
Abstract
Aims:
Electrical stimulation (EleS) can promote cardiac differentiation, but the underlying mechanism is not well known. This study investigated the effect of EleS on cardiomyocyte (CM) differentiation of human induced pluripotent stem cells (hiPSCs) and evaluated the therapeutic effects for the treatment of myocardial infarction (MI).
Results:
Cardiac differentiation of hiPSCs was induced with EleS after embryoid body formation. Spontaneously beating hiPSCs were observed as early at 2 days when treated with EleS compared with control treatment. The cardiac differentiation efficiency of hiPSCs was significantly enhanced by EleS. In addition, the functional maturation of hiPSC-CMs under EleS was confirmed by calcium indicators, intracellular Ca2+ levels, and expression of structural genes. Mechanistically, EleS mediated cardiac differentiation of hiPSCs through activation of Ca2+/PKC/ERK pathways, as revealed by RNA sequencing, quantitative polymerase chain reaction, and Western blotting. After transplantation in immunodeficient MI mice, EleS-preconditioned hiPSC-derived cells significantly improved cardiac function and attenuated expansion of infarct size. The preconditioned hiPSC-derived CMs were functionally integrated with the host heart.
Innovation:
We show EleS as an efficacious time-saving approach for CM generation. The global RNA profiling shows that EleS can accelerate cardiac differentiation of hiPSCs through activation of multiple pathways. The cardiac-mimetic electrical signals will provide a novel approach to generate functional CMs and facilitate cardiac tissue engineering for successful heart regeneration.
Conclusion:
EleS can enhance efficiency of cardiac differentiation in hiPSCs and promote CM maturation. The EleS-preconditioned CMs emerge as a promising approach for clinical application in MI treatment. Antioxid. Redox Signal. 28, 371–384.
Introduction
M
Cardiomyocyte (CM) generation from conventional approaches is laborious and time-consuming. We show electrical stimulation (EleS) as an efficacious preconditioning for CM generation. However, the pathways in human induced pluripotent stem cells (hiPSCs) activated by EleS have not been well studied. The global RNA profiling and in-depth investigations show that EleS mediated the cardiac differentiation of hiPSCs through activation of multiple pathways related to calcium signaling. Therefore, the application of cardiac-mimetic signals targeting these pathways will provide a novel approach to generate functional CMs. This knowledge will help in CM generation in cardiac tissue engineering for successful heart regeneration in a clinical setting.
Studies of heart development have demonstrated that embryonic environments (including extracellular matrix, mechanical signals, soluble factors, and electrical fields) determine the cardiac lineage commitment (1, 7). New CMs are derived from mesodermal progenitors during spontaneous differentiation (embryoid body [EB] formation) of pluripotent stem cells (24), and the physiological cues of the environment are essential to maintain the new derived CMs from hiPSCs (41).
The endogenous electric field can be detected in mouse embryonic conduction system and plays an important role in normal embryogenesis (10). However, after differentiation, the endogenous electric field might be limited within the expansion of CMs due to low yield of functional pacemaker cells (53). Therefore, the exogenous cardiac-mimetic electrical stimulation (EleS) has been applied as a conditioning treatment for the in vitro culture of CMs, particularly in myocardial tissue engineering (45, 56, 57). Additionally, the EleS approach can promote the cardiac differentiation potential of stem cells such as cardiac progenitor cells and ESCs (34, 51). We also demonstrated that the preconditioning of EleS could enhance the therapeutic efficacy of cardiac stem cells in infarcted heart (28). Thus, these studies suggest that the exogenous EleS exerts crucial effects during cardiogenesis and subsequent maturation. However, the molecular mechanisms of electrical pulses are not well known.
In this study, we sought to investigate the effect of EleS on the generation and maturation of hiPSC-derived CMs (hiPSC-CMs). The signal pathways activated by EleS were screened by next-generation RNA sequencing to reveal the relationship between physical electric pulses and biological processes. The directly involved ion channel pathways were further investigated in our cardiac differentiation model under the preconditioning of EleS, and it is our hope that investigating the molecular features of EleS should bring new insights into the process of myocardial differentiation and maturation. The knowledge of the applied EleS should then facilitate and accelerate translational studies of hiPSC-CMs in patient-specific disease modeling, drug discovery, and eventually for cell-based therapy using cardiac tissue engineering.
Results
EleS enhances spontaneous cell beating of differentiated hiPSCs
Before induction of differentiation, pluripotency of the hiPSC line was characterized. The expression of pluripotent markers such as TRA-1-60 and OCT4 was confirmed by immunostaining (Supplementary Fig. S1A, B; Supplementary Data are available online at
EleS was used to provide cardiac-mimetic pulse signals and facilitate generation of functional CMs from hiPSCs. The electrical conduction, coupling, contractile rate, and force of hiPSC-CMs are distinct from adult CMs depending on the stage of differentiation (66). The hiPSC-CMs were exposed to different frequencies (from 0 to 5 Hz), but no tonic contraction was observed (Supplementary Fig. S2C), indicating the electrophysiological plasticity of hiPSC-CMs. In parallel to the previous study (45) using a high frequency (∼6 Hz) of EleS for engineering cardiac tissues, we selected 5 Hz as a coupling condition for the generation of hiPSC-CMs. Preconditioning of EleS at a high frequency also can enhance the electrophysiological and contractile synchronization between hiPSC-CMs and mouse heart in the present study.
The initial time of spontaneous beating was recorded daily to determine the effects of EleS (1 or 1.5 V/cm) on differentiated hiPSCs. Attached EBs without EleS started contracting at 7.0 ± 0.63 days (Fig. 1A). In contrast, the spontaneous beating of attached EBs with EleS can be observed as early at 2.1 ± 0.40 days (1 V/cm) or 3.2 ± 0.40 days (1.5 V/cm) (Fig. 1A and Supplementary Movie S1). The percentage of beating EBs with EleS (1 or 1.5 V/cm) was significantly increased compared with non-EleS treatment after attachment in culture for up to 25 days (Fig. 1B). Furthermore, the spontaneous contraction frequency of hiPSC-derived EBs was increased from day 2 to 30 in a time-dependent manner, which was higher under EleS compared with non-EleS control (Fig. 1C). There was no statistically significant difference in the contraction frequency of attached EBs under 1 V/cm or 1.5 V/cm EleS. We selected 1 V/cm EleS for the following experiments not only based on these results but also because reactive oxygen species can be moderately generated (49, 51).

