
Abstract
Significance:
Renal transplantation is the treatment of choice for end-stage renal disease, during which renal grafts from deceased donors are routinely cold stored to suppress metabolic demand and thereby limit ischemic injury. However, prolonged cold storage, followed by reperfusion, induces extensive tissue damage termed cold ischemia/reperfusion injury (IRI) and puts the graft at risk of both early and late rejection.
Recent Advances:
Deep hibernators constitute a natural model of coping with cold IRI as they regularly alternate between 4°C and 37°C. Recently, endogenous hydrogen sulfide (H2S), a gas with a characteristic rotten egg smell, has been implicated in organ protection in hibernation.
Critical Issues:
In renal transplantation, H2S also seems to confer cytoprotection by lowering metabolism, thereby creating a hibernation-like environment, and increasing preservation time while allowing cellular processes of preservation of homeostasis and tissue remodeling to take place, thus increasing renal graft survival.
Future Directions:
Although the underlying cellular and molecular mechanisms of organ protection during hibernation have not been fully explored, mammalian hibernation may offer a great clinical promise to safely cold store and reperfuse donor organs. In this review, we first discuss mammalian hibernation as a natural model of cold organ preservation with reference to the kidney and highlight the involvement of H2S during hibernation. Next, we present recent developments on the protective effects and mechanisms of exogenous and endogenous H2S in preclinical models of transplant IRI and evaluate the potential of H2S therapy in organ preservation as great promise for renal transplant recipients in the future. Antioxid. Redox Signal. 28, 1503–1515.
Introduction
R
The mechanisms underlying cold IRI are complex and not fully elucidated. Some studies have reported that such events include mitochondrial injury evidenced by depletion of mitochondrial adenosine triphosphate (ATP) leading to loss of cellular homeostasis and thereby affecting several cellular pathways involved in tissue regeneration and repair (68, 78). Furthermore, excess production of free radicals in the mitochondria seems to play a key role in cold-induced mitochondrial injury (59) and may lead to inflammation and apoptosis of glomerular and tubular cells of cold-stored kidneys (55). The restoration of blood supply and temperature in renal graft during reperfusion results in excessive reactive oxygen species (ROS) generation (21, 55), which initiates a cascade of events, including activation of inflammatory and apoptotic pathways, leading to tissue injury (57). Unfortunately, the search for therapeutic strategies to mitigate cold IRI has met little success so far. Thus, the quest for safe reduction of metabolism in transplant organs continues.
It is of interest to note that mammalian hibernators are able to survive periods of low metabolic rate and body temperature without signs of renal injury in contrast to nonhibernating species. Hibernators alternate their body temperature between 4°C and 37°C without cold IRI challenge. Recently, hydrogen sulfide (H2S), a gas with a distinctive smell of rotten eggs and the third member of the gasotransmitter family, has been implicated in organ protection in deep hibernation (80) and also seems to confer cytoprotection by lowering metabolism and increasing preservation time and overall increasing renal graft survival after transplantation (2, 30, 33, 34). H2S is produced enzymatically in all mammalian species, including humans (52). Two of these enzymatic pathways are cytosolically catalyzed by cystathionine β-synthase (CBS) and cystathionine γ-lyase (CSE). The third enzymatic pathway requires the mitochondrial enzyme, 3-mercaptopyruvate sulfurtransferase (3-MST), while the fourth pathway involves the peroxisomal enzyme, D-amino acid oxidase (DAO) (74, 89).
Apart from being produced endogenously, there are various forms of exogenous H2S that have been used experimentally in the form of donor molecules. The classic form of inhalation is seriously hampered by toxic effects to bystanders, while sulfide salts (sodium hydrosulfide [NaHS] and Na2S) are short-lasting and may not always reach their target sites, particularly the mitochondria (45). Fortunately, slow-releasing H2S donors such as GYY4137 (51) and mitochondria-targeting slow-releasing H2S donors (AP39 and AP123) have been developed (35, 49, 79) and may enable translation of fundamental findings on H2S in modulating renal IRI into human kidney preservation.
In this review, we first discuss mammalian hibernation as a unique natural model of how nature deals with cold IRI with reference to the kidney as a typical organ highly vulnerable to cold IRI. We also highlight the protective role of H2S during hibernation. Next, we present recent findings on H2S as a gasotransmitter that confers cytoprotection during cold IRI in renal transplantation, suggesting that it may become a novel method of preservation to help improve graft quality and increase the long-term success of renal transplantation in the future.
Lessons Learned from Mammalian Hibernation
Mammalian hibernation may offer a great clinical promise to safely cold store and reperfuse donor organs. Hibernation is an evolved strategy that conserves energy mainly seasonally and confers a significant survival advantage to individuals among select, but diverse, mammalian lineages (67). To save energy demand during winter, mammalian hibernators undergo repetitive cycles of torpor and arousal (12, 26) (Fig. 1). Torpor is a state of reduced metabolism and body temperature usually between 4°C and 10°C, depending on the species (13), and can be viewed as the natural equivalent of cold storage of donor organs. As illustrated in Figure 1, the torpor phase lasting days to weeks is intermittently interrupted by brief periods of arousal during which metabolism and body temperature return to euthermic levels (40). Hence, the arousal phase may represent a natural equivalent of reperfusion after cold storage.

