
Abstract
Aims:
Systemic diseases often have common characteristics. The aim of this study was to investigate the feasibility of targeting common pathological metabolism to inhibit the progression of malignant and proliferative diseases.
Results:
Gefitinib-resistant (G-R) nonsmall-cell lung cancer (NSCLC) and rheumatoid arthritis (RA) were studied as conditions representative of malignant and proliferative diseases, respectively. Strong lipogenic activity and high expression of sterol regulatory element-binding protein 1 (SREBP1) were found in both G-R NSCLC cells and synovial fibroblasts from RA patients (RASFs). Berberine (BBR), an effective suppressor of SREBP1 and lipogenesis regulated through reactive oxygen species (ROS)/AMPK pathway, selectively inhibited the growth of G-R NSCLC cells and RASFs but not that of normal cells. It effectively caused mitochondrial dysfunction, activated ROS/AMPK pathway, and finally suppressed cellular lipogenesis and cell proliferation. Addition of ROS blocker, AMPK inhibitor, and palmitic acid significantly reduced the effect of BBR. In an in vivo study, treatment of BBR led to significant inhibition of mouse tumor xenograft growth and remarkably slowed down the development of adjuvant-induced arthritis in rats.
Innovation and Conclusion:
Targeting ROS/AMPK/lipogenesis signaling pathway selectively inhibited the growth of G-R NSCLC cells and the progress of RASFs in vitro and in vivo, which provides a new avenue for treating malignancies and proliferative diseases.
Introduction
M
Targeting lipogenesis through activating reactive oxygen species/AMPK signaling pathway selectively inhibited the growth of gefitinib-resistant nonsmall-cell lung cancer cells and the progress of rheumatoid arthritis synovial fibroblasts in vitro and in vivo. It provides a new avenue for treating malignancies and proliferative diseases.
Moreover, tumor necrosis factor (TNF) has been identified as an important factor in arthritis and inflammatory bowel disease, which may explain why chronic joint inflammation and intestinal diseases are comorbid in many patients (1). Therefore, elucidating the common pathological mechanisms and pathways of various complex diseases will contribute to the development of novel therapeutic strategies. For example, targeting reactive oxygen species (ROS) and the synthesis of biomass have attracted global attention. Increased ROS production was demonstrated as a common phenotype of many diseases, such as cancer and autoimmune and cardiovascular diseases (24). Therefore, targeting cellular ROS generation is developing as a potential therapeutic target of diseases.
Among three main types of biomass synthesis, de novo fatty acid synthesis is extremely active in fetal lung and lung cancers (29). Lipids play essential roles in maintaining cellular signaling homeostasis (38); the lipid bilayer is the fundamental structure of cellular membranes and promotes stabilization of protein channels and receptors (8). The pathological roles of lipids has recently attracted attention, and multiple reports have indicated that alteration of lipid metabolism is directly and closely associated with the pathogenesis and progression of a variety of diseases, such as rheumatoid arthritis (RA) (16,20), cancer (4, 29), asthma (30), obesity, diabetes (47), and atherosclerosis (2, 11). In addition, several lipid metabolic parameters are potential diagnostic and therapeutic biomarkers in the clinic (5). Obesity is frequently associated with diabetes and cancer; however, the common underlying metabolic disorder in RA and cancer has not yet been identified.
Changes in total cholesterol, low-density lipoprotein cholesterol, and triglyceride levels are generally observed in RA patients (16), while cardiovascular risk is positively correlated with RA progression (3). A population-based study has demonstrated that a significant proportion of RA patient mortality is due to cardiovascular disease (14, 28). Moreover, lipids can exacerbate arthritis progression (6). Finally, treatment with lipid-lowering statins effectively reduces the risk of RA (43). Thus, lipids are closely linked to the pathogenesis and progression of RA. The role of lipid metabolism in cancer development and treatment has been extensively studied (36). First, the physiological components of lipid rafts, which act as anchors to stabilize cellular receptors and transmembrane proteins, could initiate carcinogenesis. For example, the EGFR gene, which is a cancer driver (13), and multidrug resistance receptors, such as ABCG2, depend on lipid rafts for anchoring to the cell membrane and regulate cell response and growth (12, 40). Although noncancerous cells also require energy and lipids to support their growth and differentiation, their dependence on lipogenesis differs from that of malignant cells, which suggests that targeting lipogenesis could be a treatment strategy for malignant or highly proliferative diseases, such as nonsmall-cell lung cancer (NSCLC) and RA, in humans. Furthermore, the anticancer efficacy of CD8+ T cells has been shown to be significantly enhanced by modulating cholesterol metabolism (49).
In the current study, the role of lipogenesis in both malignant and proliferative diseases was assessed concurrently for the first time, and the potential therapeutic effect of targeting lipid metabolism was comprehensively examined in both in vivo and in vitro models of RA and gefitinib-resistant (G-R) NSCLC. The widely prescribed drug metformin and an active compound derived from Chinese medicinal plants, berberine (BBR), were used to assess this hypothesis.
Results
Sterol regulatory element-binding protein 1 functions as a biomarker and potential therapeutic target for malignant and highly proliferative diseases
To assess lipogenesis in malignant and highly proliferative diseases, we used NSCLC and RA as representative models for analysis. By comparing the expression of critical factors and enzymes associated with lipogenesis in different cell types from these two model diseases, we found that sterol regulatory element-binding protein 1 (SREBP1), a key positive regulator of sterol synthesis, was highly expressed in both NSCLC cells and MH7A human synovial fibroblasts following TNF-α stimulation but was poorly expressed in normal BEAS-2B lung epithelial cells or unstimulated human synovial fibroblasts (Fig. 1A, B). Among these NSCLC cell lines, H1975 and H1650 are G-R, HCC827 is gefitinib sensitive, and A549 and H2228 harbor wild-type EGFR that is not applicable for gefitinib treatment. Thus, we hypothesized that suppressing SREBP1 and lipogenesis can inhibit the progression of malignant and proliferative diseases. Metformin (25) and BBR (48), which have previously been identified to be effective suppressors of SREBP1, were used to confirm our hypothesis, and their inhibitory effects were compared. Interestingly, although metformin has been marketed globally, BBR showed much stronger cytotoxic effects on G-R NSCLC H1975 cells than did metformin (Fig. 1C); thus, we selected BBR as the representative drug in the following experiments.