EleS upregulates the expression of cardiogenic genes in differentiated hiPSCs
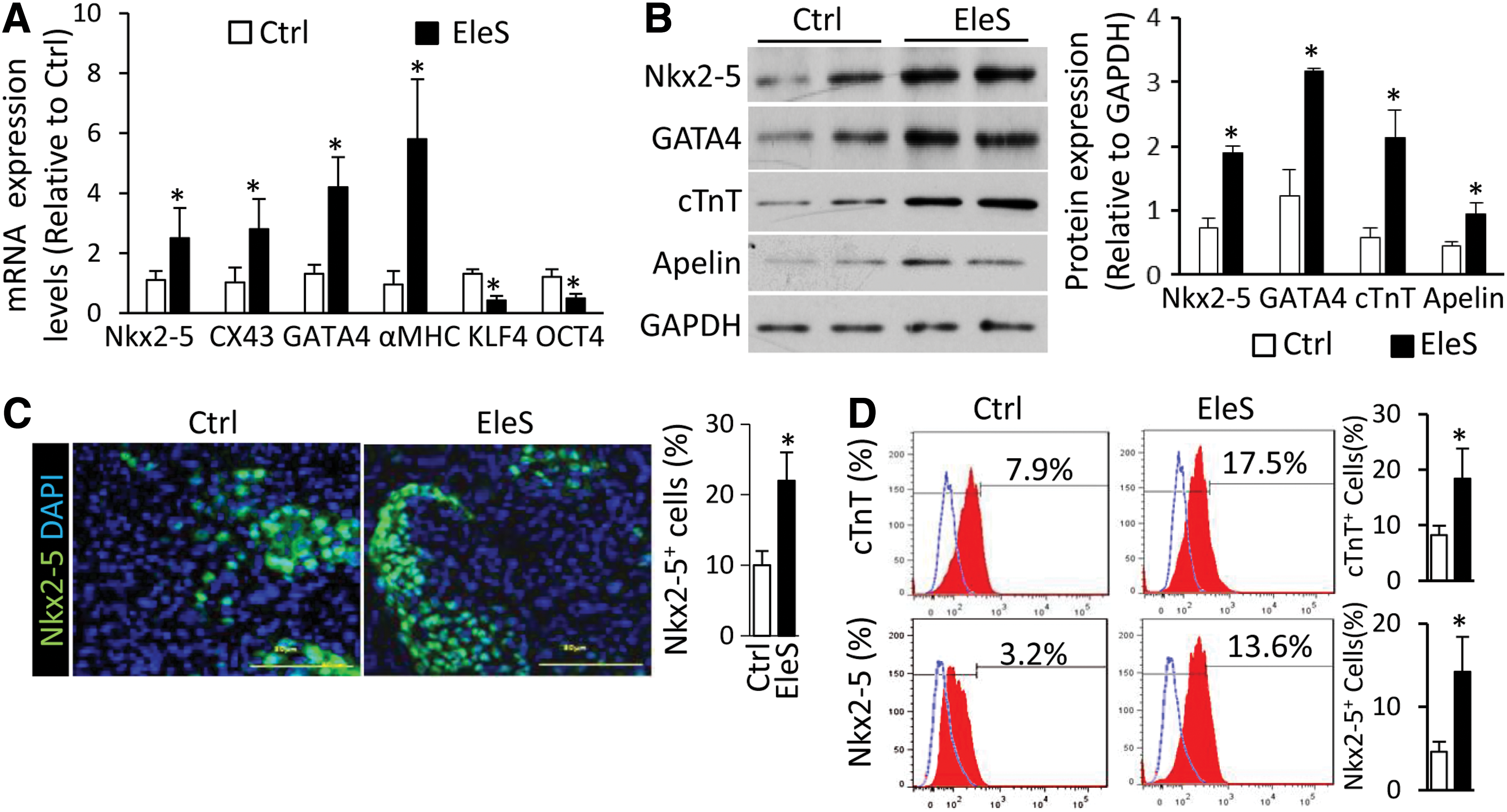
Cardiac genes were barely detected in undifferentiated hiPSCs, but were activated in spontaneously differentiated cells. Focus was placed on the relative expression levels of various genes in differentiated hiPSCs in the absence or presence of EleS. Compared with the control group, the cardiac progenitor genes (Nkx2-5 and GATA4) and structural genes (alpha myosin heavy chain [αMHC] and connexin 43 [CX43]) were upregulated by EleS in hiPSCs after 1-week induction, while the relative expression of pluripotent markers (KLF4 and OCT4) was decreased as examined by quantitative polymerase chain reaction (qPCR) (Fig. 2A). The protein levels of these cardiogenic markers (such as Nkx2-5, GATA4, and cardiac troponin T [cTnT]) and peptides [such as Apelin (59)] were also significantly increased in differentiated hiPSCs under EleS compared with the control treatment (Fig. 2B and Supplementary Fig. S3). Immunostaining also showed that Nkx2-5 located in nuclei of differentiated hiPSCs was upregulated by EleS (Fig. 2C). CM differentiation efficiency of hiPSCs was enhanced by EleS preconditioning as shown by FACS of CM markers, cTnT and Nkx2-5 (Fig. 2D). The percentage of Nkx2-5+ cells in immunostaining was different from the results of FACS, indicating the variance of sensitivity for the different staining and analytic techniques.
EleS facilitates functional maturation of hiPSC-CMs
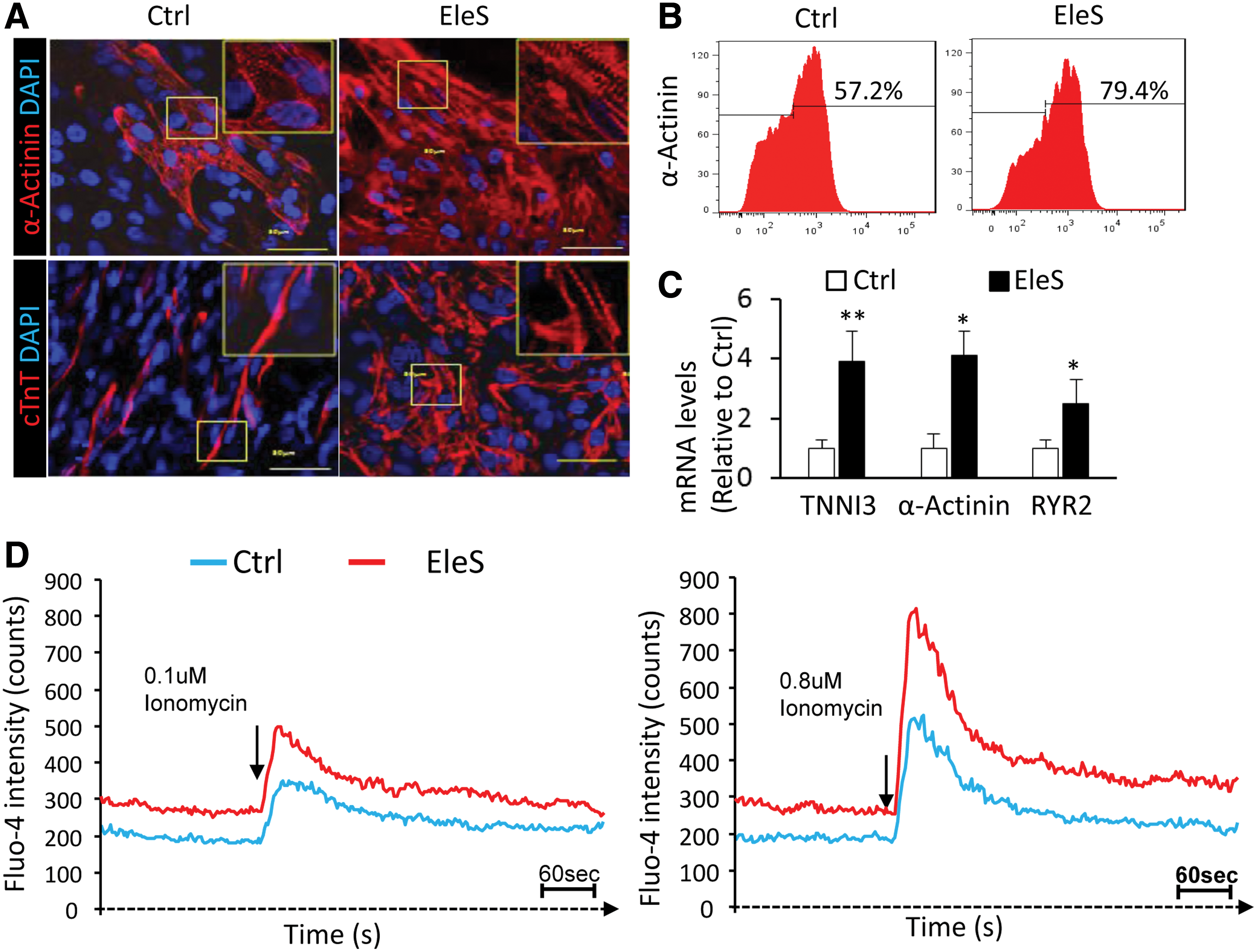
After EleS preconditioning for 2 weeks, immunostaining of cardiac structural proteins, including α-actinin and cTnT, shows that hiPSC-CMs displayed the characteristic striations indicative of sarcomere formation, while the immature structure was observed in the control hiPSC-CMs without EleS (Fig. 3A). Almost 80% of hiPSC derivatives under EleS preconditioning were α-actinin-positive CMs as identified by FACS, while the CM population spontaneously generated in the control group was less than 60% (Fig. 3B). The mRNA levels of cardiac genes (such