While the body temperature of smaller mammalian hibernators falls drastically during torpor and lasts days to weeks, followed by intermittent arousals, larger hibernators such as bears remain in torpid state at 30–35°C for 5–7 months in a year without eating coupled with decreased metabolism, immobilization, and anuria due to complete reabsorption of the glomerular filtrate (17, 76, 84) (Fig. 2). A more extreme type of torpor is seen in hibernating Arctic ground squirrels, during which core body temperature measures −2.9°C without freezing and metabolism reduced to 1% of euthermic rate (5). Interestingly, whereas torpor and arousal cause no organ injury in all hibernating animals studied thus far (19, 42, 43, 51, 92), cold storage and reperfusion result in extensive organ injury in nonhibernating species (30, 42), including humans.
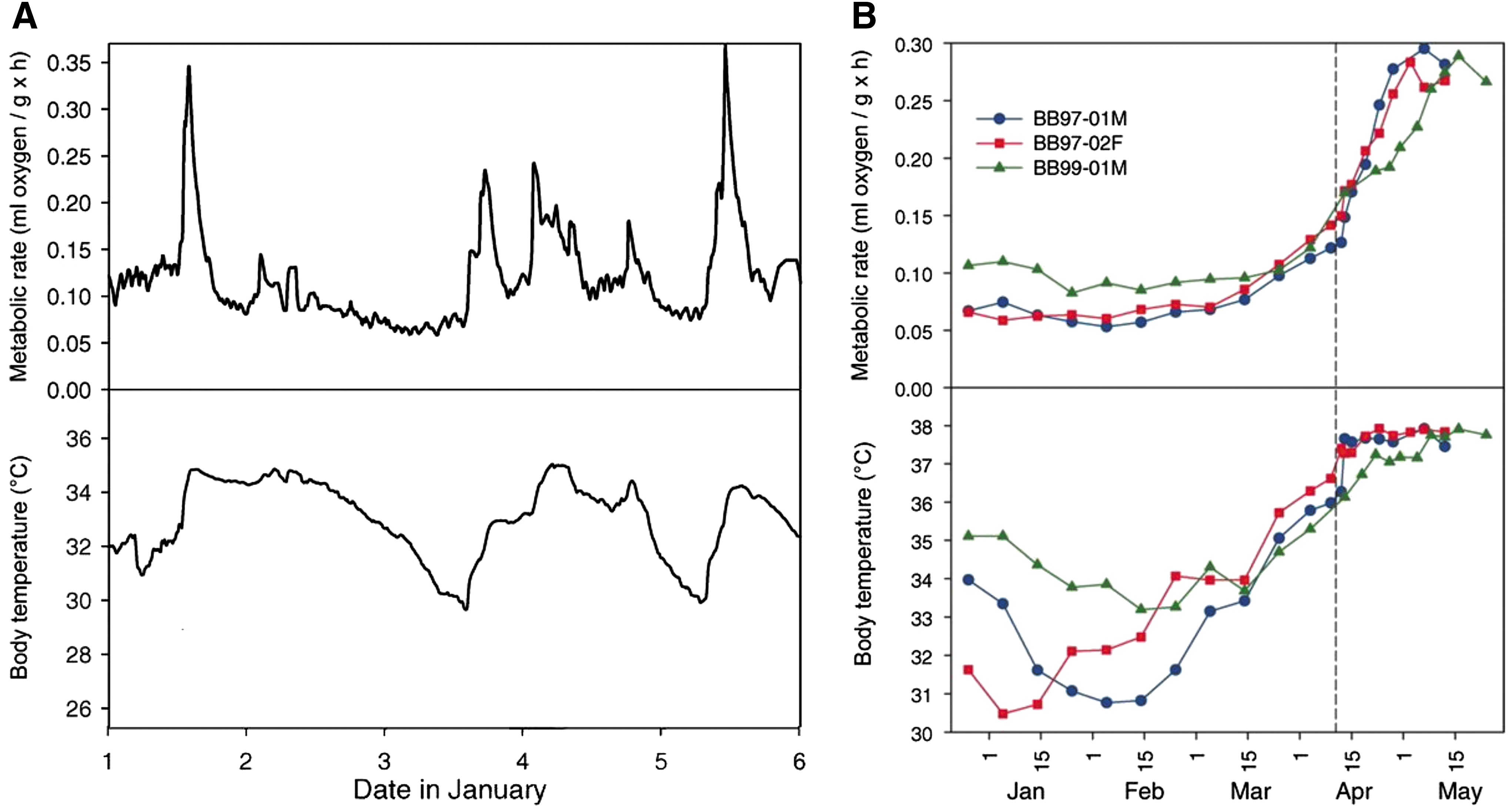
In the kidney, for example, the morphology of the renal cortex and medulla is not affected by hibernation in typical hibernating animals such as 13-lined ground squirrels and dormice, which are the most studied hibernating animals (43, 70, 92). Moreover, the ultrastructure of the glomerular basement membrane, tubular brush border, and cytoskeleton are well preserved throughout torpor and arousal—a condition that would induce extensive damage in human deceased donor kidneys (43, 92). Renal protection in hibernating animals is such a strong intrinsic event that kidneys removed from hibernating 13-lined ground squirrels and stored in cold UW solution resisted apoptotic cell death as shown by reduced caspase-3 activity (42, 43).
