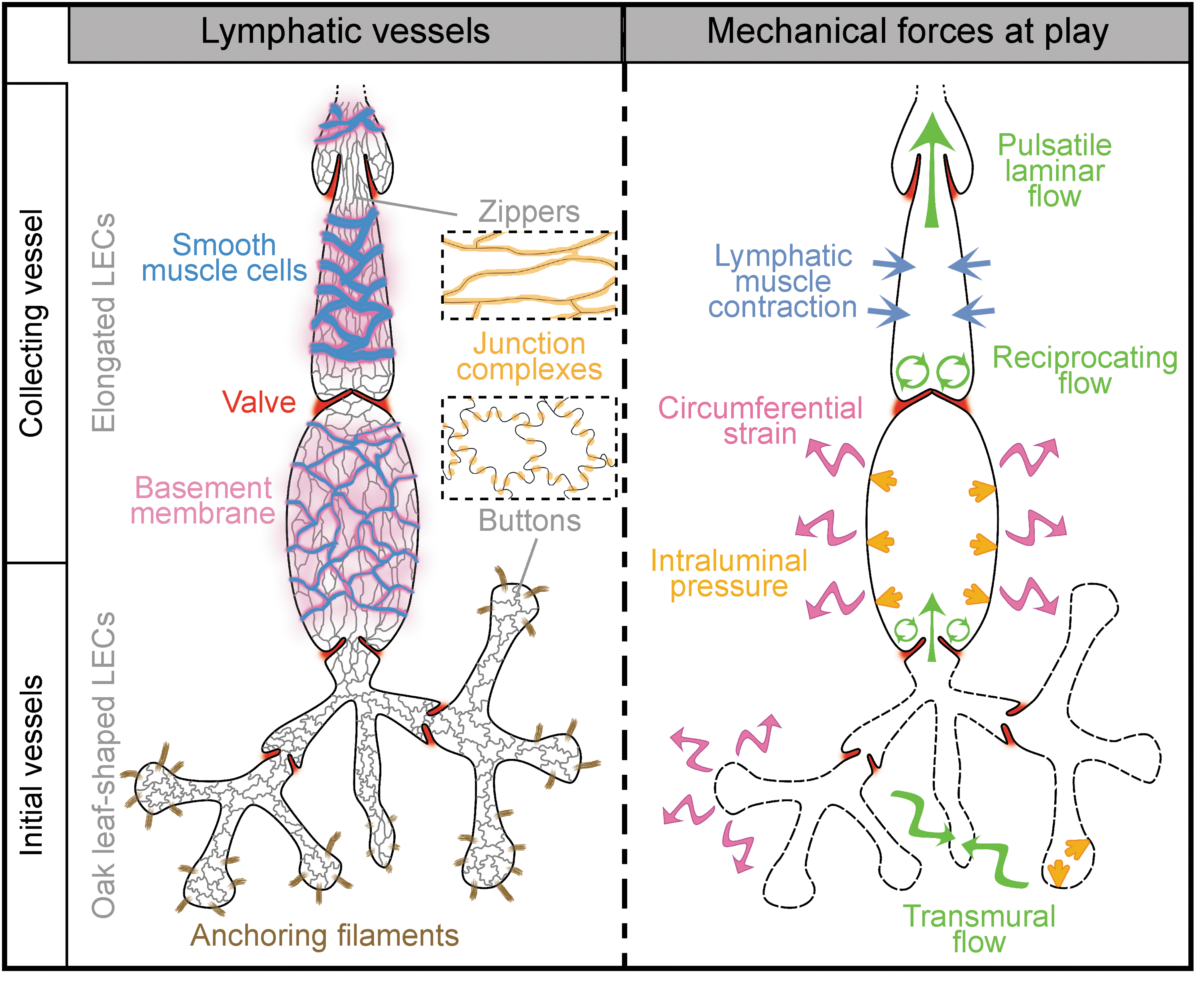
The primordial lymphatic plexus is further remodeled into two mature compartments, initial and collecting vessels, which have distinct structure and function [reviewed in Schulte-Merker et al. (105), Fig. 1]. The time of lymphatic vessel maturation onset varies from one tissue to another, for example, E14.5 in mesenteric collecting vessels or P10 in the ear dermis (74, 86).
Lymphatic Vessel Structure and Function
Initial vessels and lymph formation
Initial lymphatic vessels consist of blind-ended capillaries connected to a branched network of precollectors (Fig. 1). They lack a continuous basement membrane and mural cells. The thin endothelium consists of LECs interconnected via discontinuous, overlapping cell–cell junctions. Such button-like junctions are important features of the initial vessels as they represent the sites for entry of fluid and immune cells [(7), Fig. 2A]. LECs are also directly connected to the extracellular matrix (ECM) via anchoring filaments, which support the expansion of lymphatic capillaries in conditions of increased interstitial pressure [(65), reviewed in Schulte-Merker et al. (105)].