as TNNI3, α-actinin, and ryanodine receptor 2 [RYR2]) related to functional maturation were also upregulated by EleS (Fig. 3C). Moreover, the spontaneous cellular Ca2+ flux of hiPSC-CMs with EleS can be detected by the fluorescence of cTnT-GCaMP calcium indicator using live cell imaging (Supplementary Movie S2). As shown by Fluo-4 dye, EleS preconditioning for 2 weeks increased the intracellular Ca2+ levels of cultured hiPSC-CMs compared with control treatment (the baselines in Fig. 3D). In response to different dosages of ionomycin (a selective Ca2+ ionophore), intracellular Ca2+ levels of hiPSC-CMs were significantly increased (Fig. 3D).
Global gene expression profiles of hiPSCs during differentiation
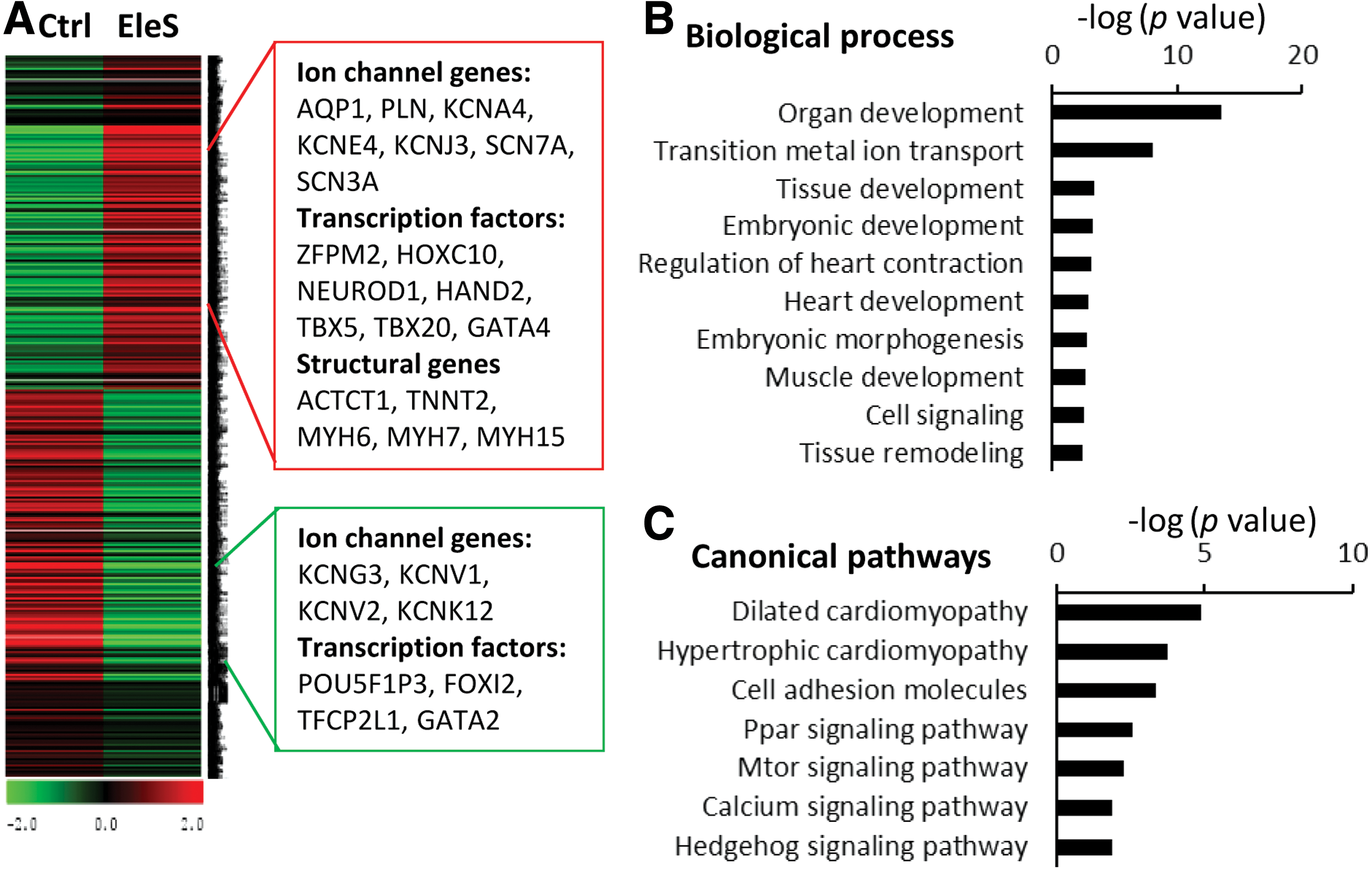
The global RNA profiles of hiPSCs were analyzed by RNA sequencing (RNA-seq) after 1 week of induction with or without EleS to identify the signal pathways activated by EleS. To increase the analytic resolution of RNA-seq, the cells attached for 7 days were used to assess early gene expression patterns changed by EleS. Selecting twofold change and p-value <0.05 as a cutoff threshold for data analysis, 558 genes were upregulated by EleS, while 513 genes were downregulated (Fig. 4A). For instance, the cardiac transcription factors (such as HAND2 and TBX5) and construction genes (such as ACTCT1 and TNNT2 [cTnT]) were upregulated by EleS (Fig. 4A). The upregulated genes activated by EleS were investigated using Gene Set Enrichment Analysis (GSEA) in Molecular Signatures Database. Based on the biological process ontology, EleS enhanced the developmental processes, including regulation of heart contraction, and heart development (Fig. 4B). Moreover, the KEGG pathway analysis shows that the canonical pathways (such as the calcium-signaling pathway) were involved in EleS-induced differentiation of hiPSCs (Fig. 4C). According to molecular function ontology, several genes with ion binding activity were upregulated by EleS (Supplementary Fig. S4). Thus, we further explore the specific signaling pathways related to electrophysiology.