First, the inhibitory effect of BBR on lipogenesis was validated. G-R NSCLC cell lines H1975 and H1650 with the highest SREBP1 expression were selected for further studies. BBR significantly suppressed the expression of SREBP1 in the H1975 (3 μM) and H1650 (6 μM), the human fibroblast cell line MH7A (40 μM), and synovial fibroblasts (80 μM) from RA patients (RASFs) (Fig. 1D). We found that BBR substantially reduced the triglyceride, cholesterol, and other lipid metabolite levels in H1975 cells (Supplementary Fig. S1A–C; Supplementary Data are available online at
IC50 Values of Berberine on Synovial Fibroblasts
BBR, berberine; RASF, rheumatoid arthritis synovial fibroblast; TGF, transforming growth factor.
Furthermore, we used both an adjuvant-induced arthritis (AIA) model in rats and a G-R NSCLC xenograft model of cancer in mice to verify the effects of BBR on suppressing malignant carcinoma and proliferative conditions in vivo. In the AIA model, the levels of SREBP1, TNF-α, and interleukin (IL)-β were all significantly increased (Supplementary Fig. S1D), and an increase in hind paw volume was considered a major indicator of arthritis. Methotrexate (MTX), the most commonly prescribed drug worldwide for treating RA, and metformin, a drug marketed for treating diabetes and cancer, were used as positive controls (33). Notably, a significant regression in edema of the hind paw was observed in AIA animals treated with 25 mg/kg BBR compared to that of the vehicle-treated AIA rats, and this regression was similar to the effects of MTX and metformin (Fig. 2A and Supplementary Fig. S1E). However, 25 mg/kg metformin showed no inhibitory effect, indicating that BBR is more effective than metformin. Moreover, bone damage induced by RA was effectively blocked by BBR at 25 mg/kg concentration (Fig. 2B). In the mouse G-R NSCLC xenograft model, at 10 mg/kg BBR successfully suppressed tumor growth (Fig. 2C), and this was almost as efficacious as the positive control drug afatinib. Both tumor volume and weight in BBR-treated animals were significantly decreased, to one half of that of the control animals. We further analyzed EGFR and SREBP1 expression in tumor xenograft samples (Fig. 2D), as well as the levels of lipid metabolites, and found that expression of EGFR and SREBP1 was significantly decreased, as well as the cellular and plasma levels of the lipid metabolites palmitic acid and mevalonic acid were substantially reduced by BBR treatment (Fig. 2E).
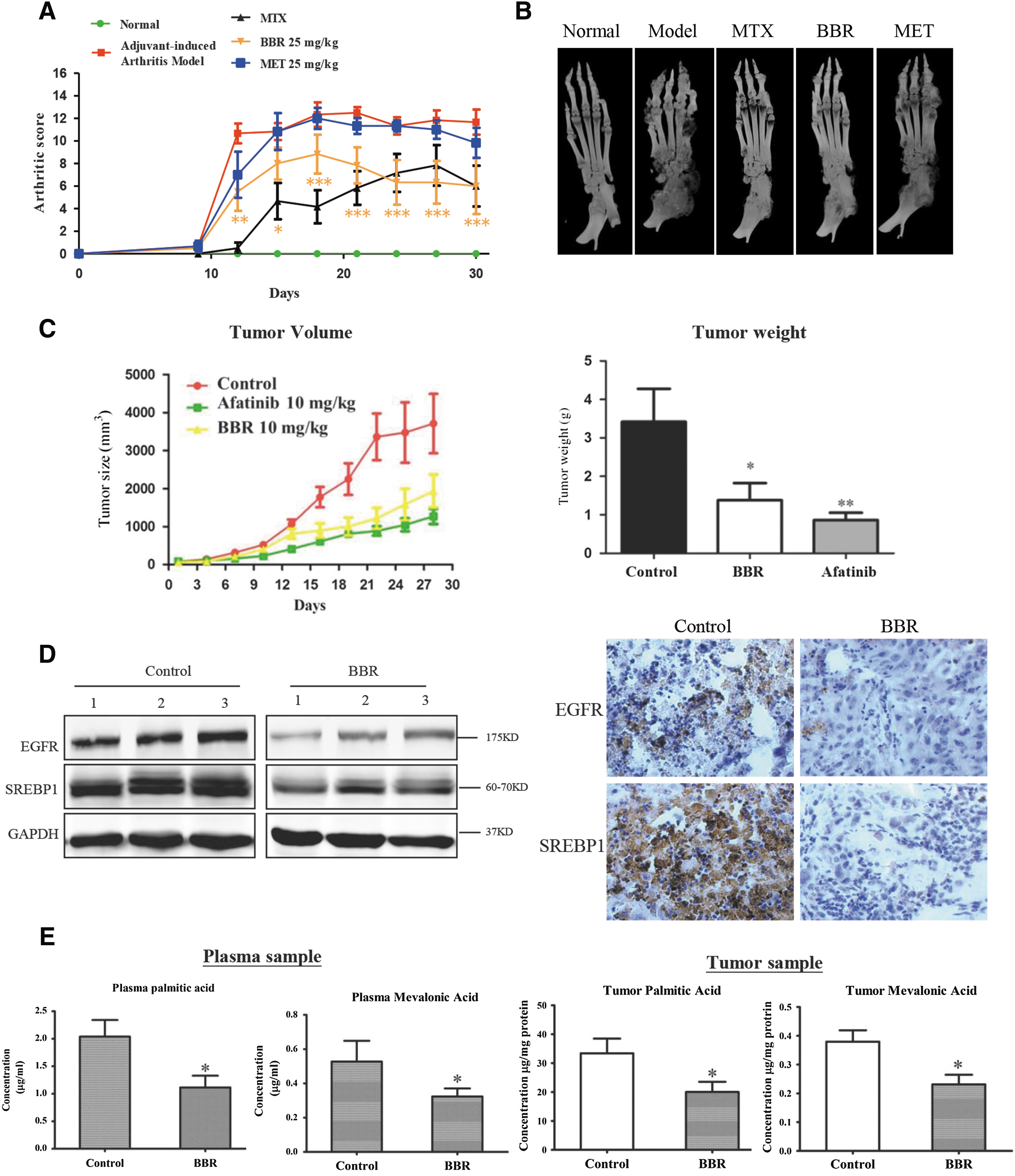
Taken together, the results showed that BBR effectively inhibited the progression of both arthritis and growth of G-R NSCLC in vivo. Importantly, the level of SREBP1, a key transcription factor for regulating genes associated with de novo lipogenesis, was increased in both G-R NSCLC and RA cells, as well as in the in vivo model, and BBR significantly decreased SREBP1 expression and the levels of its downstream lipid metabolites, indicating that suppressing SREBP1 and lipid metabolism can substantially inhibit both malignant and proliferative diseases.