The activation and suppression of mitochondrial metabolism and organ protection in hibernating animals seem tightly regulated, and the molecular mechanism of this regulation is not fully understood. Although it is yet to be identified which endogenous molecules are essential for entrance into torpor and/or arousal, H2S has so far proven important in the torpor–arousal cycle as its level is markedly increased in the lung during torpor and reduced in the arousal state in Syrian hamsters (80). This finding suggests that H2S may influence temperature-controlling neurotransmitters such as histamine, serotonin, and opioids, which have been proposed to play key roles in regulation of the torpor pattern by the central nervous system (38, 47, 69) as well as phospholipids of liver cell nuclei (48). Additionally, organ protection in hibernation has been attributed to increased production of endogenous H2S, which can also induce a torpor-like state in small nonhibernating mammals with effects on mitochondrial metabolism (8, 9, 80) and endogenous antioxidants such as glutathione and ascorbic acid (28), as well as upregulation of antiapoptotic genes such as B cell lymphoma-2 (Bcl-2) and phospho-Akt (31, 42). The renoprotective effect of H2S was also observed in rats (nonhibernating animals), in which dopamine treatment upregulated renal expression of CBS, CSE, and 3-MST and stimulated endogenous H2S production, leading to protection against cold renal IRI (30). Dopamine or serotonin treatment of rat smooth muscle aortic cells as well as rat kidney and other tissues before cold storage also protected against hypothermia/rewarming-induced apoptosis and subsequent hypothermic injury by strongly increasing the endogenous H2S level (81).
In addition, our group demonstrated that endogenous H2S production protects kidneys of hamsters (natural hibernators) subjected to 5′-adenosine monophosphate (AMP)-induced torpor-like state at 7°C body temperature (29) (Figs. 3 and 4). Furthermore, Blackstone and Roth demonstrated that pretreatment with inhaled H2S for only 20 min markedly prolonged survival without any apparent detrimental effects in mice exposed to otherwise lethal hypoxia (5% oxygen) (9). It appears that kidneys and tissues of mammalian hibernators are well protected from the damaging effects of hypothermia and reperfusion through upregulated antioxidant pathways and maintenance of mitochondrial homeostasis. These findings suggest that H2S plays an important role in modulating cytoprotective pathways that govern protection against cold IRI and allow maintenance of mitochondrial function throughout torpor and arousal.


Do Organs of Hibernators Suffer Ischemia During Torpor or Is There an Element of Organ Preservation?
Since torpor is characterized by reduced metabolism and body temperature, a major question that immediately comes to mind is whether the profound reduction in cardiac output and oxygenation of the kidney and other tissues during hibernation results in renal ischemia during hibernation. The kidneys of nonhibernating animals in a healthy state receive about 20% of cardiac output, which mostly goes to the renal cortex, while the renal medulla receives only about 10% of the total renal perfusion (7), implying that the renal medulla is highly vulnerable to ischemia even at euthermic temperature. From a clinical point of view, ischemia occurs when there is an imbalance between blood oxygen demand and supply, which may result in cell death and tissue injury (15). To date, there are no good data to strongly suggest whether clinical renal ischemia occurs during torpor and, if it occurs, whether it may serve as a primary stimulus for arousal from torpor. Likely, the profound lowering of metabolism during torpor runs parallel with the reduced perfusion, which ensures a balance between blood oxygen demand and supply and thereby prevents ischemia during torpor (22).
Despite the debate whether or not torpor is associated with (renal) ischemia/reperfusion (I/R), the profound increase in metabolic rate upon rewarming after torpor likely results in high levels of oxidative stress and can potentially lead to cellular injury if not counterbalanced by protective mechanisms. Specific adaptations, consisting of upregulation of antioxidant enzymes and antiapoptotic pathways, limit oxidative stress and protect against cell death throughout hibernation (14). It should be noted that in contrast to hypothermic I/R, arousal from torpor may not be associated with a gross fluctuation in tissue oxygenation. In the brain of torpid Arctic ground squirrels, for example, there is a high arterial oxygen pressure and low expression of hypoxia inducible factor-alpha, a marker of tissue hypoxia (56), suggesting that hibernators may not show signs of ischemia during torpor. In conclusion, it remains to be revealed whether hibernators suffer renal ischemia from a clinical perspective during torpor and arousal.
H2S Lowers Metabolic Rate at High Concentrations, but Stimulates ATP Production at Low Concentrations
At high concentration of 80 ppm, for example, H2S can suppress metabolism through reversible inhibition of mitochondrial complex IV (cytochrome c oxidase) (6, 8). Thus, it is not surprising that H2S affects metabolism in animals. Inhibition of mitochondrial respiration was implicated in the induction of suspended animation in mice (6). Inhalation of gaseous H2S induces a hibernation-like state in nonhibernating mice by profoundly reducing the metabolic rate and core body temperature and protecting against cold renal IRI (8, 9, 11). Inhibition of oxidative phosphorylation by reversible antagonism at complex IV of the mitochondrial respiratory chain has been proposed to underlie the induction of such a hypometabolic state by H2S (8).
Whereas high dosages cause hypometabolism, low concentrations of H2S (0.1–1 μM) enhance electron transport in the mitochondria by acting as an electron donor at the level of coenzyme Q—an electron carrier between complexes I and III in the mitochondrial electron transport chain (ETC). Such electron donation by H2S stimulates cellular energy metabolism and ATP production through oxidative phosphorylation (36, 60). Recent studies also suggest that in the low-oxygen environment of ischemia/hypoxia, CBS and CSE enzymes translocate from the cytosol to the mitochondria to augment mitochondrial H2S production by 3-MST through electron donation and thus protecting liver and heart mitochondria against ischemic injury (32, 83). Perhaps the mitochondrial protection and preservation of ATP synthesis offered by H2S at low concentrations explain why hibernators are protected from cold ischemic injury. In conclusion, H2S at high concentration lowers metabolism, but at low concentration acts as an electron donor and preserves mitochondrial function.