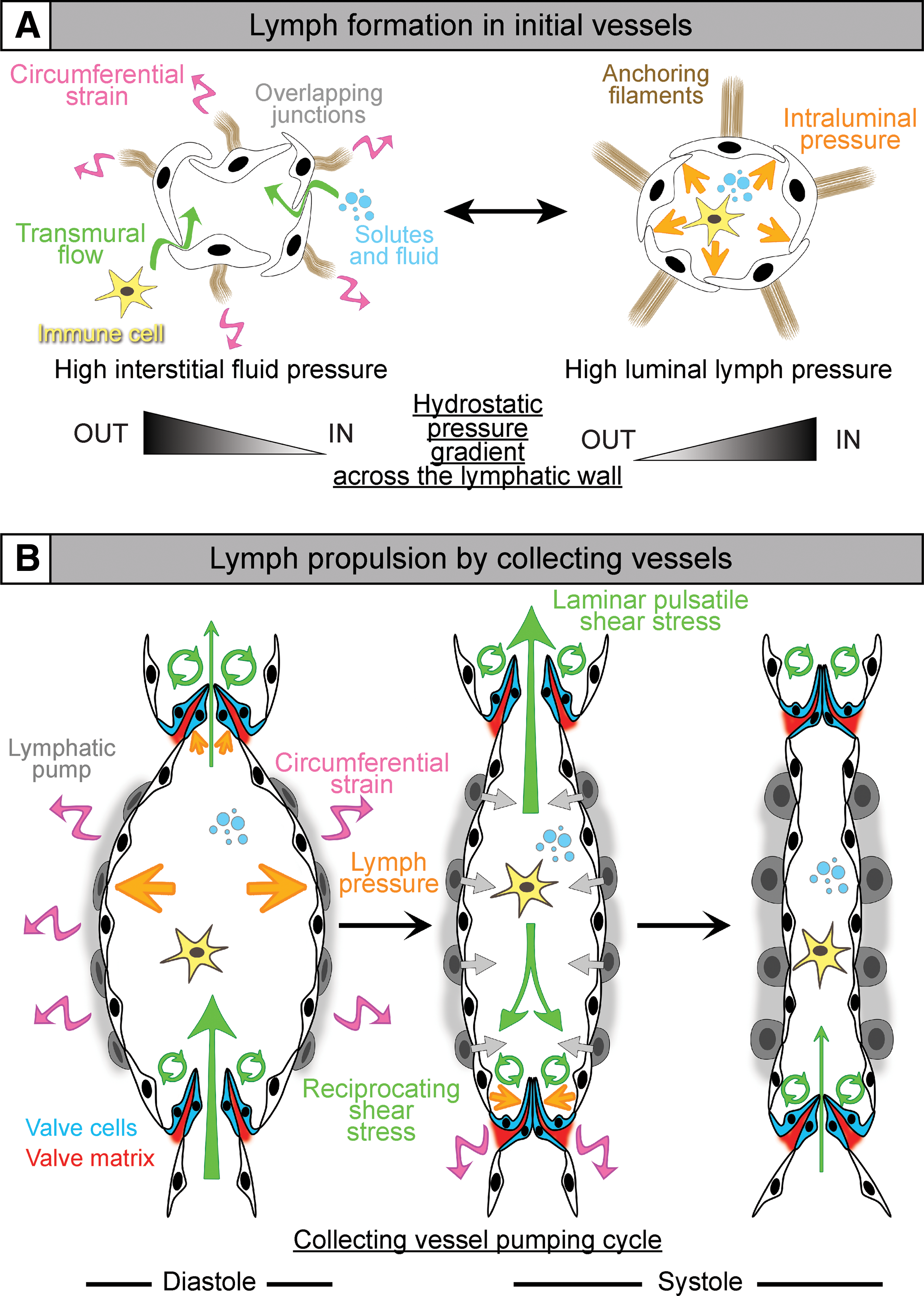
The overlapping cell–cell junctions function as primary valves, where the local hydrostatic pressure throughout the vessel wall determine their opening or closure. Fluid accumulation in the interstitium opens the overlapping junctions by stretching the anchoring filaments, facilitating transport of water, solutes, particles, and immune cells into the lumen (Fig. 2A, left panel). Once the vessel is filled, the increasing luminal pressure seals the overlaps, thus preventing lymph escape (Fig. 2A, right panel) [review in Schmid-Schonbein (104)]. Lymph then circulates to collecting vessels.
Collecting vessels and lymph transport
Different from capillaries, collecting vessels consist of a series of contractile segments, the lymphangions, covered with a basement membrane and smooth muscle cells (SMCs) and interspaced by intraluminal lymphatic valves [reviewed in Schulte-Merker et al. (105)]. Collecting vessel LECs have continuous zipper-like intercellular junctions that prevent lymph leakage [(7) and Fig. 1].
Propulsion of the lymph in collecting vessels is ensured by the coordination of two main forces: the intrinsic pump, which is triggered by contraction of the lymphatic muscle in response to an increase in the luminal lymph pressure (Fig. 2B), and the extrinsic pump, which depends on the activity of the surrounding tissues (e.g., heart beats or respiratory movements). Intersegmental valves open or close in phase with the lymphangion contractions and ensure unidirectional lymph flow (Fig. 2B). Accumulation of the lymph in a lymphangion raises the luminal fluid pressure and pushes the valve leaflets toward each other, thus closing the valve and preventing lymph backflow. Further increase in lymph pressure stretches the endothelium and activates the lymphatic pump, which opens the valve and propels the lymph to the next lymphangion (12, 117).
Biomechanical Forces in the Lymphatic Vasculature
The lymphatic vasculature is shaped and maintained by a variety of biochemical signals, such as gradients of growth factors [e.g., Vegf-c/Ccbe1/Adamts3 cascade (15, 51, 63)], guidance molecules [Nrp1/Sema3a/PlexinA1 (16, 53)], and chemokines [Ccl2/Ccr2/Ackr2 (66)]. In addition, both developing and mature lymphatic vessels are subjected to diverse biomechanical forces generated by circulation of the lymph or by strain applied on the vessel wall, both by lymph mechanics and external mechanical forces.
Lymph flow
Two different types of flow prevail in the lymphatic vascular network. Mostly present in initial lymphatics, transmural flow results from lymph formation when fluid is transferred through the capillary endothelium from the interstitial space to the vascular lumen, following pressure gradients [reviewed in Chiu and Chien (26)]. It depends on the endothelium integrity and is increased during, for example, inflammation, where proinflammatory cytokines weaken endothelial cell–cell junctions (75, 80).
In collecting vessels, the endothelium is less permeable and is mostly subjected to higher luminal flow, which generates shear stress of the lymph on the luminal side of endothelial cells (Fig. 2B). Two distinct types of shear stresses are found in the lymphatic collecting vessels: (i) a laminar pulsatile shear stress, present in the linear part of lymphangions, as well as on the luminal side of the valve leaflets, and (ii) a reciprocating shear stress in the valve sinus, where lymph flow is more turbulent due to a conjunction of forward and retrograde fluid movements (117).