Inhibition of lipogenesis selectively suppresses the growth of human synovial fibroblasts and G-R NSCLC cells
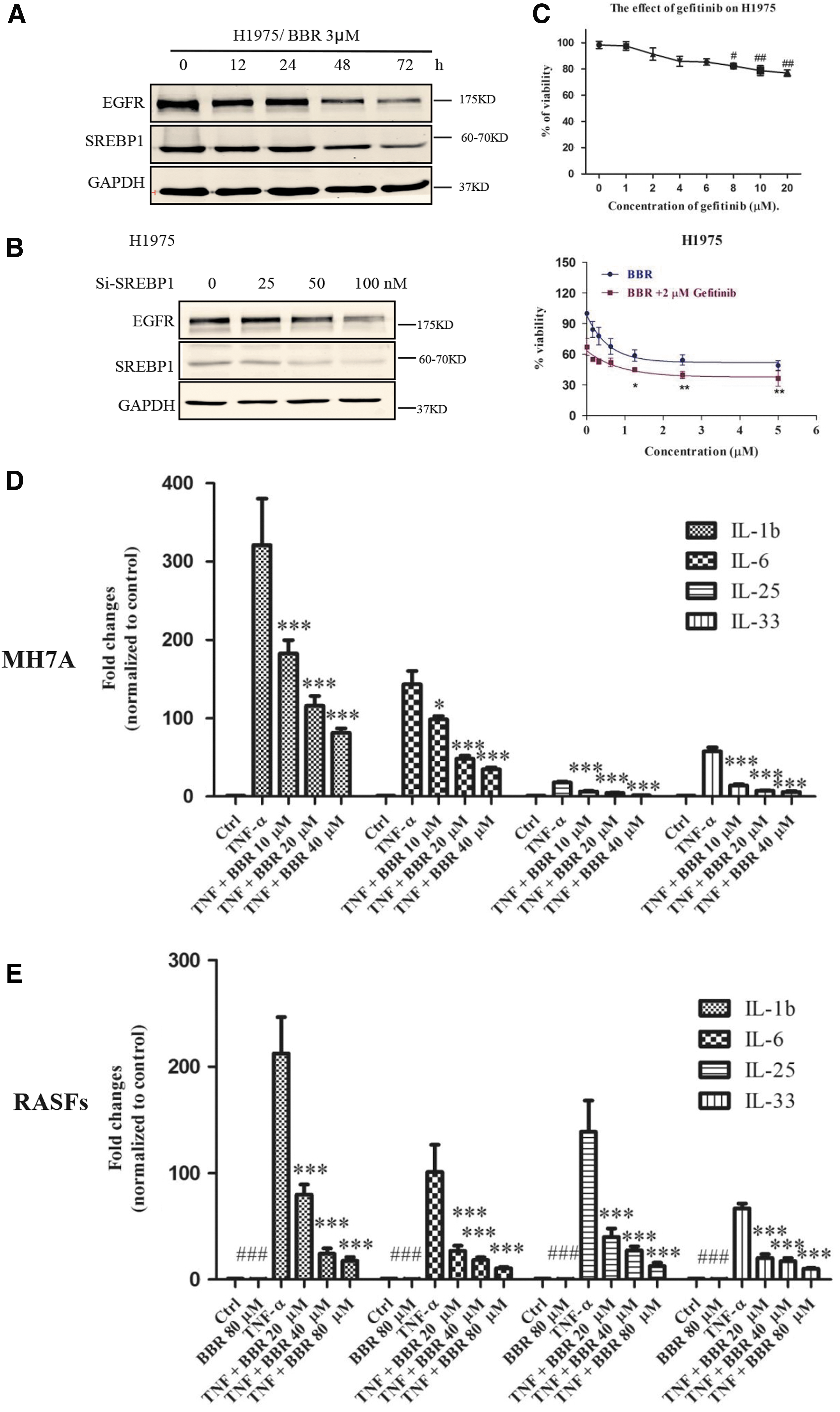
To determine whether targeting lipogenesis indiscriminately affects normal cells, fibroblasts, and malignant cells, we compared the cytotoxicity toward the normal and abnormal cells in both animal models. First, the cytotoxic effect of BBR was compared among different types of lung cancer cells (Table 2 and Supplementary Fig. S2). Intriguingly, BBR showed a strong cytotoxic effect on G-R NSCLC cells but not on normal cells: the IC50 value of BBR toward BEAS-2B normal lung epithelial cells was ∼10-fold higher than that toward H1975 G-R NSCLC cells containing a double mutation (L858R and T790M) in EGFR. These results suggest that BBR may be specifically cytotoxic toward G-R NSCLC cells. To determine why BBR is selective and has the strongest effect on G-R NSCLC cells, we hypothesized that BBR inhibits EGFR signaling by suppressing lipogenesis. First, EGFR expression was investigated. We found that the level of EGFR was in accordance with that of SREBP1, which was significantly downregulated by BBR (Fig. 3A). To further clarify whether downregulation of EGFR is mediated through suppression of lipogenesis, we used siRNA to knock down SREBP1. Knockdown of SREBP1 significantly reduced the protein level of EGFR (Fig. 3B). Moreover, a combination of BBR and gefitinib showed interactive effects on H1975 cells (Fig. 3C), which indicated that BBR could be combined to use with gefitinib in G-R NSCLC cells.
IC50 Values of Berberine on Nonsmall-Cell Lung Cancer Cells
To assess the effect of BBR on RA, we used MH7A cells and RASFs stimulated with TNF-α to induce generation of the proinflammatory cytokines IL-1β, IL-6, IL-8, IL-25, and IL-33. The results demonstrated that BBR effectively attenuated the upregulation of inflammatory cytokines induced by TNF-α (Fig. 3D, E, and Supplementary Fig. S3).