EleS activates the calcium signaling pathways during hiPSC differentiation
An increase of extracellular Ca2+ influx has been reported in the environment with electric or magnetic fields (6, 49, 65), which is consistent with our hiPSC differentiation models. However, it is still unclear how the commitment of hiPSCs into the cardiac lineage is influenced by calcium signaling under EleS. Ca2+-binding proteins (calmodulin [CAML] 1 and 3) were upregulated in hiPSCs by EleS induction for 1 week (Fig. 5A). In hiPSCs induced with EleS, the expression of cardiac genes (GATA4, TBX5, CX43, and TNNI3) was inhibited by multiple calcium signaling inhibitors, including verapamil (Ca2+ ion channel blocker), W-7 (CAML antagonist), and TMB-8 (intracellular Ca2+antagonist), although αMHC was not significantly impacted by verapamil (Fig. 5B). Furthermore, Western blotting showed that the expression of cardiac transcription factors (GATA4 and MEF2) in hiPSCs was enhanced by EleS compared with non-EleS control, whereas the calcium signaling inhibitors reversed the effects of EleS (Fig. 5C and Supplementary Fig. S3). We found that histone deacetylase 1 (HDAC1) was downregulated by EleS compared with non-EleS control, but this effect was reversed by the calcium signaling inhibitors (Fig. 5C and Supplementary Fig. S3). The levels of cardiac contraction proteins (cTnT and αMHC), gap junction proteins (CX43), and cardiogenic peptides (Apelin) were also significantly increased in differentiated hiPSCs under EleS conditioning compared with the non-EleS control, but these gene activations by EleS were consistently abolished by calcium signaling inhibitors (Fig. 5C and Supplementary Fig. S3). EleS thus enhanced cardiac gene expression of hiPSCs through the activation of intracellular Ca2+ signals and the downstream genes (Fig. 5D).
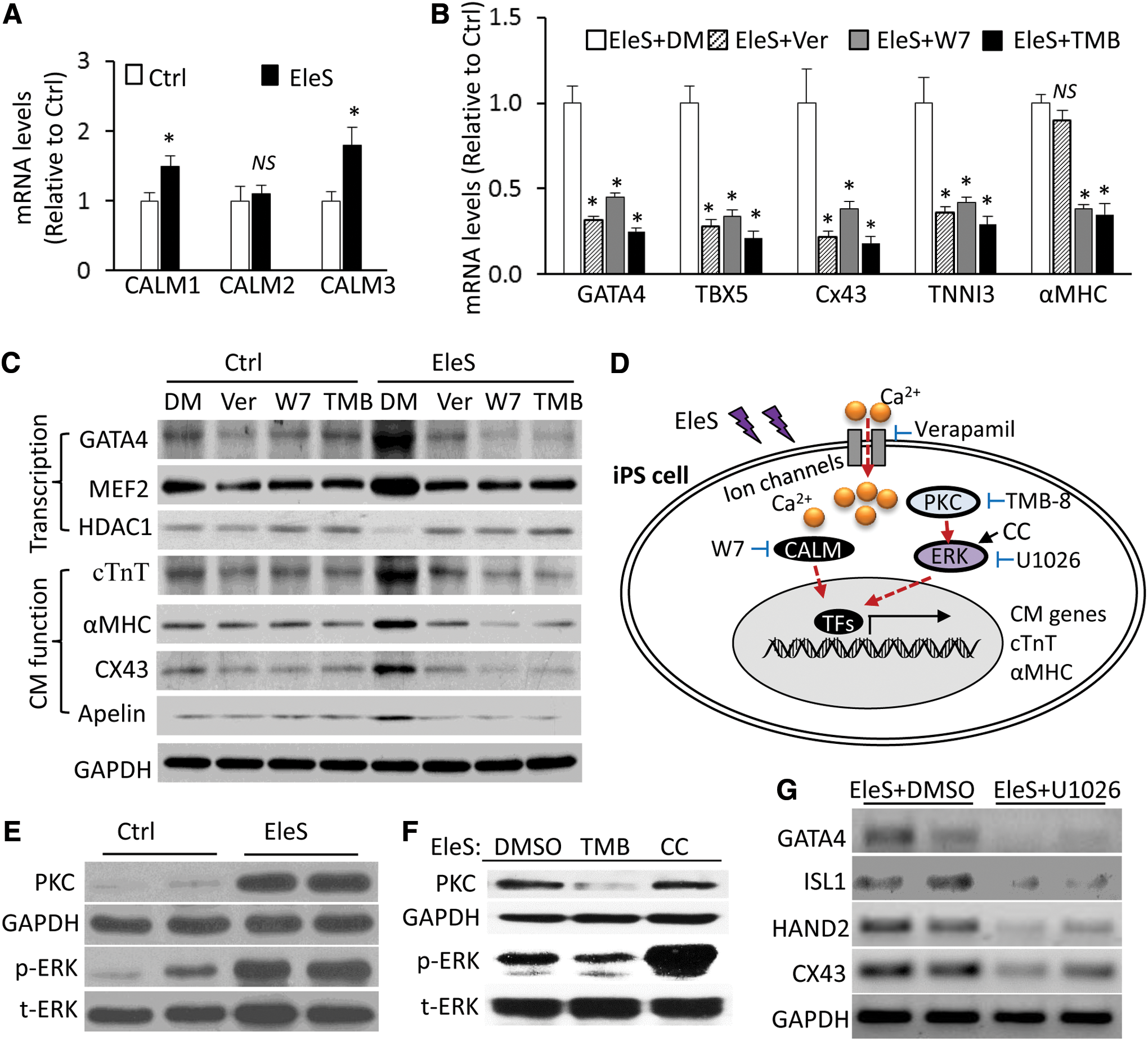
Additionally, we found that PKC expression and ERK phosphorylation were enhanced by EleS preconditioning in hiPSCs (Fig. 5E and Supplementary Fig. S3). Under EleS, the treatment of TMB-8 [inhibitor of both intracellular Ca2+and PKC (3)] can decrease the PKC expression and ERK phosphorylation compared with dimethyl sulfoxide (DMSO) treatment (Fig. 5F and Supplementary Fig. S3). The treatment of chelerythrine chloride [CC, activator of MAPK pathways (67)] can increase ERK phosphorylation, but not impact on the PKC expression compared with DMSO treatment (Fig. 5F and Supplementary Fig. S3). The expression of cardiac genes of hiPSCs under EleS was eliminated by ERK inhibitor U1026 (Fig. 5G). These data suggest that intracellular Ca2+ and PKC were required for ERK phosphorylation during CM differentiation and that activation of ERK cannot reversely affect its upstream PKC expression (Fig. 5D).