Shear stress has been extensively studied over the past decades in blood vessels and disturbed shear stress has been identified as an important trigger of endothelium inflammation such as observed in atherosclerosis [reviewed in Chiu and Chien (26)]. The insights in the role of shear stress in lymphatic vessels will be discussed later in the chapter.
Cyclic strain
Vessel wall stretch results from circumferential expansion of both capillaries and collecting lymphatic vessels during fluid uptake and transport, respectively (Fig. 1A, right panel). In initial lymphatics, fluid entry into the capillary lumen leads to expansion of the capillary endothelium, which generates tensional forces via anchoring filaments. Collecting vessels are subjected to cyclic strain due to regular increase of the intraluminal pressure generated by valve closure and lymph accumulation in lymphangion [reviewed in Zawieja (117)], resulting in active contractions of the lymphatic muscle, which propels the lymph forward.
External mechanical factors
Lymphatic vessels also experience exogenous wall strain locally as a result of organ function, such as during breathing or cardiac contractions [reviewed in Zawieja (117)]. Another recently identified example of external forces is the lymph absorption and propulsion in lacteals, the blind-ended, finger-shaped lymphatic capillaries in intestinal villi, which is driven by active contraction of the villus muscle and plays an important role for dietary fat drainage (10, 27).
Recently, several studies interrogated the contribution of such mechanical forces to lymphatic vascular development and function, which will be the scope of our review with greater emphasis on shear stress in lymphatic vessels.
Molecular Mechanisms of LEC Responses to Biomechanical Forces
Mechanical forces in early lymphatic vascular development
Vegf-c/Vegfr-3 signaling plays a critical role in early lymphatic vascular development and targeted inactivation or loss-of-function mutations in these genes leads to severely impaired embryonic lymphangiogenesis and lymphedema in humans (41, 56, 57).
A study by Planas-Paz et al. investigated whether increased interstitial fluid pressure (IFP) and ensuing cell stretching provide an additional cue for the early steps of lymphangiogenesis (Fig. 3). Indeed, rise of IFP at E12 occurs in parallel with the expansion of primordial lymphatic vessels and activation of Vegfr-3 signaling (94). Loss-of-fluid experiments in E11.5 mouse embryos showed that decreasing IFP during this critical period prevented LEC elongation/stretch, diminished Vegfr-3 phosphorylation, and resulted in decreased LEC proliferation (94). On the other hand, augmenting IFP in gain-of-fluid experiments through the injections of phosphate buffer saline into the jugular region of E11.5 embryos led to further LEC elongation, increased phosphorylation of Vegfr-3, and expansion of the lymphatic vasculature (94).
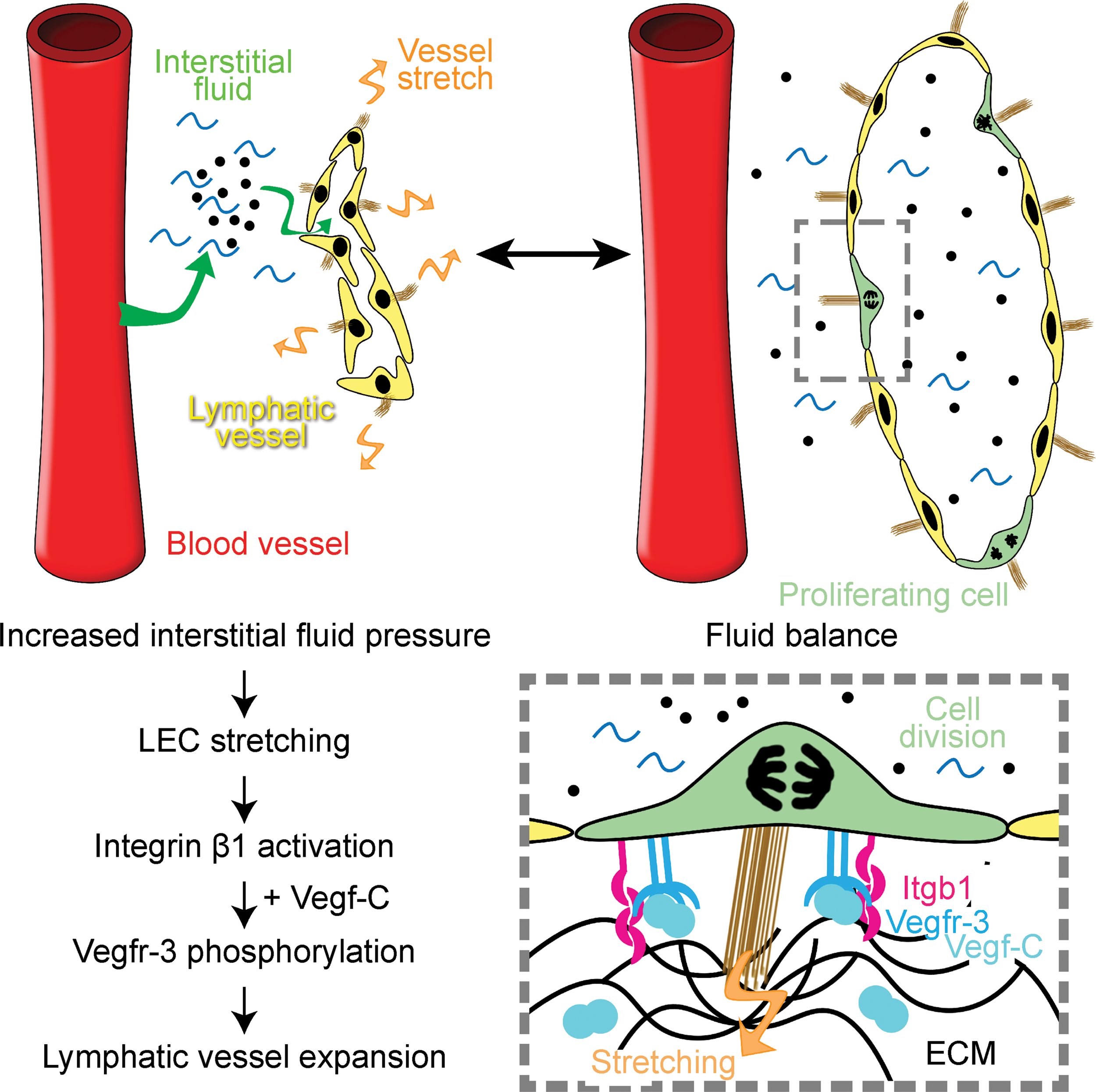
Mechanistically, stretch-induced activation of VEGFR-3 in LECs was mediated by β1 integrin, possibly via Src activation. These pioneering studies thus demonstrate how combined action of biochemical signaling (VEGF-C) and mechanical forces (stretching) synergistically activates key lymphangiogenic VEGFR-3 signaling, linking tissue demand for fluid drainage with expansion of the lymphatic vasculature.