Next, the essential role of suppression of lipogenesis in the anticancer or antiarthritic effects mediated by BBR was examined. A critical intermediate metabolite of lipogenesis, palmitic acid, was applied with BBR to the cell culture, and the results showed that coadministration of palmitic acid significantly relieved the cytotoxic effect of BBR on G-R NSCLC cells and RASFs (Fig. 4A–D). On the basis of these results, we concluded that BBR effectively inhibited lipogenesis and thus selectively suppressed the growth of G-R NSCLC cells and the inflammation mediated by human synovial fibroblasts.

The mitochondrial respiratory chain is crucially involved in the suppressive effects of BBR on both human fibroblasts and malignant cells
Since BBR effectively suppressed lipogenesis, to further explore the candidate targets for regulating lipogenesis, we first determined the effect of BBR on apoptosis and the cell cycle. BBR did not significantly increase the percentages of apoptotic cells in the H1975 cell population (Supplementary Fig. S4A), while it caused significant G2 arrest in both G-R NSCLC cells and RASFs (Fig. 4E, F, and Supplementary Fig. S4B, C), indicating that BBR suppressed the growth of G-R NSCLC carcinoma and the proliferation of human synovial fibroblasts through induction of cell cycle arrest rather than apoptosis. Since lipogenesis is directly involved in cell cycle progression (18), these results are consistent with and are a further confirmation of the suppressive effect of BBR on lipogenesis.
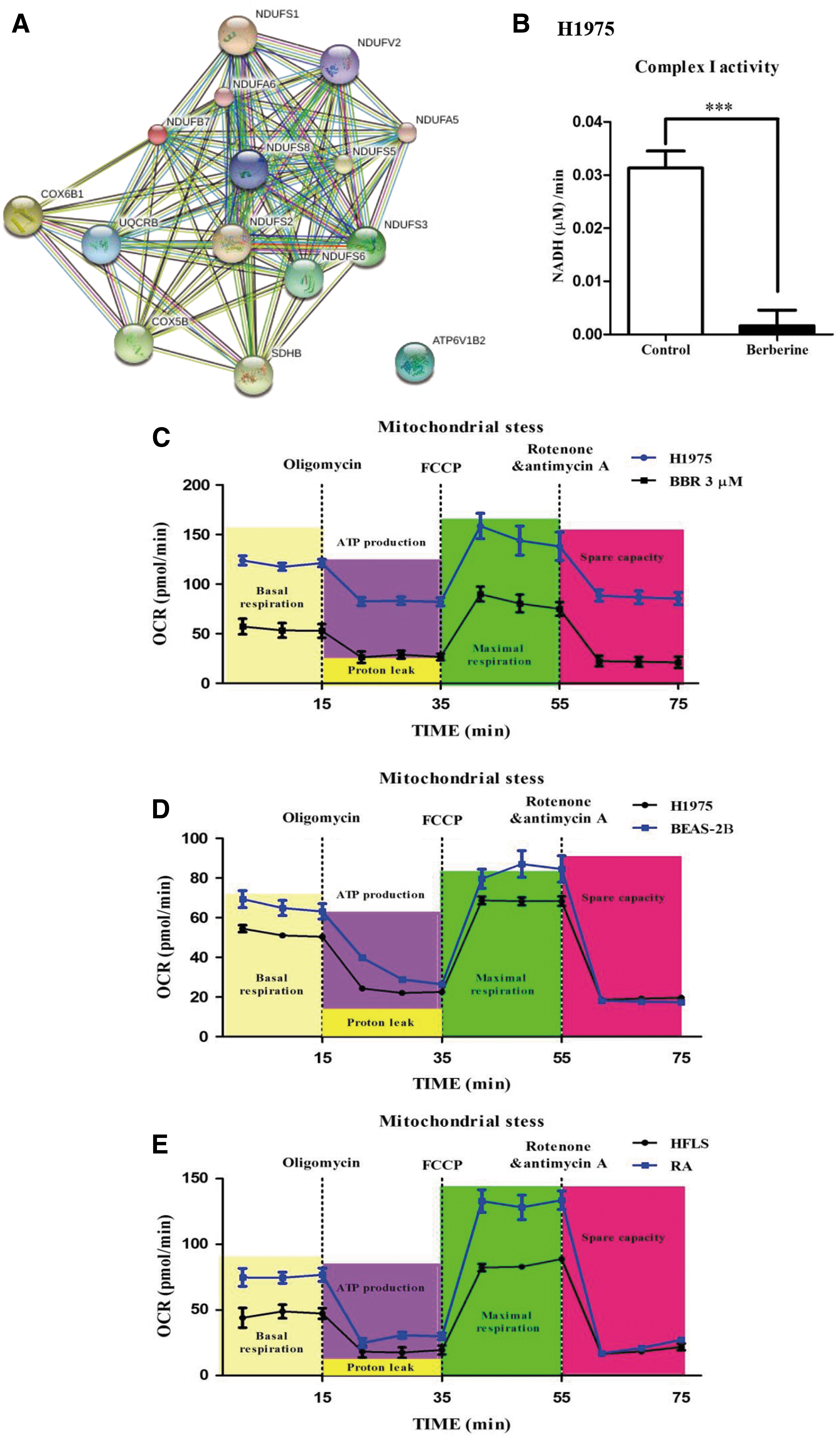
To identify the potential targets of BBR, we performed iTRAQ proteomic analysis. The results demonstrated that 107 genes showed a more than twofold change in expression following BBR treatment (3 μM). Of these, 61 genes were downregulated, while 46 genes were upregulated (Table 3). Bioinformatic analysis of these genes indicated that 15 genes were associated with the mitochondrial respiratory chain, which was the center of the connection network generated from the data and was significantly suppressed by BBR (Fig. 5A and Supplementary Fig. S5). Enzyme activity assays of complex I in the mitochondrial respiratory chain also demonstrated that BBR significantly inhibited the activity of complex I (Fig. 5B).
Proteomic Analysis of Berberine Targets
The list of proteins whose levels changed by more than twofold.