EleS-preconditioned hiPSC-CMs improve cardiac functions of ischemic heart
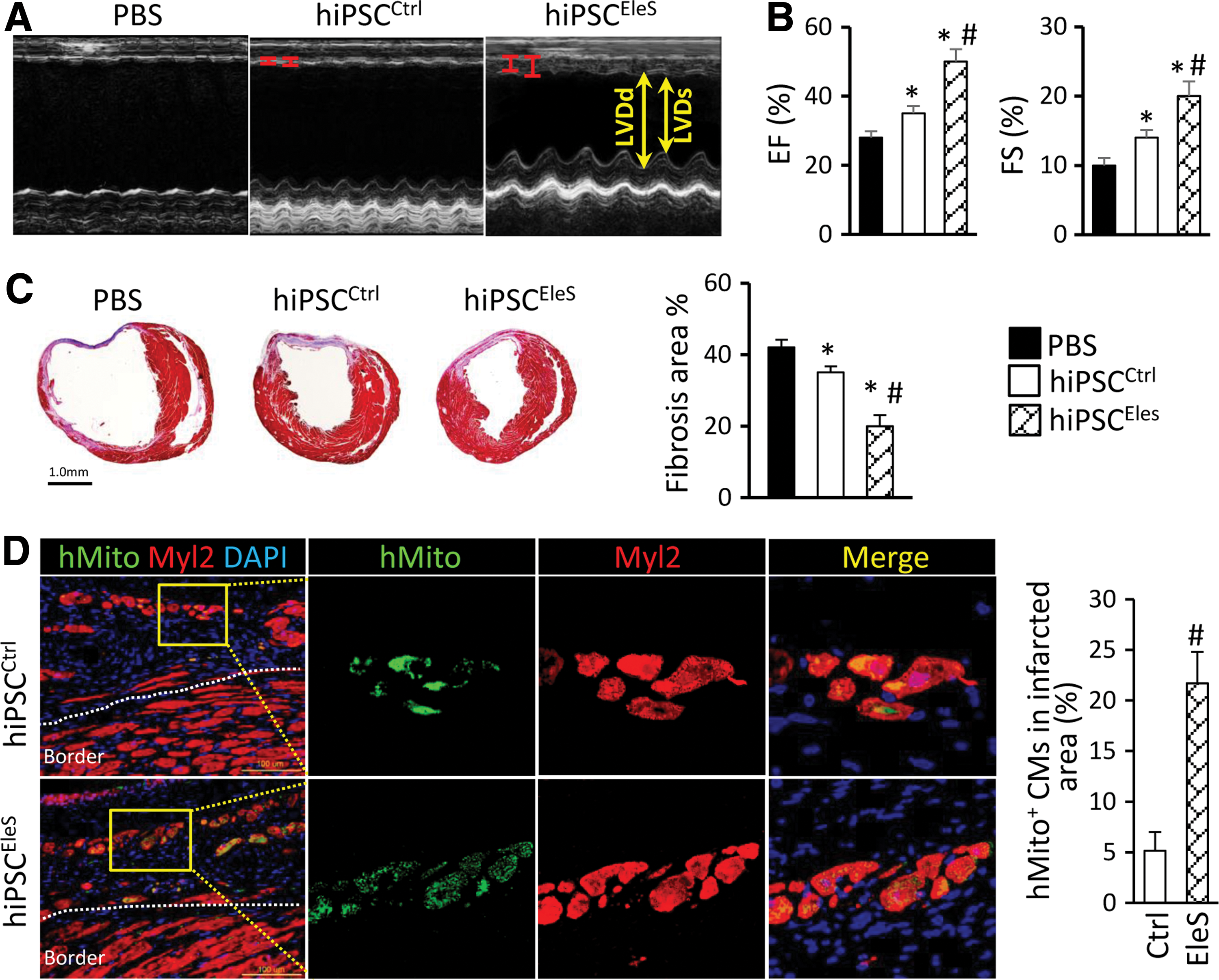
The percentage of hiPSC-CMs in the mixed derivatives was high in the induction system (∼60% in control vs. ∼80% in EleS treatment), as shown in Figure 3B. Subsequently, the control-treated (hiPSCCtrl) or EleS-preconditioned hiPSC derivatives (hiPSCEleS) were injected into the infarcted heart of severe combined immunodeficiency (SCID) mice to evaluate the therapeutic effects of hiPSC-derived CMs. In these experiments, cardiac functions were assessed by echocardiograph after cell delivery. At week 4 after cell transplantation, cardiac functions of MI mice were improved in the hiPSCCtrl group, as evidenced by decreasing left ventricular end-diastolic diameters (LVDd) and left ventricular end-systolic diameters (LVDs) and increasing ejection fraction (EF) and fractional shortening (FS) when compared with the phosphate-buffered saline (PBS) treatment (Fig. 6A, B). Remarkably, the cardiac protective effect of hiPSCs was significantly enhanced by EleS preconditioning compared with the non-EleS-treated cells (Fig. 6A, B). Microscopic analysis by Masson's trichrome staining showed that the average infarct area of the hiPSCEleS group was smaller than that of the hiPSCCtrl or PBS group (Fig. 6C). Finally, mitochondrial protein (hMito, identified using the specific antibodies) was used to determine whether the engrafted hiPSC-CMs contributed to functional restoration after MI. While the endogenous CMs in the injury border zone were not stained by the hMito antibody in MI mice (Fig. 6D), the number of hMito+ CMs in the hiPSCEleS group was fourfold higher than that of the hiPSCCtrl group in infarcted heart (Fig. 6D). Teratoma formation was not observed in our experiments after transplantation of the hiPSC derivatives for over 1 month because the pluripotent genes in hiPSCs were silenced during the differentiation process (Supplementary Fig. S2B).
Discussion
The heart is an electromechanical pump with CMs that are constantly exposed to regular electrical stimuli during early development and into adulthood (10). External EleS has been reported to mimic the electric properties of native heart tissue and facilitate CM differentiation (49). hiPSCs have been emerging as a promising substitute of ESCs for cardiovascular cell regeneration, and current iPSC technology using adult somatic cells avoids ethical legal issues and is technically feasible (30, 54). However, few reports focus on the role of EleS in CM differentiation of hiPSCs and the underlying mechanisms. It is also unknown whether the EleS-preconditioned hiPSC-CMs are superior to the standard hiPSC-CMs used in cell therapy for MI treatment. Our results indicated that (i) EleS promoted the CM differentiation potential and maturation derived from hiPSCs; (ii) EleS induced hiPSC differentiation through Ca2+/PKC/ERK pathways; and (iii) EleS-preconditioned hiPSC-CMs significantly improved the heart functions of MI mice after cell transplantation.