Fluid shear stress and morphogenesis/maintenance of lymphatic valves
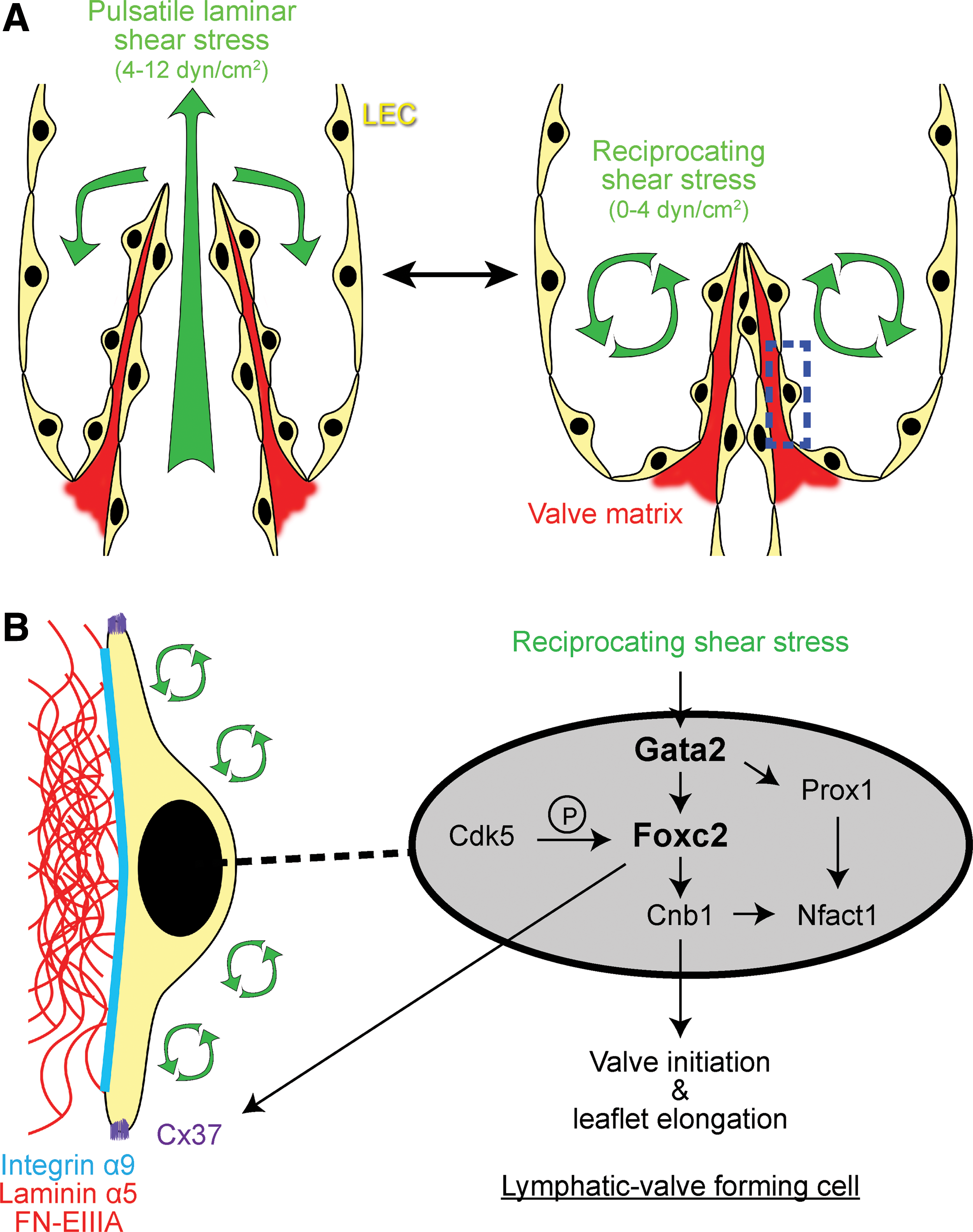
Lymphatic valves are specialized structures in collecting vessels that prevent lymph retrograde flow (Fig. 1). They are usually bileaflet, with each leaflet comprising an ECM core covered on both sides with specialized LECs. Lymphatic valve endothelial cells express a distinct subset of markers in comparison with the neighboring lymphangion cells (Fig. 4): they are characterized by high levels of transcription factors, Prox1, Foxc2, Gata2, and Nfatc1 (60, 86, 100), ECM receptors and components, such as Itga9, laminin alpha5, FN-EIIIA, and emilin-1 (8, 30), and other signaling molecules, such as ephrinB2-EphB4 (78, 118), Vegfr-3 (73, 86), Notch1 (84), Tie1 (97, 106), angiopoietin-2 (106, 119), gap junction proteins Cx37, Cx43, and Cx47 (55, 100), and planar cell polarity proteins, Celsr1 and Vangl2 (112).