Since the mitochondrial respiratory chain involves oxygen consumption and energy production, to clarify whether the suppression of the mitochondrial respiratory chain is the major cause of selectivity of BBR on different types of cells, we compared the oxygen consumption rate (OCR) of the cells. OCR is a critical indicator of mitochondrial function and directly reflects oxidative phosphorylation (OXPHOS) of the mitochondria, which are the major source of ATP for eukaryotic cells. First, compared with the control H1975 cells, treatment with BBR (3 μM) dramatically decreased the OCR and extracellular acidification rate (ECR) of the cells (Fig. 5C and Supplementary Fig. S6A). Moreover, the basal OCRs of BEAS-2B (normal lung epithelial cells) and H1975 cells were compared (Fig. 5D). Interestingly, H1975 cells had a higher OCR than did BEAS-2B cells, which is consistent with the cytotoxic effects of BBR. Similarly, a much higher OCR was also observed in RASFs than in non-RA fibroblasts (primary human fibroblast like synoviocytes [HFLS] cells derived from normal synovial tissues) (Fig. 5E). Higher rates of oxygen consumption by the cells indicated increased sensitivity of the cells to BBR treatment.
Mitochondria are the major organelles involved in energy and ROS generation. Dysfunction of mitochondria will lead to energy deprivation and ROS accumulation (15). Therefore, we first examined whether BBR caused ATP reduction. As expected, the cellular ATP level in BBR-treated H1975 cells was significantly decreased (Supplementary Fig. S6B). It is well established that nutrient deprivation or high levels of stress can increase mitochondrial fission and/or decrease fusion (45). Second, the cellular ROS level was analyzed and compared. BBR notably enhanced the ROS generation within 3 h (Supplementary Fig. S7A). Collectively, our findings show that BBR inhibits lipogenesis mainly by targeting AMPK pathway and thus suppresses the growth of NSCLC and the inflammatory responses elicited by RA fibroblasts.
ROS–AMPK activation signaling is required for the inhibitory effects of BBR on both human synovial fibroblasts and malignant cells by suppressing lipogenesis
AMPK is an energy sensor and regulates energy homeostasis of cells by detecting changes in the AMP/ATP ratio. As expected, the deprivation of ATP by BBR directly activated AMPK and its direct downstream pathway, which involves Acetyl-CoA carboxylase, even at 3 h (Fig. 6A, B). Moreover, treatment with the AMPK inhibitor compound C (10 μM) significantly blocked the inhibitory effect of BBR (Fig. 6C–F), whereas overexpression of AMPK (to simulate the effect of AMPK activation) significantly suppressed cellular viability; a dominant negative mutant of AMPK was used as a control (Fig. 6G). Since BBR could effectively induce AMPK activation and ROS generation, we explored whether there are direct associations involved. Cells were cotreated with BBR (3 μM) and ROS blocker N-acetylcysteine (NAC, 10 mM). Interestingly, the activation of AMPK induced by BBR was mostly counteracted by NAC, which indicated that the activation of AMPK was largely dependent on the stimulation of ROS increasing (Supplementary Fig. S7B). These results suggested that ROS increasing and AMPK activation are essential requirements for the anticancer and antiarthritic effects of BBR.

Discussion
Lipids are composed of multiple hydrophobic molecules and have many crucial roles (9). The important physiological roles of lipids can be divided into four categories: lipids are a source and store of energy, major structural components of biological membranes, the building blocks for the synthesis of several hormones (chemical messengers) and bile acids, and components of the nervous system. From the perspective of pathology, reports showing that many diseases have specific alterations in lipid metabolism are increasing dramatically (27, 34). Changes in lipid metabolism are involved in numerous cellular processes, including the signal transduction that regulates cell growth and the synthesis of membranes, as well as energy homeostasis (36) and inflammation (39).
We found that lipogenic activity was increased in NSCLC cells and RA fibroblasts in both cell culture and in vivo animal models (Figs. 1 and 2), indicating a pathophysiological mechanism of malignancy and highly proliferative growth. Therefore, suppression of lipogenesis led to selective inhibition of the growth of malignant or hyperproliferative cells. Moreover, the expression of cancer driver genes, such as EGFR, was strongly associated with lipogenesis in cells. EGFR is a transmembrane receptor and localizes on cellular membrane to exert its signaling transduction. To stabilize on cell membrane, EGFR requires an anchor for fixation. The anchor is also named lipid raft, which mainly comprised cholesterol, sphingolipids, and gangliosides (37). It has been well established that the levels of lipid rafts are increased in various cancers and play a functional role during tumorigenesis (26, 31). Failure to stabilize cellular EGFR will lead to EGFR ubiquitination and degradation. Consequently, knockdown of SREBP1, which is a key positive regulator of cellular sterol biosynthesis, substantially reduced the level of EGFR protein and induced cell death, thus significantly sensitizing G-R NSCLC cells to gefitinib. Therefore, targeting lipogenesis is a promising strategy to treat malignant and highly proliferative diseases and may be used to sensitize cells to drugs via induction of receptor degradation (Fig. 7).