H2S Protects and Prolongs Renal Graft Function During Cold Ischemia/Reperfusion and After Renal Transplantation
Since the discovery of the therapeutic potential of H2S, there has been more focus on the protective role of H2S in warm IRI in the kidney compared with cold IRI in renal transplantation. However, emerging evidence suggests that exogenous H2S administration protects renal cells against cold I/R-induced apoptosis, vasoconstriction, oxidative stress, and inflammation and thus increases the survival rate of the renal graft after transplantation. For example, following 18 h of cold (4°C) storage, NaHS treatment at 10 min before and after reperfusion has been shown to enhance renal blood flow (RBF) and increase renal graft function of porcine kidneys (41). Studies by our own group have demonstrated that addition of NaHS to UW solution during preservation of rat donor kidneys significantly mitigates cold IRI and improves post-transplant renal graft function and survival following both moderate and severe periods of cold (4°C) storage (53, 55). Thus, H2S protects and prolongs renal graft function during cold I/R and after renal transplantation.
H2S scavenges ROS and preserves renal function during cold I/R
It is well known that ROS is a chief mediator of cold IRI in human donor kidneys and therefore the use of H2S as an antioxidant has been suggested to mitigate oxidative stress during cold storage and reperfusion. Hosgood and Nicholson (41) observed a significant reduction of oxidative stress in porcine renal graft following treatment with NaHS before and during reperfusion. We also found that dopamine treatment preserved kidney function and integrity via maintenance (and upregulation) of renal CBS, CSE, and 3-MST and elevation of serum H2S, resulting in a marked decrease in the levels of renal malondialdehyde (MDA; a product of lipid peroxidation in the cell and a marker of oxidative stress) in a rat model closely matching clinical whole-body deep cooling and rewarming (12). Moreover, pharmacological inhibition of H2S production with amino-oxyacetic acid (AOAA) in dopamine-treated hypothermic rats strongly reduced renal expression of CBS, CSE, and 3-MST and serum H2S and significantly increased renal oxidative stress, resulting in severe renal injury (30).
This finding supports our previous study in which we showed that dopamine and serotonin doubled CBS expression via mammalian target of rapamycin, leading to increased endogenous H2S production and reduced ROS levels in cultured smooth muscle cells (81). This might also be the mechanism by which dopamine upregulates renal CSE and 3-MST under hypothermia/rewarming condition. Along the same train of thought, AOAA treatment also markedly reduced the serum H2S level (Fig. 3B) and creatinine clearance (Fig. 4B) in Syrian hamsters subjected to 5′-AMP-induced torpor-like state at 7°C body temperature (Fig. 3A) (29). However, a major caveat in the use of AOAA (and other endogenous H2S inhibitors) is its unspecificity to H2S-producing enzymes and therefore would require knockout/knockdown models to complement its use.
The antioxidant effect of H2S is also observed through therapeutic concentrations of the novel mitochondrial-targeting molecule, AP39 (49), which significantly reduced ROS production and preserved mitochondrial membrane integrity in our novel in vitro model of cold hypoxia/hypercapnia and warm reoxygenation (H/R) injury (54) as well as in other in vitro models of hypoxia/oxidative stress injury in renal epithelial cells and in a rat model of renal IRI (1), showing a direct mitochondrial effect of AP39. Mechanistically, H2S regulates ROS production by enhancing the transport of homocysteine, which is reduced to cysteine, serving as a substrate for the synthesis of glutathione (the most abundant antioxidant in cells) and inhibiting the activity of nicotinamide adenine dinucleotide phosphate oxidase (NADPH oxidase [NOX]; a major source of ROS in the kidney) (73). Although the effect of H2S on NOX expression and activity has not been described extensively in the context of renal transplantation, Han et al. (39) recently reported that H2S administration via NaHS increased total glutathione and other antioxidant levels and significantly decreased the expression of NOX4 and NOX1 (members of the NOX family), MDA production, and glutathione-to-oxidized glutathione ratio in a mouse model of warm renal IRI and in human vascular smooth muscle cells (62).
It is important to note that apart from functioning as an ROS scavenger, H2S also boosts the endogenous antioxidant defense system by activating other antioxidant enzymes, such as glutathione, catalase, superoxide dismutase, and nuclear factor erythroid 2-related factor 2 (Nrf-2), and by increasing levels of N-acetylcysteine (18, 75) (Fig. 5). However, considering the low concentration of endogenous H2S in blood and tissues, its ability to directly scavenge ROS and its direct interaction with oxidants cannot completely account for its antioxidant effect. Studies suggest that H2S interaction with nitric oxide and carbon monoxide (members of the gasotransmitter family) in the mitochondrial respiratory chain could activate their individual antioxidant activities via activation and opening of ATP-sensitive potassium (K+ ATP) channels (61, 66, 94) and therefore could serve as a backup mechanism of the antioxidant activities of H2S. In conclusion, H2S treatment offers protection against cold IRI partly through its potent antioxidant action, which also includes recruitment and activation of other endogenous antioxidant pathways.