Analyses of knockout mice demonstrated that many of these molecules are required for various stages of valve formation (Table 1). Transcription factors, Foxc2 and Gata2, appear to be at the top of the hierarchy as Foxc2 −/− mice display complete agenesis of lymphatic valves and do not form lymphatic collecting vessels, and Gata2 is important for the induction of Foxc2, at least in vitro (59, 86, 92, 111). In mouse embryos, development of lymphatic valves is initiated around E15.5 when the lymphatic valve-forming cells, characterized by high expression of the transcription factors, Foxc2 and Prox1, can be identified in the remodeling primary plexus (8, 86, 100).
n.d., not determined; ECM, extracellular matrix; LEC, lymphatic endothelial cell; SMC, smooth muscle cell.
Localization at the sites of lymphatic branching points and bifurcations suggests an important role of disturbed flow patterns in the development of valves (54, 100). Indeed, valve initiation timing in mouse embryo parallels the onset of active lymphatic drainage, and culture of embryonic collecting vessels in the absence of flow resulted in rapid regression of the valves (100).
Importantly, cultured LECs exposed to oscillatory shear stress (OSS) to mimic the recirculating flow acquire many characteristics of lymphatic valve-forming cells, including increased expression of FOXC2 and GATA2 (59, 100). Knockdown of GATA2 or FOXC2 leads to reciprocal loss of induction by FOXC2 or GATA2 by shear stress [(59) and Agalarov Y., unpublished observation], suggesting the presence of transcriptional feedback mechanisms important for valve initiation. Alternatively, it is possible that loss of any of the key valve transcriptional components (Prox1, Gata2, Foxc2) alters sensing of shear stress by LECs and prevents any subsequent response to shear.
In vitro studies of LECs exposed to OSS identified some of the molecular links between mechanical forces and signaling pathways involved in the valve formation process. In addition to the induction of FOXC2 and GATA2 expression, OSS also elevates expression of the gap junction protein, CX37, and activates Ca2+/calcineurin/NFATc1 signaling. Importantly, such responses of LECs to shear stress are dependent on the presence of PROX1 and FOXC2, demonstrating a cooperative process between mechanical forces, transcriptional regulation, downstream gap junctional communication, and Ca2+ signaling in the development of lymphatic valves (100, 102) (Fig. 4B).
An additional layer of regulation includes the control of Foxc2 activity by phosphorylation: Foxc2 contains a 70-aa phosphorylation domain harboring 7 proline-directed protein kinase phosphorylation sites, which is important for Foxc2 recruitment to the chromatin in LECs (48). Serine/threonine kinase Cdk5, previously noted for its role in neuronal development and neurodegeneration (25), was recently identified as one of the upstream kinases of Foxc2, important for Foxc2 target gene expression under shear stress conditions (71).
Termination of lymphangiogenic Vegfr-3 signaling is another prerequisite for transformation of capillary-like primary lymphatic plexus into collecting vessels and formation of valves. Laminar shear stress, present in lymphangions (117), and likely also during the onset of collecting vessel formation before valve development (100), may play a role in this process through the induction of epsin1 and 2, ubiquitin-binding adaptor proteins involved in endocytosis, and termination of Vegfr-3 signaling (73). Indeed, developing collecting lymphatic vessels progressively reduce their levels of Vegfr-3 (86), while the expression of epsin 1 and 2 is increased (73), and mice deficient in epsin 1 and 2 fail to develop mature collecting lymphatic vessels and valves (73). However, the significance of Vegfr-3 signaling in lymphatic valve leaflet endothelial cells still remains to be understood as inactivation of Vegfr3 in LECs does not affect postnatal lymphatic valve development (116).
Compartmentalization of the lymphatic vasculature into capillaries and collecting vessels is essential for efficient lymph transport. A fundamental but also clinically important question is how such organization is maintained throughout life.
Continuous Gata2 expression is important for maintenance of the postnatal lymphatic valves: mice without Gata2 in LECs have malformed valves, which are also reduced in numbers, and mispatterned collecting vessels (59).
Inducible postnatal inactivation of Foxc2 revealed that this transcription factor is critical for lifelong maintenance of the lymphatic collecting vessels (101). Foxc2 lecKO mice display rapid valve degeneration, loss of other collecting vessel characteristics, and develop fully penetrant lethality due to extensive lymph leakage (101). Mechanistically, Foxc2-deficient LECs are characterized by disrupted cell–cell junctions and a disorganized cytoskeleton, leading to an abnormal response to shear stress, cell hyperproliferation, and apoptosis. Transcriptome analyses demonstrated that Foxc2 controls the expression of approximately 35% of the shear-responsive genes in LECs. Interestingly, aberrant proliferation of Foxc2-deficient LECs in response to shear stress was mediated by increased Yap1/Taz signaling, previously shown to regulate mechanosensitive responses in other cell types (34, 101).
Effects of shear stress on the lymphatic endothelium
Many studies investigated the responses of blood endothelial cells to shear stress; however, there are only a handful of reports to date, which compared them with LECs. In many aspects, the responses of these two endothelial subtypes are similar, for example, both blood and LECs align in the direction of flow in laminar shear stress, activate Ca2+ signaling, nitric oxide (NO) production, and display nuclear export of NF-kB [e.g., (5, 13, 14, 58, 85, 100)].
Recent work of Baeyens et al. analyzed in cultured blood and lymphatic endothelial cells the concept of fluid shear stress set point, which postulates the existence of a preferred range of shear stress for efficient vascular responses (21). In line with different levels of shear stress experienced by blood or lymphatic endothelium, the set point of human umbilical vein endothelial cells ranged between 10 and 20 dyne/cm2, as determined by cell alignment and NF-kB translocation, whereas LECs were considerably more sensitive with a set point of 4–10 dyne/cm2, which is in the range of shear stress values observed in the rat mesenteric lymphatic collecting vessels (117). Laminar shear stress induced phosphorylation of VEGFR-3 in LECs and high VEGFR-3 levels increased the sensitivity of endothelial cells to shear (5), suggesting that VEGFR-3 levels represent one of the determinants of the shear set point in different vascular beds (Fig. 5).