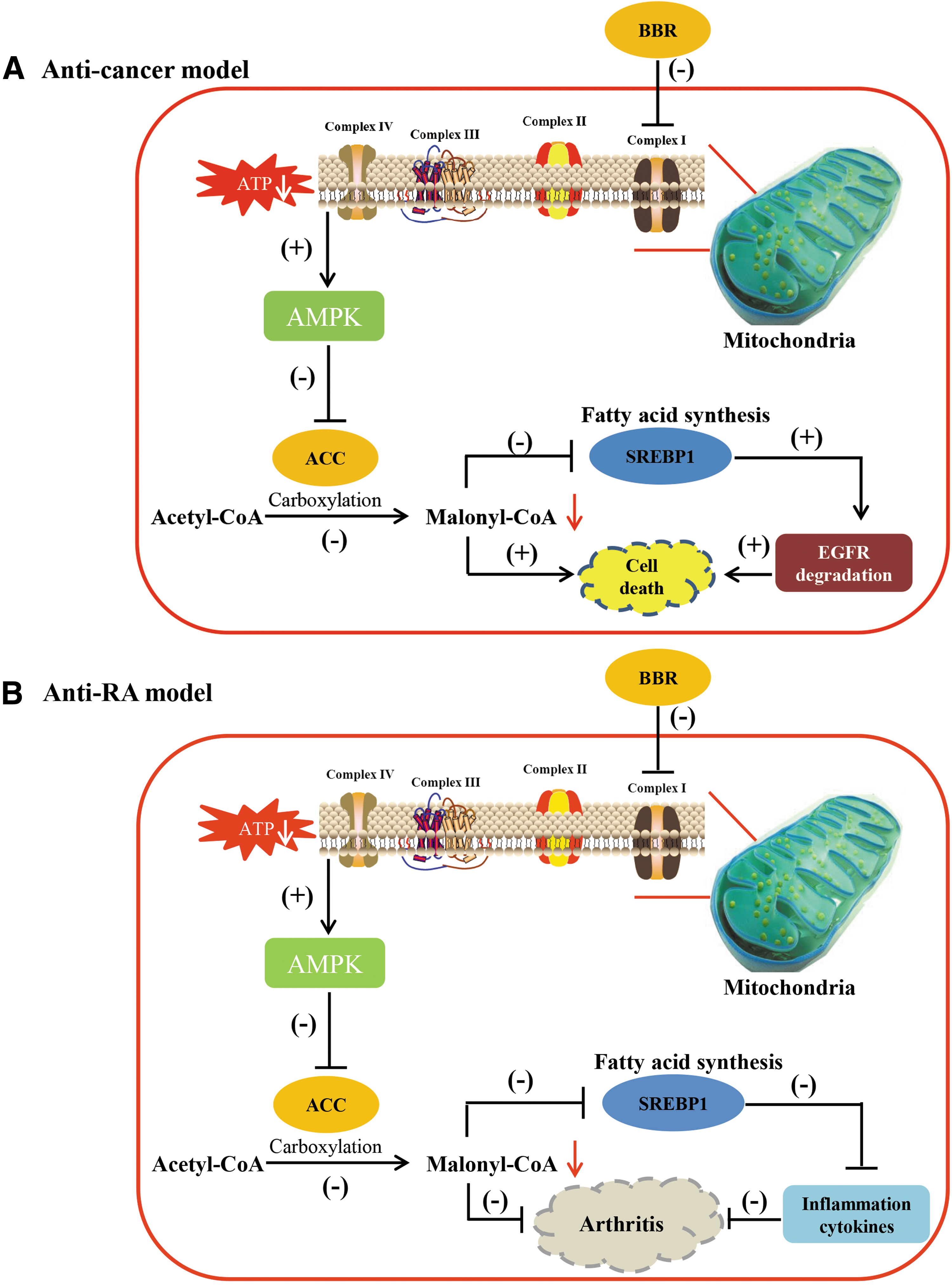
Chinese herbal medicines have received increased attention as a source of new drugs (42). BBR has been reported to be an isoquinoline alkaloid isolated from Coptidis Rhizoma (Huanglian), which has diverse pharmacological activities, including detoxification, antidiabetic, anti-inflammatory (10, 41, 52), and anticancer properties (32, 51). Moreover, it was identified as a potent cholesterol-lowering agent with a unique mechanism distinct from that of the cholesterol-lowering drug statins as well (19). For its function on antidiabetes and lipid metabolism, BBR has been clearly elucidated, it even has better efficacy than that of metformin (50).
In this study, we examined whether BBR could suppress lipid metabolism to treat carcinoma and RA for the first time. Previously, BBR was shown to effectively accelerate cellular metabolism; for example, in the treatment of diabetes and obesity, BBR displayed beneficial effects through its activation of AMPK (21). Both in vitro and in vivo studies indicated that the growth of G-R NSCLC cells and RA fibroblasts was remarkably inhibited by BBR, and its anticancer and antiarthritic effects could be largely blocked by addition of intermediate metabolites in the fatty acid and cholesterol synthesis pathways. BBR directly affected the mitochondrial respiratory chain, inhibited OXPHOS, and resulted in ROS accumulation and AMPK activation. Intriguingly, ROS enhancement exerted a critical role in AMPK activation. Blocking ROS generation could mostly weaken the AMPK activation induced by BBR. Moreover, pretreatment with the AMPK inhibitor compound C mostly counteracted the effect of BBR and led to recovery of the viability of cells, which suggested that the activation of ROS/AMPK signaling pathway could effectively suppress lipogenesis and thus cell proliferation, which could be developed as a therapeutic target.
Since the action mechanism of BBR is very similar to that of metformin, which has been identified as an inhibitor of mitochondrial complex I (44, 46), we used it as a parallel control. However, better efficacy was observed in BBR-treated group, and 25 mg/kg of BBR induced significant suppression in the AIA rat model, while metformin at the same dose had no effect in this arthritis model. Attractively, for intragastric oral administration, there is no LD50 of BBR found, which indicated that it is very safe for human use and has great potential for drug development (17).
We also assessed the OCRs of different cell types and found that they varied. H1975 NSCLC cells or RASFs tend to have a higher rate of mitochondrial oxygen consumption than that of normal cells. It is very interesting that OCR is higher in the tumor cells with respect to normal lung epithelial cells. The underlying mechanism may associate with cellular characteristics. H1975 cells contain double mutation on EGFR and depend on EGFR-PI3K-mTOR signaling pathway for survival and growth. This signaling pathway is directly involved in biomass synthesis (nucleotides, lipids, and amino acids), which is mainly performed in mitochondria. Therefore, we speculate that in H1975 cells, mitochondrial function is required. Moreover, due to the rapid growth rate, H1975 cells are much eager for macromolecules than normal cells. Therefore, the EGFR mutation in H1975 cells may be the cause leading to higher OCR of H1975 cells. This metabolic difference could be an Achilles' heel in various diseases and therefore points to a new strategy for treating cancers as well as RA.
In summary, targeting lipid metabolism or upstream molecules has significant beneficial effects in many diseases. The selective action of BBR on cells from diseased tissues or malignant and hyperproliferative cells, but not normal cells, is highly attractive for future clinical study. Moreover, BBR is a strong candidate for drug development to treat cancer and RA by suppressing lipogenesis. Using BBR monotherapy to modulate lipid metabolism may be a novel therapeutic strategy for patients suffering from malignant and highly proliferative diseases.
Materials and Methods
Materials
BBR and metformin powder were purchased from Sigma-Aldrich (St. Louis, MO). Propidium iodide (PI) and annexin V/PI staining dye were purchased from BD Biosciences (San Jose, CA). Cell lysis buffer (RIPA) and primary antibodies against actin, total EGFR, and FAS were purchased from Cell Signaling Technology (Danvers, MA). Fluorescein-conjugated goat anti-rabbit and mouse antibodies were purchased from Odyssey (Lincoln, NE).