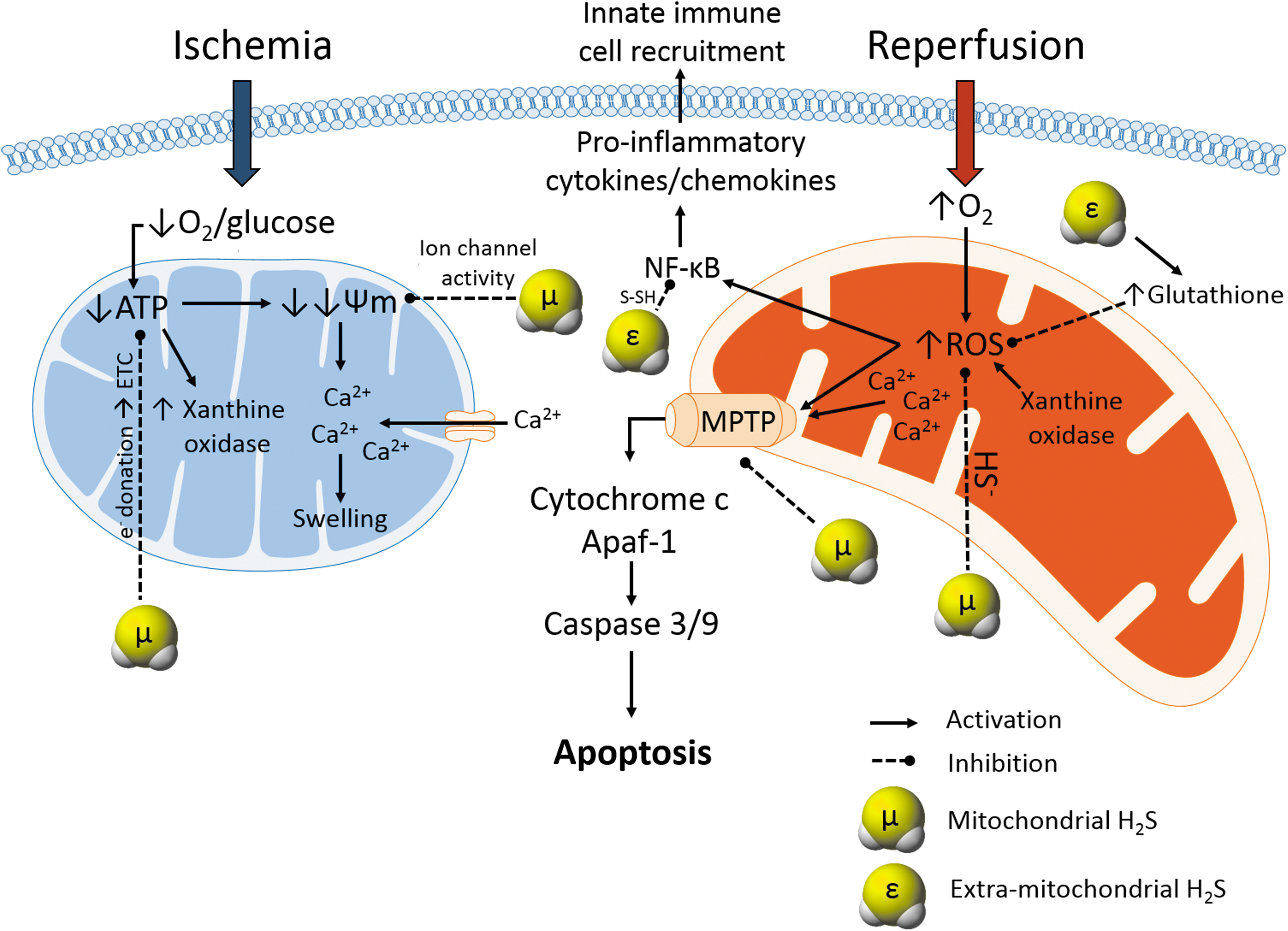
H2S maintains mitochondrial homeostasis and inhibits apoptosis during cold renal I/R
We have previously shown that H2S treatment protects donor kidneys against cold IRI at least partly by decreasing apoptosis in a rat model of renal transplantation. This was evidenced by a marked reduction in rates of renal graft apoptosis in conjunction with decreased expression of the proapoptotic gene, BH3-interacting domain death agonist (BID), and upregulation of the antiapoptotic gene, extracellular signal-regulated kinase-1 (ERK-1), compared with UW-treated renal grafts without H2S supplementation (53, 55). Similar to our studies, a recent study by Meng et al. demonstrated that the antiapoptotic role of H2S was associated with an increased expression of the antiapoptotic protein, Bcl-2, and decreased expression and activities of proapoptotic proteins (p38, Bax, and caspase-3) in both in vitro and in vivo models (58).
Considering both the increase in systemic H2S levels and the importance of mitochondrial protection for preservation of renal function during mammalian hibernation, it has been increasingly suggested that H2S may preserve mitochondrial integrity in a similar manner during cold IRI. We have recently produced novel and exciting evidence that supports this potential protective mechanism of H2S. In our study, we utilized AP39 to investigate whether targeting H2S release to mitochondria improves the potency of its protective effects during cold renal IRI. Using an in vitro model of H/R injury, we demonstrated that treatment of rat kidney cells with nanomolar concentrations of AP39 was at least 1000-fold more potent at preserving cellular viability following cold H/R injury compared with similar concentrations of the nonspecific H2S donor, GYY 4137 (54). Treatment of renal grafts with AP39 during prolonged cold storage also resulted in improved post-transplant graft function and survival, suggesting that targeting of H2S delivery to mitochondria could represent a viable and potent clinical strategy for minimizing the effect of IRI during transplantation (54).