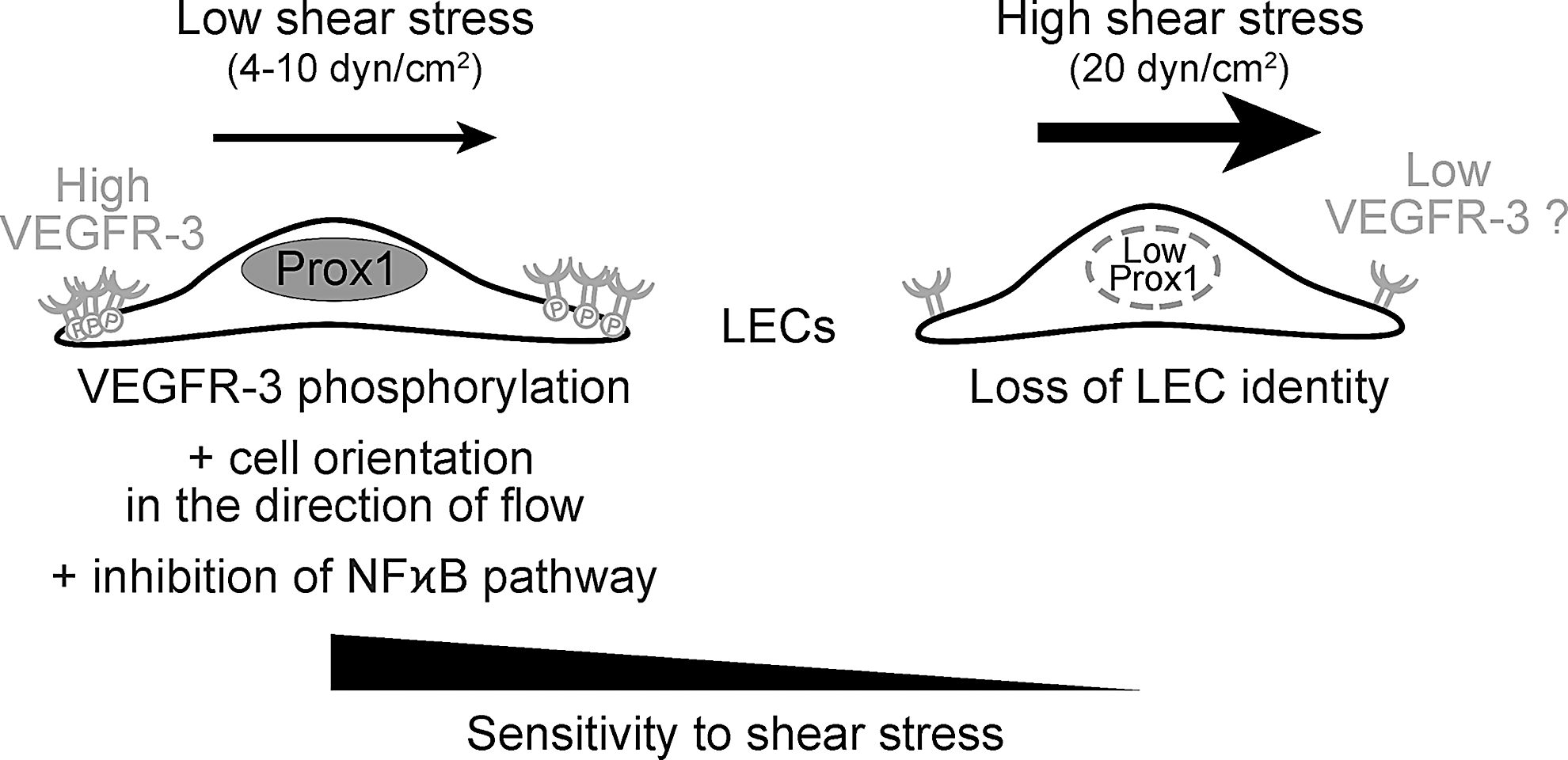
Even low 0.5 or 1.0 dyn/cm2 shear stress induced release of ATP from cultured LECs, which further led to increased production of endothelial nitric oxide synthase (eNOS) via activation of the purinergic P2X/2Y receptor and enhanced Ca2+ signaling (58). Shear stress-dependent NO production by eNOS in LECs allows coordination between shear stress and lymphatic pumping by regulating the contractile activity of SMCs.
High sensitivity of LECs to low levels of shear stress is therefore important to adapt the lymphatic function to the needs of tissue draining, thus maintaining the body fluid balance.
Shear stress control of endothelial cell identity
Arterial versus venous endothelial cell identity is not only encoded genetically but it can also be modified by the level and the type of shear stress (20, 62, 64). For example, local manipulations of blood flow in the developing chick embryo led to reversal of arterio-venous differentiation in accordance with the new flow type (64). Furthermore, in vitro exposure of cultured endothelial cells to pulsatile shear, found in arteries, was more efficient in inducing the expression of arterial markers than constant shear found in veins (20). Mechanistically, shear stress seems to impart arterial identity by increasing Notch signaling (49, 77).
Interestingly, high shear stress, exceeding the physiological range experienced by lymphatic vessels, reduces the expression of PROX1, a key regulator of LEC identity, and VEGFR-3 expression (23, 93, 107) (Fig. 5).
Mice that lack hematopoietic adaptor protein Slp76 implicated in T-cell receptor and other signaling pathways have impaired lymphatic development due to abnormal separation of newly established lymphatic vessels from the blood vasculature. It is possible that excessive shear stress, such as observed in lymphatico-venous shunts in Slp76-deficient mice, may shift the identity of lymphatic vessels toward a blood vascular phenotype and perhaps further stabilize this phenotype by increasing the vascular set point (23).