Currently available CM differentiation techniques were inefficient or not readily transferable across stem cell lines (41). Most in vitro differentiation protocols have been developed based on activating the cardiomyogenesis-induced signaling pathways to enhance the generation of CMs from hiPSCs. A range of different growth factors (such as BMP4, activin A, or bFGF) or small molecules (such as Wnt or GSK inhibitors) have been tested to improve the process of cardiomyogenesis in ESCs or iPSCs (9, 31, 70). In addition to the stimulation of soluble factors, the bioelectrical pulses may determine the emergence of spatial patterns and aid in tissue morphogenesis during embryo development (57). To develop electric field as an efficacious stimulus in CMs generated from hiPSCs, the biomimetic system without additional growth factors was designed to deliver electric signals mimicking the bioelectrical environment of native heart. Intriguingly, a higher percentage of the spontaneously beating hiPSC-derived EBs can be observed as early at 2 days after EleS compared with those without EleS. It is known that spontaneous beating represents a phenotype of functional CMs. Furthermore, the hiPSC-CMs retain the electrophysiological plasticity that is different from adult human CMs. The spontaneous contraction of hiPSC-derived EBs (∼60/min) after EleS displayed a similar frequency to the adult human heart. The cardiogenic factors such as GATA4 and Nkx2-5 in differentiated hiPSCs were upregulated by EleS as well as the structural markers (cTnT and αMHC). Thus, these findings suggested that EleS was a potent inducer for cardiac differentiation of hiPSCs.
The generation and maintenance of functional CMs are of critical importance for the clinical applications of hiPSCs (66). In the present study, the applied EleS facilitated the morphological formation of CMs as evidenced by the formation of sarcomeric striations. The sarcomere is known as the fundamental unit of CM contractility. Under EleS, the sarcomeric structure of hiPSC-CMs became more organized, facilitating force generation. The yield of sarcomeric α-actinin-positive CMs from hiPSCs with EleS was up to 80%. Furthermore, calcium handling is one of the most widely characterized properties of functional CMs. The Ca2+-handling proteins, including RYR2, were upregulated in hiPSC-CMs generated with EleS. As monitored by fluorescent calcium indicator, the synchronized cardiac Ca2+ transients were present in the hiPSC-CMs with EleS, which were barely observed in the hiPSCs without EleS (data not shown). We also found that the basal intracellular Ca2+ levels were increased in hiPSC-CMs induced with EleS. The external Ca2+ influx is an important initial trigger for excitation–contraction coupling in adult CMs or hiPSC-CMs (22). In addition, the release of Ca2+ from intracellular stores of differentiated hiPSCs was demonstrated by the response to ionomycin in the present study. Therefore, EleS preconditioning was beneficial for the maintenance and functional maturation of hiPSC-CMs, including contractility and calcium handling.
It is the first time the global RNA profiles of hiPSCs under EleS have been revealed to understand the signal pathways activated by EleS. The next-generation RNA-seq has become a key milestone in the field of systems biotechnology. It not only reflects the quantitative gene expression levels but also identifies the transcript isoforms (64). This technique was applied to generate a high-resolution transcriptome map comparing the gene expression of hiPSCs with or without EleS. Intriguingly, multiple ion channel genes, transcription factors, and structural genes were significantly upregulated under EleS, but the underlying mechanisms are not fully known. Importantly, RNA-seq with gene set analysis tools such as GSEA can evaluate expression patterns of gene groups rather than those of individual genes (21, 42). The analyses of RNA-seq data confirmed that the expression changes for genes were involved in CM development, cell adhesion/cytoskeletal components, and calcium signaling pathways during differentiation of hiPSCs with EleS. Intriguingly, the transcriptome of neonatal CMs revealed that the pulse-specific gene expression activated by EleS was independent from evoked contractions (37). These significant pathways activated by EleS are required for further investigation to understand the molecular mechanisms of CM generation from iPSCs or ESCs. This study focuses on calcium signaling based on the gene and pathway profiling.
Although EleS was reported as an approach for regulating cell differentiation or migration (15, 51), the potential signaling pathway in hiPSCs activated by EleS has not been well studied. In the electrical field, the first responsive molecules are the migrated ions, although the zero direct current average voltage is produced by the stimulator device to avoid electrolysis (34). It is a general electrophysiological phenomenon that the cell membranes are depolarized at the cathode-facing side with hyperpolarization at the anode-facing side (15). Redistribution of cell surface receptors was also reported under alternating current electric fields (13). Therefore, the voltage-dependent ion channels are open in response to EleS and subsequent movement of ions. The extracellular Ca2+ influx increased by electric fields has been demonstrated in cardiac progenitors or ESCs (6, 49). In the present study, the addition of verapamil calcium channel blocker can significantly decrease the expression of cardiac genes such as GATA4 and TBX5 in hiPSCs, indicating that the opening of calcium channels was an early response to EleS. Furthermore, the increasing Ca2+ in cytoplasm can activate Ca2+-binding proteins such as CAML, as shown in the present study. The treatment of CAML antagonist or intracellular Ca2+ antagonist can dramatically block the expression of cardiac genes, including cTnT and αMHC, in differentiated hiPSCs under EleS. These findings documented that the calcium signaling pathway plays an essential role in CM generation from hiPSCs under EleS preconditioning.
The crosstalk between Ca2+, PKC, and ERK signals was demonstrated in B cells (19, 44). We also found that calcium could potentially activate ERK phosphorylation through upregulation of PKC, suggesting that these pathway networks participate in the differentiation process. Interestingly, HDAC1 acted as a downstream gene and is downregulated in response to EleS. Studies demonstrated that downregulation of HDAC1 could enhance chromatin acetylation in cardiac genes, thereby promoting cardiac differentiation (35, 36, 38). Therefore, the epigenetic pathways, including histone modification, are involved in EleS environment and require further investigation. In addition to cardiac transcription factor and structural genes, Apelin, a secreted peptide that was reported to enhance cardiac differentiation of mouse and human ESCs (59), was upregulated by EleS preconditioning. Thus, our study demonstrated that EleS modulated the intracellular Ca2+ levels, influenced enzyme function, and then induced multiple signal pathway networks, resulting in the cardiac lineage commitment of hiPSCs.