Furthermore, AP39 stimulates mitochondrial respiration and generates mitochondrial sulfide and persulfide (49, 86), all of which act to maintain mitochondrial function during periods of hypoxia and thus make it an ideal candidate for organ preservation. Additionally, H2S (NaHS and AP39) inhibited Ca2+-mediated opening of mitochondrial permeability transition pore (MPTP) via a cyclophilin D-independent mechanism, preventing the release of proapoptotic factors from the mitochondria of hypertensive rat heart and also protecting rat myocardium against IRI (24, 44, 77). Hence, as hypothermia stimulates Ca2+ influx and mitochondrial Ca2+ accumulation (16), it is highly possible that H2S treatment of donor kidneys blocks Ca2+ influx and mitochondrial Ca2+ accumulation during cold storage by exerting a direct inhibitory effect on MPTP at a different site than cyclophilin D, thereby preventing the opening of MPTPs and preserving mitochondrial integrity (Fig. 5). Taken together, exogenous H2S appears to exert an antiapoptotic effect during cold renal IRI, which could be mediated by inhibitory actions against MPTP formation.
H2S induces renal vasodilation during cold I/R
Cold storage of donor organs induces vasoconstriction, which has been partially implicated in cold-induced organ injury during transplantation. Emerging reports indicate that H2S induces vasodilation and thereby mitigates injurious effects of cold-induced vasoconstriction. Xia et al. (89) first demonstrated that infusion of NaHS in intrarenal arteries in rats increases RBF and glomerular filtration rate (GFR) by increasing vasodilation in pre- and postglomerular arterioles and decreasing intrarenal vascular resistance, which would attenuate renal damage and improve oxygen and nutrient delivery to maintain functioning renal cells. This enhancement of RBF by H2S may result from activation and opening of K+ ATP channels and dilatation of blood vessels as this appears to be the underlying mechanism of protection by H2S in myocardial ischemia (93).
Following this initial demonstration of the effects of H2S on renal hemodynamics, Hosgood and Nicholson used a porcine model of renal transplantation to demonstrate that H2S treatment significantly increases RBF in donor kidneys 10 min before and after reperfusion compared with the control group without H2S supplementation (41). We have also observed that NaHS administration improves RBF and reduces the renal resistive index in a representative porcine model of donation after cardiac death (DCD) kidney transplantation (2). In addition, we subsequently developed a rat model of DCD renal transplantation to show that treatment of DCD kidneys with H2S during both warm and cold IRIs results in decreased post-transplant graft injury and improved subsequent graft function and survival (37). Thus, by improving resistive index and RBF, H2S may improve perfusion parameters, which then may translate to improved renal function upon transplantation (Fig. 6).
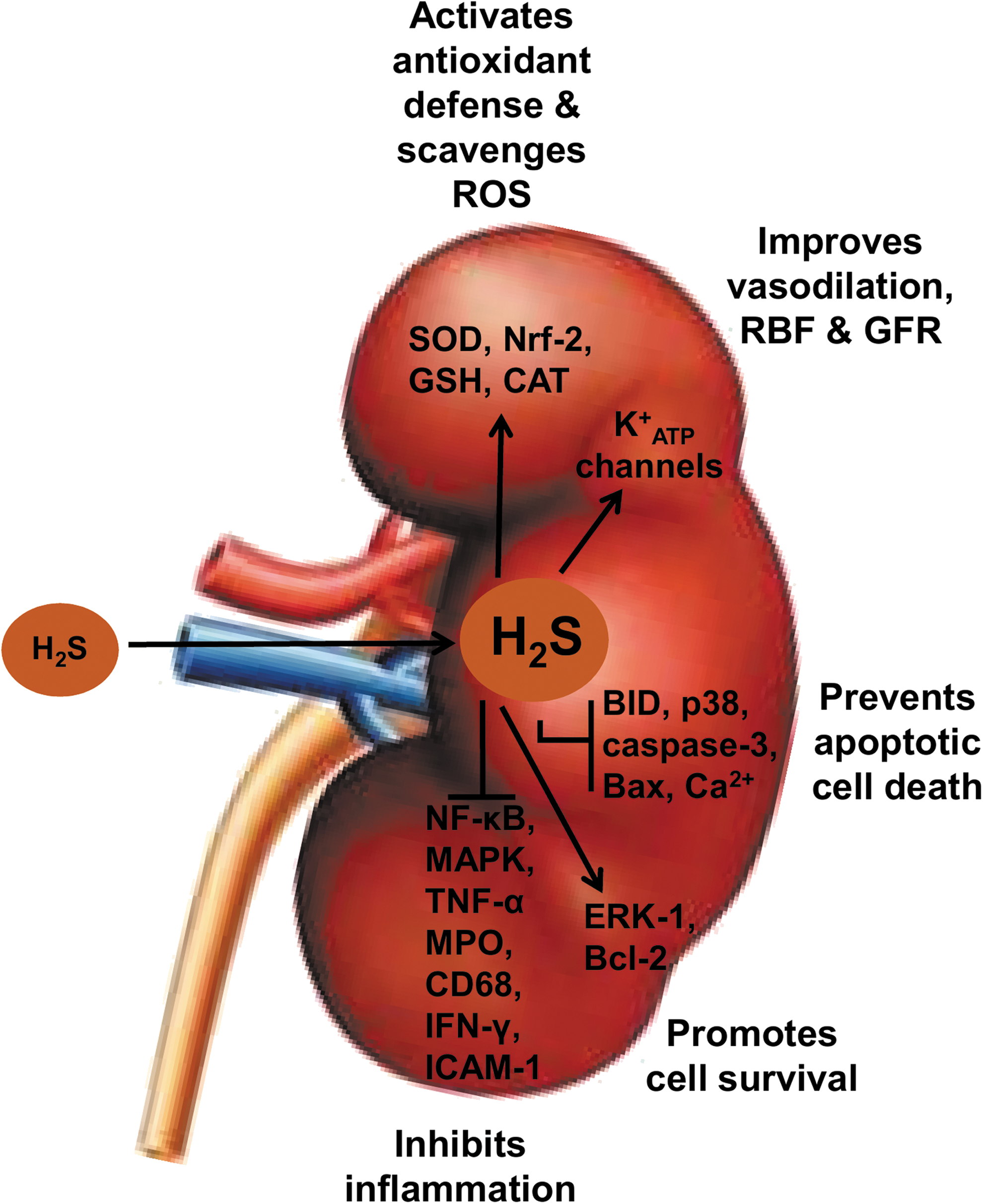
It is important to point out that cyclosporine, a widely used immunosuppressive drug after renal transplantation, induces nephrotoxicity, which begins with renal and systemic vasoconstriction, leading to reduced GFR and ultimately interstitial fibrosis and tubular atrophy (72). Interestingly, a recent study has shown that H2S infusion before and after reperfusion of cold-stored porcine kidneys reverses cyclosporine-induced vasoconstriction and glomerular atrophy by inducing vasodilation and increasing RBF, suggesting an additional protective role of H2S-induced vasodilation during the transplant procedure (50). Thus, the vasodilatory effects of H2S appear to reduce renal resistance and increase RBF in donor kidneys following cold storage, resulting in improved reperfusion and function of renal grafts in the acute post-transplant period.
H2S suppresses renal inflammation induced by cold I/R