Interestingly, at the same time, mice deficient in Clec-2, a platelet receptor involved in the binding of platelets to LECs that are phenotypically similar to Slp76 knockout mice, also display reduced lymph flow because of blood backflow into lymphatic vessels. Such animals have impaired lymphatic valve development, further demonstrating the critical role of shear stress in lymphatic vessel remodeling during development (111).
Maintenance of the lymphatic capillary function
Mature lymphatic capillaries are characterized by a fragmented and thin basement membrane and discontinuous, button-like cell–cell junctions (Fig. 2A). Formation of button junctions begins during late development and continues postnatally in parallel with increased animal activity and, consequently, fluid transport (115). The mechanisms of such junctional specialization are not completely understood yet; however, button junction formation is promoted by glucocorticoid receptor signaling (115) and requires angiopoietin-2 and VE-cadherin phosphorylation (45, 119). Functionally, discontinuous junctions in initial lymphatic vessels are important for ensuring easy access of interstitial fluid and immune cells to the lymphatic capillary lumen.
Interestingly, such transmural flow itself affects permeability and organization of the LEC-cell junctions, as well as the expression of cell adhesion molecules and chemokines. In vitro analyses demonstrated that transmural flow downregulates and reorganizes PECAM-1 and VE-cadherin patterns and increases lymphatic permeability of the lymphatic endothelial layer (80). Importantly, transmural flow also increased the production and secretion of chemokine CCL21, essential for attraction of CCR7-expressing dendritic cells. In addition, production of adhesion molecules, ICAM-1 and E-selectin, important for lymphocyte transmigration, was also increased (80).
These data reveal a contribution of mechanical signaling to maintenance of the lymphatic vessel physiological function, such as trafficking of antigen-presenting cells [reviewed in Platt and Randolph (95)].
Clinical Relevance of Altered Lymphatic Mechanotransduction
Mutations in mechanosensitive genes in primary lymphedema
Loss-of-function FOXC2 mutations cause human hereditary disease LD, which is characterized by late onset lymphedema, venous dysfunction, and metaplasia of Meibomian glands (18, 35, 36). Given a key role of Foxc2 in regulating shear stress responses of LECs and formation of lymphatic valves (86, 92, 100), it is tempting to speculate that LD is a disease of impaired mechanotransduction, in which abnormal responses of LECs to fluid shear stress lead to defective valve development, increased vessel permeability, and impaired lymph transport (101). Furthermore, as induction of eNOS by shear stress is reduced upon loss of Foxc2 (101), it may also contribute to impaired pumping function of the collecting lymphatic vessels (13).
Of interest, mutations in GATA2, necessary for Foxc2 induction by shear stress in vitro (59), are a genetic cause of Emberger syndrome, characterized by primary lymphedema and a number of other features, including predisposition to myelodysplastic syndrome and acute myeloid leukemia (44, 60, 89).
Missense mutations impairing tyrosine kinase activity of VEGFR-3 were identified in patients with Milroy disease, characterized by congenital lymphedema of the lower limbs (56). Since VEGF-C/VEGFR-3 signaling is so central for the lymphangiogenic response (57), it may be difficult to dissociate the role of VEGFR-3 as a transducer of VEGF-C signaling versus its role as a regulator of shear stress set point in vivo (5). However, it is worth noting that Milroy disease patients display profoundly abnormal lymph transport, but only approximately 50% reduction of lymphatic capillary density in the affected feet (79). These data indicate that in addition to lymphatic hypoplasia, other factors may contribute to impairment of the lymphatic function in these patients.
Interestingly, studies in animal models demonstrated that in vivo administration of VEGF-C and VEGFR-3-specific mutant VEGF-C156S acutely increases the lymphatic pump activity of collecting vessels, which was reduced after administration of Vegfr-3 inhibitor, suggesting a direct link between Vegfr-3 activation and collecting vessel activity (17). Thus, a potential explanation of Milroy disease phenotype is that in addition to a prolymphangiogenic role in capillaries, low but detectable Vegfr-3 in collecting lymphatic vessels may be responsible for shear stress sensing and subsequent production of lymphatic pump regulators, such as NO or prostaglandins.
PIEZO-1 (FAM38A) is a large transmembrane protein recently shown to act as a mechanically activated ion channel in several cell types (6). Piezo1 −/− mice die at midgestation and have a disorganized and decreased yolk sac vasculature (98). Furthermore, endothelial-specific inactivation of Piezo1 also leads to embryonic lethality and perturbed vascular development, characterized by endothelial cell misalignment (70). In vitro Piezo1 promotes alignment in the direction of flow of blood endothelial cells subjected to laminar shear stress through the regulation of Ca2+ influx and calpain-2 activity (70, 98). Thus, Piezo1 appears to act as a bona fide sensor of shear stress in endothelial cells.
A recent exciting study demonstrated that PIEZO1 function is especially relevant to the lymphatic vasculature as homozygous and compound heterozygous mutations in PIEZO1 cause an autosomal recessive form of generalized lymphatic dysplasia, characterized by widespread lymphedema with systemic involvement of intestine, heart, and respiratory system (37). Analysis of lymph transport in patients with Piezo1 mutations by lymphoscintigraphy demonstrated rerouting of flow, suggesting failure of superficial collecting vessel function, which would be consistent with the role of Piezo1 in the shear stress response. Piezo1 is the third gene, together with CCBE1 and FAT4, found to be associated with generalized lymphatic dysplasia (1, 2); it will be therefore interesting to explore a potential mechanotransduction connection between these three genes.