The therapeutic effects of hiPSC-CMs with or without EleS were evaluated in infarcted heart of immunodeficient SCID mice. A promising strategy for regenerating damaged myocardium is to exploit cardiac tissue engineering using iPSC-CMs with biomaterials for in vitro construction of 3D structures (18, 33). In this study, long-term incubation enhanced the spontaneous differentiation of hiPSCs, while preconditioning of EleS facilitated CM generation. EleS contributed to the maintenance and functional maturation of hiPSC-CMs, consistent with other studies (11, 48, 56). The hiPSC-CMs in the control (∼60%) or EleS (∼80%) group were the majority of injected cell populations. The in vivo studies showed that the heart function of MI mice was significantly improved by implanted hiPSC-CMs with EleS compared with the non-EleS treatment. Moreover, the substitution of scar tissue with the implanted cells was identified in the infarcted area using human species marker such as hMito. The cell engraftment of hiPSC-CMs with EleS in infarcted heart was much higher compared with the control hiPSC-CMs, although the injected cell number was slightly different.
The beneficial effects of engrafted hiPSC-CMs resulted from the preconditioning of EleS. Speculation based on the in vitro differentiation and maturation studies would be that the hiPSCs with EleS could generate more functional CMs than the control hiPSCs did. The hiPSC-CMs pretreated with EleS mimicking heart environment could more readily incorporate into the mouse myocardium, compared with the control hiPSC-CMs with incompatible electromechanical properties. Therefore, the functional hiPSC-CMs can replenish the cells lost in infarcted area, and then restoration of heart function resulted from cell engraftment or functional integration. In addition, cell engraftment can minimize the adverse scar growth, ultimately reducing adverse remodeling (23, 25, 43). Although other effects such as angiogenesis or antiapoptosis (by paracrine factors) cannot be excluded, our previous studies have demonstrated cell engraftment of implanted CMs as the primary contributor to the cardiac functional restoration by using a specific cell suicide approach (61). Thus, EleS appears as an important preconditioning approach for CM generation from hiPSCs in the design of cell-based therapies for infarcted heart.
However, the risk of arrhythmia is a major challenge hampering the clinical applications of iPSC-CMs in repair of failing heart. When electromechanical features of implanted cells are not synchronized with the host heart, arrhythmia such as ventricular tachycardia can be induced via reentrant circuits or graft automaticity (12, 27). Recent preclinical studies have indicated that transient, nonlethal ventricular tachycardia was increased after implantation of iPSC- or ESC-CMs, but this effect was dramatically decreased after 2 weeks and the engrafted CMs showed contractile synchronization with the host heart (14, 52). Therefore, techniques to enhance the functional integration of hiPSC-CMs through control of contractility rates are necessary to minimize the risk of arrhythmia after transplantation. In agreement with a recent report (16), we found that the frequency of autonomously beating hiPSC-CMs was increased in correlation with the increasing frequencies of EleS pacing. It was demonstrated that the acquired beating frequency of hiPSC-CMs could be maintained and continuously transferred to the surrounding cells even after EleS was discontinued (16). The electrophysiological plasticity of hiPSC-CMs during differentiation or maturation allows generation of CMs with different beating frequencies induced by EleS for the desired target region. Thus, optimization of EleS frequency will be required in future preclinical studies of large animals to conform to the beating frequency of hiPSC-CMs for the host heart.
Additionally, it is still a challenge to obtain pure CM populations using currently available CM enrichment approaches such as FACS or metabolic selections (50, 58), but the final purity of hiPSC-CMs can reach 90% through these purification approaches. However, most CMs would be lost or damaged during the sorting processing, resulting in low CM yield (17). Contaminating cells also cannot be eliminated through these metabolic selections because other cells with high mitochondrial density such as skeletal myocytes (47) may still survive during the selection process. Discouragingly, the purified CMs did not attach well after plating on gelatin-coated plates, resulting in additional cellular losses (17). Conversely, the nonmyocytes such as fibroblasts or endothelial cells derived from hiPSCs supported the cell survival of hiPSC-CMs through cell–cell interactions or paracrine effects. Therefore, the hiPSC derivatives were selected in this study without the purifying process to maintain a ready growth environment of hiPSC-CMs. Mixed hiPSC derivatives were assessed to determine their role in promoting cardiac functional restoration in MI animals.
Additionally, mixed hiPSC derivatives were used instead of purified CMs for the RNA-seq study, demonstrating that the different expression patterns were associated with the numbers of hiPSC-CMs. However, the expression patterns were not a direct result of the increased number of CMs. Instead, the gene expression of hiPSC-CMs was impacted more significantly by the growth environment. It should be noted that the starting cells (hiPSCs) of the two groups were the same and the percentage or number of generated CMs was dependent on the condition of stimulation. The differentiation of non-CMs was also influenced or changed by the condition of EleS. Therefore, the molecular mechanisms of EleS were elucidated by revealing the global gene expression profiles of the entire hiPSC derivatives. In future studies, cell components of cardiac tissue engineering will be investigated by isolating the desirable cells for CM growth combined with the optimally defined medium containing the cardiogenic cocktails (29) with EleS preconditioning to improve CM differentiation and maturation.
In summary, EleS was shown to directly enhance CM differentiation of hiPSCs and continuous EleS preconditioning can facilitate the functional maturation of derived CMs. Mechanistically, EleS mediated the expression of the key cardiac transcription factors and structural genes through activation of calcium signaling pathways as well as cooperation with PKC/ERK pathway networks. The transplantation of hiPSC-CMs pretreated with EleS can effectively incorporate and repair the infarcted heart of MI mice.
Materials and Methods
All research protocols conformed to the Guidelines for the Care and Use of Laboratory Animals published by the National Institutes of Health (National Academies Press, eighth edition, 2011). All animal use protocols and methods of euthanasia used in this study were approved by the University of Cincinnati Animal Care and Use Committee.
hiPSC culture and EB formation
The episomal hiPSC line was purchased from Life Technologies (Invitrogen) and adapted to feeder-free culture conditions. The cell maintenance and differentiation approaches were performed according to the manufacturer's instructions. The differentiation protocol of hiPSCs through EB formation was modified according to our previous report (61). Briefly, hiPSCs were maintained in Essential 8™ culture medium on culture dishes coated with vitronectin (0.5 μg/cm2) at 37°C in a humid atmosphere of 5% CO2 and expanded for 3–4 days. Similar to ESCs (17, 26), spontaneous cardiac differentiation of hiPSCs was achieved by using fetal bovine serum (FBS)-containing medium. The iPSCs that grew out from the culture colonies were digested with Dispase solution (Gibco) at 37°C for 3–5 min and placed in ultralow attachment plates (Corning) in suspension culture for 7 days. Differentiation medium consisting of 0.1 mM nonessential amino acids, 1 mM L-glutamine, 0.1 mM β-mercaptoethanol, and 20% FBS in high-glucose Dulbecco's Modified Eagle Medium (DMEM) (Gibco) was used to initiate EB formation.