Apart from antioxidant, antiapoptotic, and vasodilatory effects, the therapeutic properties of H2S in renal transplantation also include an anti-inflammatory component. For example, we have previously demonstrated that H2S treatment of rat renal graft during cold storage significantly decreased infiltration of myeloperoxidase (MPO)-positive neutrophils and CD68-positive macrophages and expression of the proinflammatory cytokines interferon-gamma (IFN-γ) and intercellular adhesion molecule-1 (ICAM-1) in the acute post-transplant period (55) (Fig. 5). Our finding was confirmed by a recent study in which H2S administration suppressed influx of MPO-positive neutrophils in the kidneys of rats subjected to warm IRI (1).
While the anti-inflammatory property of H2S in renal transplantation remains to be revealed, other studies have reported more potent anti-inflammatory effects and mechanisms of H2S in various disease models (58). The slow-releasing H2S donor, GYY4137, inhibits nuclear factor-kappa B (NF-κB) and mitogen-activated protein kinases (p38, ERK1/2, and JNK1/2) (88), two key mediators of inflammation. Such anti-inflammatory effect of H2S was partially involved in myocardial protection through increased plasma H2S concentration and CSE activity in a rat model of myocardial IRI (58). H2S inhibits the expression of leukocyte adhesion molecules, thereby preventing leukocyte rolling and attachment to the endothelium (91). This inhibition was associated with activation of K+ ATP channels by H2S as K+ ATP antagonist reversed the leukocyte inhibitory effect of H2S and increased leukocyte adherence (91). These mechanisms underlying the anti-inflammatory action of H2S may thus partly account for renal protection against cold IRI in transplantation.
Protective Role of H2S Against Cold IRI During Transplantation of Other Solid Organs
Exogenous H2S treatment also proved protective against cold IRI in transplantation of various solid organs other than the kidney. In a rabbit model of lung transplantation, studies by George et al. showed that inhalation of gaseous H2S at 150 ppm for 2 h before 18 h of cold storage significantly decreased the ROS level, improved oxygenation, lowered pulmonary artery pressure, and prevented cold ischemic injury, thus leading to improved graft function during reperfusion compared with the control group (33, 34). This suggests that H2S is a potent scavenger of free radicals as discussed above. In a different experimental setting of cold IRI during lung transplantation, 3 h of cold storage, followed by NaHS treatment 15 min before reperfusion, significantly increased lung tissue expression of CSE and H2S levels and reduced membrane lipid oxidation levels, resulting in improved pulmonary function and lung protection following transplantation compared with control rats (87). Administration of propargylglycine (PPG; CSE inhibitor) 15 min before reperfusion markedly decreased lung CSE protein and H2S levels and increased lipid peroxidation and MPO activity, culminating in overproduction of ROS and pulmonary injury after transplantation (87). This suggests that reduced production and malfunction of endogenous H2S contribute to cold IRI in the lung.
These findings also suggest that both endogenous and exogenous H2S treatments protect against cold IRI partly via their antioxidant properties. Although the mechanism by which PPG downregulates CSE expression and activity within such a short time is unknown, it is possible that lung CSE-producing cells are more sensitive to PPG than those of other organs, which might have resulted in death of CSE-producing cells within this short period. Moreover, the dose of PPG (37.5 mg/kg) used in the study might be higher than that used in other studies. This, therefore, needs further investigation. In a rat model of DCD liver transplantation, Na2S and gaseous H2S treatments increased the levels and activities of liver antioxidants and strongly reduced the levels of lactate dehydrogenase, MDA, and other liver damage markers during cold I/R and hence improved liver microcirculation, morphology, and function (3, 4). Thus, H2S-mediated cytoprotection against tissue IRI observed during renal transplantation is translatable to other tissues and should be further investigated to improve the outcomes of all solid organ transplantations.