Obesity and reduced transmission of mechanical stimuli to lymphatic vessels
Impaired lymphatic vascular function is associated with increased adiposity both in human lymphedema and in some animal models with defective lymphatic vascular development (46, 113). In turn, obesity is one of the factors that contribute to the development of lymphatic vascular dysfunction, including in patients with secondary lymphedema (42, 91). Although the underlying mechanisms are not well established, one of the contributing factors could be a reduced transmission of the extrinsic pumping forces, including arterial pulsations and skeletal muscle contractions, to lymphatic vessels in the presence of large amount of adipose tissue, which would result in impaired lymph flow. In addition, it is conceivable that paracrine signals from adipocytes or adipose tissue stroma may affect lymphatic vessel properties, such as permeability and vascular SMC contractility.
Tumor desmoplasia and lymphangiogenesis
Lymphangiogenesis frequently occurs in solid tumors, which release high levels of lymphangiogenic growth factors VEGF-C/D, and is usually associated with poor prognosis [reviewed in Duong et al. (33)]. Tumors are also characterized by the growth of connective tissue, or desmosplasia, and enhanced ECM production, deposition, and cross-linking leading to formation of stiff stroma, which contribute to tumor progression by potentiating cancer cell growth, survival, and migration (68). In addition, tumor IFP is usually higher compared with that of the surrounding normal tissue because of leaky tumor blood vessels (50). Importantly, the increased ECM stiffness and IFP have been shown to promote embryonic lymphangiogenesis through the increased activation of β1 integrin and Vegfr-3 signaling (94). Thus, it may be interesting in the future to study to what extent mechanical factors conspire with growth factor signaling to promote formation of the tumor-associated lymphatic vasculature.
Tissue fibrosis and impaired lymphatic vascular function
There is sufficient evidence that solid tumors are able to mount a lymphangiogenic response in the desmoplastic microenvironment. However, long-standing tissue fibrosis, such as found in chronic lymphedema patients, may also impair the lymphatic vascular function by acting as a mechanical barrier for expansion of lymphatic capillaries, which normally occurs when fluid accumulates within tissues, or by compressing the collecting vessels. Profibrotic transforming growth factor β (TGFβ) signaling is also known to directly inhibit the lymphangiogenic response (88). Indeed, it has been shown in several animal models that TGFβ-mediated fibrosis impairs lymphatic regeneration, LEC proliferation, and migration; interferes with vessel formation; and impairs lymphatic function (3, 4, 28).
Stiffening of connective tissues also impairs the lymphatic vascular function in systemic sclerosis or scleroderma, a progressive autoimmune disease, characterized by accumulation of collagen in the skin, and in visceral organs in the systemic forms (67). Analyses of the lymphatic vasculature in patients with systemic sclerosis demonstrated impaired drainage capacity of superficial lymphatic capillaries as well as progressive disappearance of lymphatic vessels at later stages of the disease, which may further contribute to the development of overt fibrosis in such patients (99).
Diabetes and regulation of chronic inflammation
Diabetes and resulting chronic hyperglycemia cause significant blood endothelium damage and dysfunction characterized by increased vascular inflammation and decrease in NO bioavailability. While cardiovascular pathology is well documented in diabetes patients [reviewed in Beckman et al. and Paneni et al. (9, 90)], relatively little is known about how this disease affects the lymphatic vascular function. Increased dermal lymphatic vessel density and endothelial cell proliferation are described in type 2 diabetes patients (43), and LECs isolated from such patients produce higher levels of chemokines and leukocyte trafficking receptors (43), indicating their exposure to a proinflammatory microenvironment. Interestingly, leptin receptor-deficient db/db mice display strongly increased collecting lymphatic vessel permeability, which could be rescued by exogenous NO administration (103).
It is tempting to speculate that low NO levels in diabetic lymphatic vessels contribute to impaired wound healing and other cardiovascular pathologies by limiting lymphatic pumping and decreasing vessel integrity, which in turn will compromise fluid and immune cell transport and will lead to maintenance of a chronic inflammatory state.
Conclusions and Further Directions
Although important insights have been gained in the past few years into the role of mechanotransduction in lymphatic vascular biology, our knowledge is far from being complete. Part of the problem lies in difficulties in modulating the lymph flow rate or flow patterns in vivo, as opposed to several approaches already developed for blood vessels [e.g., (24, 64, 96)]. Another current barrier is the paucity of quantitative information about the mechanical properties of lymphatic vessels and the surrounding tissues, as well as about the mechanical forces, experienced by lymphatic vessels in various animal models, with a notable exception of rat mesenteric vessels, for which many parameters have already been characterized (19, 32, 40, 117). Additionally, an intrinsic limitation of biomechanical studies resides in the fact that all mechanosensitive molecules are also used in other biochemical signaling cascades, making unambiguous assignment of in vivo changes to the effects of mechanical forces inherently difficult.
An important remaining question is the mechanism of shear sensing in LECs and to what extent regulators, previously identified in blood endothelial cells, such as Pecam-1, VE-cadherin, integrins, or mechanosensitive Ca2+ channels (Piezo or TRPV), are at play in the lymphatic endothelium. Among other questions that will need to be addressed in the future are the following: To what extent biomechanical forces contribute to lymphangiogenesis and lymphatic vessel maturation in different pathological conditions, especially in cancer and inflammation? How does disease or aging affect the mechanical properties of lymphatic vessels (39)? Is there a role for tissue- or disease-specific ECM rigidity or organization? How can the knowledge of lymphatic vessel biomechanics be used to provide additional opportunities for intervention in human disorders?
Footnotes
Acknowledgments
The work in authors' laboratory is supported by the Swiss National Science Foundation (PPP0033-114898, CRSII3-141811, and 31003A-156266), MEDIC, Gebert Rüf, Novartis and Swiss Bridge foundations, TheraLymph ERA-NET E-Rare Research Programme (FNS 31ER30_160674), The Commission for Technology and Innovation, Fondazione San Salvatore, Swiss Cancer League (KLS 3406-02-2014), and the People Programme (Marie Curie Actions) of the European Union's Seventh Framework Programme FP7/2007 to 2013 under REA grant agreement 317250.