Conditioning of EleS
After 7 days of EB formation, the hiPSC colonies were seeded on vitronectin-coated dishes with CM growth medium containing 20% M199 (Gibco), 0.1 mM nonessential amino acids, 1 mM L-glutamine, and 20% FBS in low-glucose DMEM (Gibco). After cell attachment for 12 h, EleS was added using the C-Pace electric stimulator (IonOptix), which contains two electrodes emitting bipolar stimuli without electrolysis. As previously described (28), the electric field (1 or 1.5 V/cm) was established to mimic endogenous electrical signals in native mouse heart with a biphasic square pulse (5 ms) at 5 Hz frequency for 1–30 days. In addition, the attached EBs without EleS were cultured as baseline controls. The timing of the formation of the beating cell clusters/clones was recorded daily, and effects of the electric field were assessed as an independent factor. Other factors, such as BMP4 and activin A [with potential effects on cardiac differentiation (41)], were not used in our study to isolate the role of EleS and allow for study of the underlying mechanisms.
RNA extraction and real-time PCR
Total RNA was isolated from various treatment groups of cells using the RNeasy Mini kit (Qiagen), and cDNA was prepared using the Omniscript-RT kit (Qiagen). Each PCR was performed with specific primers, and the QuantiTect SYBR green PCR kit (Qiagen) was used with BIO-RAD-iQ5 optical module as previously described (61). The mRNA level was standardized to internal control (GAPDH) and expressed as fold changes. These relative levels of cellular gene or mRNA expression with EleS were normalized by the gene expression levels in control cells (without EleS). All of the PCR primer sequences in this study are listed in Supplementary Table S1.
RNA-seq
Total RNA was isolated from hiPSCs using the RNeasy Mini kit (Qiagen). RNA-seq and data analysis were performed by the Genomics, Epigenomics, and Sequencing Core Center at University of Cincinnati. The sample processing, sequencing, and data analysis were described in previous publications (68, 69). Three replicates were performed of the RNA-seq experiment, and the p-value confirmed the confidence levels of gene expression.
Western blot
The cells from various treatment groups were lysed with ice-cold cell lysis buffer plus protease inhibitor (Sigma). The protein samples were mixed and resolved in 6 × SDS/PAGE sample buffer and boiled for 10 min before loading on proper polyacrylamide gels (Precast Gels; ISC Bioexpress). As previously described (32), the electrophoresed proteins were transferred to nitrocellulose membranes (Bio-Rad). The membranes were incubated for 60 min with 5% dry milk and Tris-buffered saline to block nonspecific binding sites. The specific antibodies used for detection of antigens of interest are listed in Supplementary Table S2.
Immunocytochemistry
Immunocytochemical analysis was performed as previously described (63). Briefly, the hiPSCs were fixed with 4% paraformaldehyde for 15 min at 4°C and permeabilized in 1% Triton X-100 in PBS, followed by two washes with PBS. After treatment with serum blocking buffer for 10 min, samples were incubated with specific primary antibodies, followed by their related secondary antibodies. Immunostaining was observed under the fluorescence microscope (Olympus). Antibodies used are listed in Supplementary Table S2.
Flow cytometry
As previously described (61), single-cell suspensions of differentiated iPSCs were trypsinized 20 days after differentiation. About 106 cells were fixed on ice with 4% paraformaldehyde for 30 min and permeabilized in 0.1% saponin for 30 min, followed by 1% FBS on ice for 30 min. Thereafter, cells were stained with antibodies against CM genes, followed by a secondary antibody, for 1 h at room temperature. Analysis was performed using the FACS Aria instrument (BD Biosciences). Cells with an adequate size and granularity were accounted for analysis.
Calcium dye loading and fluorescence imaging
Intracellular Ca2+ was measured in cell monolayers loaded with the Ca2+-sensitive probe Fluo-4-AM (Invitrogen). After loading with 10 μM of Fluo-4-AM in serum-free medium at 37°C for 60 min, cells seeded on the coverslip were washed and the fluorescence signal was detected by a confocal laser scanning microscope (Olympus), as previously described (20). The intracellular Ca2+ levels were analyzed by FACS in response to different dosages of ionomycin (Sigma). Additionally, spontaneous cellular Ca2+ flux was detected using cTnT-GCaMP calcium indicator under the control of cTnT promoter (2) (Addgene ID: 46027), which was transiently transfected in hiPSC-CMs and the autofluorescence was recorded by the confocal microscope.
Acute MI model and cell transplantation
To minimize immune rejection, 10–12-week-old immune deficiency (SCID) mice (Stock No: 001303) were purchased from The Jackson Laboratory. Ligation of left anterior descending (LAD) coronary was performed as described in our previous publication (62). Briefly, mice were anesthetized by intraperitoneal administration of 0.1% ketamine and 0.02% xylene. Heart was exposed by left-sided minimal thoracotomy and LAD coronary artery was ligated using 6-0 silk. The cells were exposed to EleS (or 0 V/cm as control) for 3 weeks before implantation into the infarcted heart. The intramyocardial delivery of the hiPSC derivatives (2 × 105 cells) containing derived CMs with or without EleS was performed as described previously (60). Approximately 30 μl of cell suspension was injected into the border zone of the infarcted heart at three different areas. Four weeks later, the hearts were excised and analyzed by immunocytochemistry staining.
Cardiac function assessment by echocardiography
Transthoracic echocardiography (iE33 Ultrasound System; Phillips) was performed with a 15-MHz probe. Echocardiographic data were analyzed in an unbiased manner that was blinded to the experimental groups. Animals were placed supine on an electrical heating pad at 37°C under light isoflurane anesthesia (usual maintenance level 1.5% isoflurane/98.5% oxygen). Hearts were imaged in 2D long-axis view at the level of the greatest LV diameter with animals under light general anesthesia. This view was used to position the M-mode cursor perpendicular to the LV anterior and posterior walls. LV end-diastolic and end-systolic diameters were measured from M-mode recordings. LV EF was calculated as EF (%) = [LVDd3 − LVDs3/(LVDd)3] × 100. LV minor axis FS was also determined as [(LVDd − LVDs)/LVDd] × 100. All measurements were performed according to the American Society for Echocardiography leading-edge technique standards and averaged over three consecutive cardiac cycles.
Immunohistochemistry in paraffin-embedded tissues
Heart tissue sections were harvested, fixed in 10% formalin, embedded in paraffin, and sectioned at 8 μm thickness. After deparaffinization and rehydration, the sections were treated with citric acid/microwave antigen retrieval for 10 min. After treatment with serum blocking buffer for 20 min, slides were then incubated with antibodies (such as myosin light chain 2 [Myl2] and hMito) for 2 h at room temperature. After removing the primary antibody, fluorescein-labeled secondary antibody was added and incubated for 1 h at room temperature. Nuclei were stained with DAPI. Immunostaining was then observed microscopically.
Statistical analysis
All values are expressed as mean ± SEM. Comparisons between two mean values were evaluated by an unpaired Student's two-tailed t-test and between three or more groups were evaluated by one-way ANOVA, followed by Bonferroni post hoc analysis. Statistical significance was accepted at p-value <0.05.
Footnotes
Acknowledgments
This work was supported by the National Institutes of Health grants HL-110740 and HL-107957 (Y.W.) and the National Natural Science Foundation of China (81520108004) (H.T.Y.). The authors thank Prof. Xiang Zhang in Genomics, Epigenomics, and Sequencing Core, University of Cincinnati, for RNA-seq assistance.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.