Cell lines and cell culture
BEAS-2B, A549, HCC827, H820, H1650, H1975, and H2228 cells were purchased from ATCC (Manassas, VA) and cultivated in RPMI 1640 medium supplemented with 10% fetal bovine serum (Invitrogen, Carlsbad, CA), 100 U/ml penicillin, and 100 μg/ml streptomycin (Invitrogen). The BEAS-2B culture flasks were precoated with a mixture of 0.01 mg/ml fibronectin, 0.03 mg/ml bovine collagen type I, and 0.01 mg/ml bovine serum albumin dissolved in bronchial epithelial basal medium (BEBM) medium (Lonza, Allendale, NJ). MH7A, a human RA fibroblast line, was purchased from ATCC and cultured in Dulbecco's modified Eagle's medium (DMEM). RASFs are primary RA fibroblasts isolated from RA patients and cultured in DMEM supplemented with 10% fetal bovine serum. HFLS cells were purchased from Cell Application and cultured in Human Synoviocyte Growth Medium. All the cells were cultivated at 37°C in a 5% CO2 incubator. In the cotreatment experiments, the cells were pretreated with compound C for 1 h and then cotreated with BBR.
3-(4, 5-Dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide assay
Cells were seeded in a 96-well microplate at 5000 cells/well and cultured overnight for cell adhesion. Various concentrations of BBR were added with the vehicle control, dimethyl sulfoxide (DMSO), and the microplates were incubated for 24 or 48 h. Each dose was assayed in triplicate. 3-(4, 5-Dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (5 mg/ml, 10 μl) solution was added to every well, and the plate was incubated for 4 h. Then, 100 μl of solution (10% sodium dodecyl sulfate [SDS] and 0.1 mM HCl) was added to each well, and the plate was further incubated at 37°C for another 4 h to dissolve the formazan crystals. Finally, the absorbance of the plate was measured at 570 nm (absorbance) and 650 nm (reference) using a microplate reader (Tecan, Männedorf, Switzerland). The cell viability was calculated as the percentage change in the absorbance of the treated cells divided by the absorbance of the untreated cells.
Assessment of apoptosis via annexin V/PI staining
After treatment, the cells were harvested, washed with phosphate-buffered saline (PBS), and resuspended in 1 × binding buffer. Then, the cells were double stained with annexin V/PI for 15 min at room temperature in the dark. The apoptotic cells were quantified by flow cytometry (BD FACSAria III; BD, Franklin Lakes, NJ).
Cell cycle assessment by PI staining
Cells were seeded into six-well plates with 1 × 105 cells/well and cultured in an incubator overnight for cell adhesion. After treatment with BBR, the cells were trypsinized and centrifuged. The cells were washed with PBS and centrifuged to remove the PBS. The pellet was resuspended and fixed in 70% ethanol for 30 min at 4°C. After centrifugation, the cells were stained with PI for 30 min and then analyzed by flow cytometer.
ROS detection
After treatment with different concentrations of BBR for 3 h, cells were stained with DCFDA for 15 min and loaded onto flow cytometer.
Western blot analysis
Cells were lysed in RIPA lysis buffer with protease and phosphatase inhibitors to extract the total protein. The concentration of the total protein extract was determined with a Bio-Rad DCTM Protein Assay Kit (Bio-Rad, Hercules, CA). Then, 30 μg of total protein lysate was loaded onto a 10% SDS-polyacrylamide gel electrophoresis gel; the separated proteins were transferred to a nitrocellulose membrane. Membranes were blocked with 5% milk without fat in TBST for 1 h at room temperature. Primary antibodies (Santa Cruz Biotechnology; 1:500 dilution; Invitrogen and Cell Signaling Technology; 1:1000 dilution) were incubated overnight at 4°C. After the membrane was washed three times with TBST (5 min/wash), a secondary fluorescent antibody (1:10,000 dilutions) was added to the membrane at room temperature for 1 h. Actin was used as the loading control and for normalization. The signal intensity of the membranes was detected with the LI-COR Odyssey Scanner (Belfast, ME).
RNA extraction
RNA was extracted from NSCLC cells with an RNA extraction kit (Invitrogen) according to the manufacturer's instructions. After the medium was removed, 0.3 or 0.6 ml lysis buffer with 1% 2-mercaptoethanol was added to the cells. The total cell lysate was transferred to an RNase-free tube and vortexed until the cell pellet was dispersed. One volume of 70% ethanol was added to the cell lysate and then mixed thoroughly to disperse any visible precipitate. The mixture was transferred to a spin cartridge and centrifuged at 12,000 g for 15 s at room temperature to discard the flow-through. The spin cartridge was sequentially washed with wash buffer I and wash buffer II. Finally, RNA was eluted with 50 μl of RNase-free water. The RNA concentration was determined using Nano2000, and RNA quality was assessed by electrophoresis.
cDNA synthesis
The synthesis of first-strand cDNA was carried out following the instructions of the cDNA synthesis kit (Roche, Basel, Switzerland). One microgram of DNase-treated RNA was used for each cDNA synthesis. First, RNA sample was incubated with 2 μl of random primers and 1 μl of oligo (dT) primer provided with the kit at 65°C for 10 min and then cooled on ice for 2 min. The reaction mixture, which contained reaction buffer, RNase inhibitor, and reverse transcriptase, was added to the tube and then incubated at 25°C for 10 min and 55°C for 30 min. The reaction was terminated by heating at 70°C for 15 min. The final product (cDNA) was stored at −80°C until further use.
Polymerase chain reaction and real-time polymerase chain reaction
Polymerase chain reaction (PCR) was performed following the instructions of the FastTaq DNA Polymerase Kit (Roche). Each PCR consisted of 1 × PCR buffer, 200 μM dNTP, 200 nM forward primer, 200 nM reverse primer, 0.2 μl Taq, and 50 ng cDNA template in a total volume of 25 μl. The PCR running protocol is as follows: 94°C for 5 min, followed by 35 cycles of 94°C for 30 s, annealing for 30 s, 72°C polymerization for 1 min, and a final extension at 72°C for 10 min. Real-time PCR was performed using FastStart Universal SYBR Green Master. The reaction mixture preparation consist of 10 μl of SYBR Master Mix, 1 μl forward and reverse primers, 0.2 μl template, and water to a final volume of 20 μl. The PCR protocol was 94°C for 10 min, followed by 40 cycles at 94°C for 10 s and 60°C for 30 s.