Conclusion and Future Perspectives
To our knowledge, there are no therapeutic strategies that can successfully mitigate IRI associated with organ transplantation. Therefore, the therapeutic use of exogenous H2S added to standard preservation solutions or inducing endogenous H2S seems a promising approach to prolong renal graft preservation and mitigate unavoidable cold IRI during transplantation of kidney and other solid organs (Fig. 7). Targeting exogenous H2S administration to mitochondria increases the potency of its protective effects during IRI, implicating mitochondria as a primary site of action for H2S-mediated renoprotection. Exogenous H2S could potentially control mitochondrial mechanisms of IRI pathogenesis directly by driving ATP synthesis via donation of electrons to the ETC (36, 60), modulation of mitochondrial Ca2+ channels to prevent Ca2+ influx (82, 85), direct inhibition of mitochondrial ROS (20, 63), and prevention of MPTP formation (77) or indirectly via inhibition of proinflammatory NF-κB signaling (71) and upregulation of cellular antioxidant enzymes such as glutathione (46, 90).
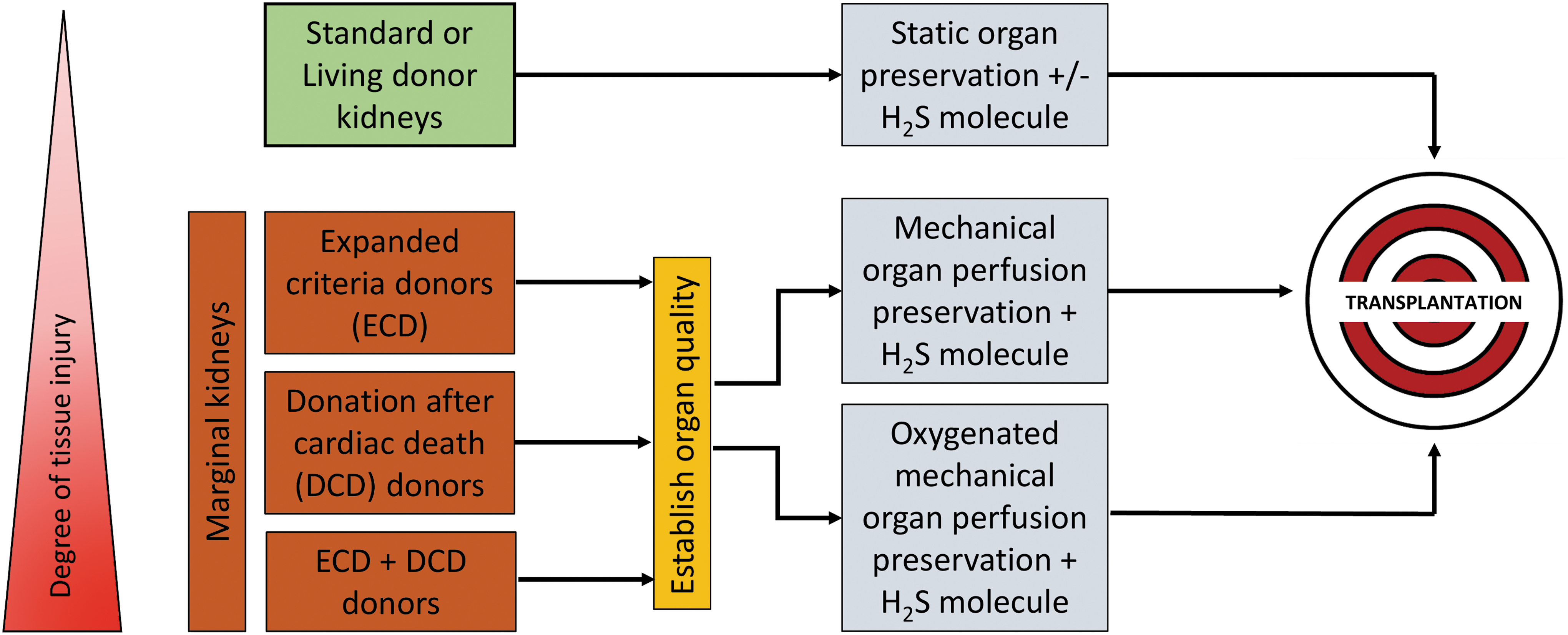
Although extensive investigation in defining molecular mechanisms behind IRI and the use of various therapeutic strategies have been proposed, ongoing clinical studies, including those utilizing rPSGL-Ig (a p-selectin inhibitor) and mitoQ (an antioxidant), have provided some satisfying results in preventing renal IRI (25). In all cases, more clinical trials with improved standardization of protocols are required to contribute to important advances in clinical therapy (65). Apart from involvement of H2S in transplantation, additional lessons can be learned from dissecting mechanisms by which nature protects organs of hibernating animals from the damaging effects of cold exposure and reperfusion. Understanding such mechanisms will be beneficial for the development of more potent forms of the current therapeutic or novel therapeutic agents. In addition, future investigations should be aimed at developing new drugs or a combination of current therapeutic agents that could afford a multitarget approach to minimize IRI during transplantation. In this way, we may be able to take advantage of natural cytoprotective mechanisms utilized by hibernating mammals, such as H2S-mediated mitochondrial preservation, to ultimately improve clinical outcomes and quality of life for renal transplant recipients.