Transfection
siRNA transfections were performed with Lipofectamine™ LTX Reagent (Invitrogen). 1.5 × 105cells were seeded to each well of six-well plate 1 day before transfection. The transfection complexes were prepared as follows: 100 μM siRNA was diluted in 100 μl Opti-MEM® I Reduced Serum Medium (Invitrogen) and mixed gently. Next, 4 μl LTX Reagent was directly added to each diluted solution sample and incubated for 5–15 min at room temperature. Finally, the transfection mixture was added to the well, mixed gently by rocking the plate back and forth.
Automated liquid chromatography-mass spectrometry/mass spectrometry analysis of the peptides
After collection, protein lysate was trypsin digested, and then the peptide was analyzed using ultrahigh-performance liquid chromatography with an Agilent 1290 Infinity system (Agilent, Santa Clara, CA) as previously reported (23). 0.1% formic acid in Milli-Q water (A) and 0.1% formic acid in acetonitrile (B) consist of mobile phase. Chromatography was performed on an ACQUITY UPLC® BEH C18 column (2.1 × 100 mm, 1.7 μm) and performed on Waters Mass spectrometry (Milford, MA). After acquiring raw data, the results were analyzed by Agilent Bioconfirm protein deconvolution software (Agilent).
OCR assay
Initially, 3000–5000 cells were suspended in 80 μl medium and seeded in each well of a Seahorse plate from B–G; wells A and H contained 80 μl medium without cells. Then, the plate was incubated at 37°C for overnight adhesion. BBR was added to wells E–G for 24 h, while wells A–D contained DMSO, as the control. After 24 h, the culture medium in all wells of the plate was removed and replaced by 80 μl assay medium. Oligomycin (a complex V inhibitor) at 10 μM, carbonyl cyanide-4-(trifluoromethoxy) phenylhydrazone (FCCP) (a protonophore) at 0.5 μM, and antimycin A and rotenone (inhibitors of complex III and I) at 0.5 μM were added to the kit pack according to the user manual. After calibration, the mitochondrial OCR of cells was detected by an Extracellular Flux Analyzer (Seahorse Bioscience, MA).
Targeted metabolomics using the UPLC/MS
The plasma or homogenized tumor sample (200 μl) was mixed with 200 μl of 10 mM n-ethylmaleimide in PBS buffer and 1000 μl of methanol containing 10 ng/ml internal standard
The LC-MS analysis was carried out with ACQUITYTM ultra-Performance LC (Waters) equipped AB 4000 Q-TRAP mass spectrum (AB Sciex). A Waters X BridgeTM BEH C18 analytical column XP (2.5 μm, 3.0 × 100 mm; Waters, Torrance, CA) was selected for LC-MS/MS metabolomics analysis. A linear gradient mobile phase composed of 0.1% formic acid water (solvent A) and methanol (solvent B) was used mixed with a 25-min gradient program: 0–3.0 min (0–1% B); 3.0–10.0 min (1–3% B); 10.0–14.0 min (3–50% B); 14.0–18.0 min (50–95% B); and 18.0–22.0 min (95–0% B). The injection volume was 10 ml and flow rate was 0.6 ml/min. Multiple reaction monitoring detection mode was used for detection with electrospray source (ESI) in both positive and negative ion modes. The m/z 171.1 → 125.2 was used to quantify internal standard Phe-d5. The detailed MS condition included ion spray voltage at ±4500 V; curtain gas at 20 psi; source temperature at 450°C; ion source gas 1 at 40 psi; and ion source gas 2 at 40 psi. The UPLC–MS data were collected by AB Analyst Software (Version 1.6.2).
Method validation for metabolic assay with quality control samples
The standard stock solutions and internal standard stock solution of Phe-d5 were prepared at 1 mg/ml in 100% methanol and stored at −40°C. Blank plasma used for quality control (QC) samples was prepared from mice plasma, which has been stripped of endogenous materials by adding 6 g/100 ml charcoal activated powder (Sigma-Aldrich, MO). This suspension was stirred at room temperature for 2 h and centrifuged for 20 min at 13,500 rpm at 4°C. The supernatant was further filtered using a Millipore Express PES Membrane (Merck Millipore Ltd., Germany). The obtained blank plasma was confirmed to be free of biomarker by LC-MS/MS. Then, the QC samples with three different concentrations were generated by spiking standard solutions to the blank plasma and were processed as mentioned above. The standard curves with eight different concentrations were generated in the identical way as QC samples. Validation experiments were carried out for evaluation of the precision and accuracy of developed method. Moreover, a series of QC samples as well as standard curves were running at 50 sample intervals.
Animal model of adjuvant-induced arthritis
Arthritis model was induced by injection with complete Freund's adjuvant, which contains 0.2 mg of mycobacterium tuberculosis H37Ra and mineral oils through the base of the tail. Control groups were treated with an equal volume of saline solution, and other groups were treated as following: positive control group: 7.6 mg/kg MTX; BBR 25 mg/kg; metformin (MET) 25 mg/kg and administered by oral gavage. Treatments were given daily from the first injection for a period of 30 days.
Xenograft model
The xenograft model was established as previously described (23). Briefly, 1 × 106 H1975 cells/100 μl were mixed with 50 μl Matrigel and then implanted into the right forelimb of each nude mice. After the diameter of tumors reached 5 mm3, the mice were to be treated with BBR (10 mg/kg) and afatinib (10 mg/kg) 5 days/week through oral administration. The tumor volume was calculated by the following equation: volume = (width2 × length)/2.
Statistical analyses
All data are expressed as mean ± standard error of the mean of three individual experiments. Differences between groups were determined using a one-way analysis of variance using GraphPad Prism 5 (GraphPad Software, La Jolla, CA), followed by Bonferroni's test to compare all pairs. Student's t test was used to compare two groups. The level of significance was set at p < 0.05 for all tests.
Footnotes
Acknowledgment
This work was supported by FDCT grants from the Science and Technology Development Fund of Macao (Project code: 086/2015/A3 and 005/2014/AMJ) grant to E.L.-H.L.
Authors' Contributions
E.L.-H.L., Y.X., and X.-X.F. performed the research and analyzed the data. Z.-W.Y., J.L.W., and R.K.-T.K. performed and analyzed the mass spectrometry. Y.F.Z., L.L.L., Y.-L.Z., and Z.B.J. performed animal study. C.-L.W., J.H., and D.K.X. performed real-time PCR of samples. L.L., X.J.Y., Z.Q.L., J.J.F., and J.-X.H. planned and supervised the experiments. X.-X.F. and E.L.-H.L. wrote the manuscript. All authors revised and approved the manuscript.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.