
Abstract
Aims:
Pyroptotic cells are characterized by plasma swelling, membrane blebbing, and disintegration of the cell membrane mediated by spectrin-based membrane skeleton and intercellular competitive tension activities. The spectrin-based membrane skeleton is involved in membrane organization through the regulation of intercellular tension. Using genetically encoded tension sensors to attain noninvasive force measurements in structural proteins, we investigated how cytoskeletal structural tension influences changes in plasma morphology during pyroptosis and the regulatory mechanism of cytoskeletal structural tension that underpins pyroptosis.
Results:
The results indicate that increasing spectrin tension is caused by osmotic swelling. Hightened tension of spectrin was closely associated with the shrink tension transmitted synergistically by microfilaments (MFs) and microtubules (MTs). However, the increment of spectrin tension in pyroptotic cells was controlled antagonistically by MF and MT forces. Different from MF tension, outward MT forces participated in the formation of membrane blebs. Spectrin tension caused by inward MF forces resisted pyroptosis swelling. Stabilization of MF and MT structure had little influence on intracellular tension and pyroptosis deformation. Pyroptosis-induced cytoskeletal structural tension was highly dependent on calcium signaling and reactive oxygen species generation. Blocking of membrane pores, nonselective ion flux, or elimination of caspase-1 cleavage resulted in the remission of structural forces associated with pyroptosis failure.
Innovation and Conclusions:
The data suggest that subcellular tension, in terms of magnitude and vector, is integral to pyroptosis through the mediation of swelling and blebbing and the elimination of structural tension, especially MT forces, may result in pyroptosis inhibition.
Introduction
Pyroptosis is inflammatory necroptosis with pore formation in the plasma membrane, followed by cell swelling, massive membrane blebbing, and disruption. Pyroptosis can alter cell shapes locally and globally through plasma membrane tension (1, 10, 49). This feature of pyroptosis requires cytoskeleton remodeling and forward stretching of the plasma membrane to resist cytoskeletal tractive forces. In this way, pyroptotic cells promote tension-dependent changes in the plasma membrane (4). Spectrin acts as a scaffold to crosslink transmembrane proteins, actin, and tubulin cytoskeleton that sense plasma membrane tension. It also can integrate these joint inputs into the dynamics of the cytoskeleton and mediate tension-dependent changes in cell shape and cytoskeletal structure (8, 19, 25, 40). The junction protein ankyrin provides a binding site for spectrin and tubulin (3, 29, 30), while protein 4.1 facilitates spectrin actin binding (34, 40, 55). Membrane tension and cytoskeletal activity are tightly connected; thus, they adjust intercellular tension transmission and regulate cytoskeleton deformation in complex physicochemical environments (14).
This study establishes a novel method to analyze cytoskeleton-dependent tension vectors. The subcellular tensions, their magnitude, and direction influence cell swelling and blebbing during cell pyroptosis. Outward microtubule (MT) forces contribute to bleb formation, while spectrin tension elicited by inward microfilament strain modifies membrane swelling during pyroptosis. The remission of structural tensions, especially MT forces, is closely associated with pyroptosis failure. Similar to chemical and electrical cell activity, intracellular cytoskeletal structural tensions are essential contributors to metabolic life in living cells.
The spectrin-based membrane skeleton balances the cytoskeletal structural tension between the plasma membrane and cytoskeleton, thus determining the shape of cells (15, 17, 48, 57). Although little is known about how the spectrin–actin–tubulin network is formed and connects to the tension transmission to cause swells and blebs during pyroptosis, some differences between membrane skeletons have been recognized. Cortical microtubules (MTs) line the shafts of blebs that slide via cytoplasmic dynein and kinesin power sources to expand the blebs (26). F-actin, present throughout the blebs, forms myosin assemblies to stabilize the blebs (2, 41, 42). The tension from myosin contraction can be transmitted to spectrin through the actin cytoskeleton, while the tension activated by dynein and kinesin can be transmitted to spectrin through the tubulin cytoskeleton (7, 28, 32, 52). The cells are also regulated by extracellular forces, such as osmotic pressure, to prompt inward and outward tension, harmonize intracellular tension, and accommodate shape and volume changes (5, 13). Moreover, inter- and intracellular tensions coparticipate in the regulation of swelling and blebbing in pyroptosis (39, 51).
Caspase-1-dependent insertion of a pore into the plasma membrane results in the loss of membrane integrity in pyroptosis (27). The membrane pores dissipate the balance of cellular ionic gradients, leading to water influx, osmotic pressure alterations, and perturbation of cytoskeleton tension, thus promoting swelling and blebbing in pyroptotic cells (10). It has been demonstrated that osmotic swelling and blebbing are prevented by membrane pore blocker osmoprotectants with molecular diameters greater than the functional diameter of the pores, or inhibition of nonselective ion fluxes by the cytoprotective agent glycine. Furthermore, calcium signals from the activation of calcium channels and reactive oxygen species (ROS) generated through inflammatory responses have been recognized to participate in pyroptosis (10, 33, 49, 50, 53, 58). The formation of the membrane pores reportedly requires the rearrangement of the host cellular cytoskeleton. However, whether or not pyroptosis is associated with intracellular structure tension remains unclear.
Due to the absence of a method to measure forces across structural proteins at the subcellular level, it has been difficult to elucidate how cytoskeleton tension influences changes in plasma morphology and function during pyroptosis. We previously developed a genetically encoded fluorescence or förster resonance energy transfer (FRET)-based tension sensor named spectrin–M-cpstFRET (ScpS; cpstFRET, circularly permutated sstFRET) (22), actin-cpstFRET-actin (AcpA) (23), and αtubulin -cpstFRET-βtubulin (TcpT) to use in cell pyroptosis research. These noninvasive and biocompatible tension sensors were hung on the cytoskeleton, allowing the transformation of the tensile forces into optical signals during the expansion or contraction of the cytoskeletal networks (12, 20, 22, 43, 44). Here we clarify and extend these findings and provide mechanistic insights into how subcellular tensions as a vector parameter (magnitude and direction) are regulated during cellular swelling and blebbing, and how this structure tension affects spectrin function during cell pyroptosis.
Results
Cytoskeleton homeostasis is sensitive to spectrin tension during pyroptosis or apoptosis
Pyroptosis and apoptosis are two common forms of programmed cell death, involving rearrangement of the cytoskeleton and alteration of cell volume. To compare differences between pyroptosis with apoptosis, we first analyzed intracellular construction characteristics of both. Using confocal microscopy to examine the cytoskeleton, we assessed actin and tubulin with fluorescein isothiocyanate (FITC) staining and labeled spectrin with tetraethyl rhodamine isothiocyanate (TRITC) in MCF-7 cells. Interestingly, we found that there were no differences between the cytoskeleton deformations in the control, pyroptotic, and apoptotic cells (Fig. 1A), although pyroptotic cells were swollen and apoptotic cells were shrunken (Fig. 1B, C). These morphological changes should be accompanied by cytoskeleton reorganization. Spectrin participates in cytoskeleton reorganization and membrane tension changes via protein–protein interactions and intracellular tension transmission. To test this, we analyzed the contractile tension in spectrin and its effect on morphogenesis using a FRET-based tension sensor, which reports resonant angle twisting toward a perpendicular configuration by a reduction in energy transfer (45). A FRET module was incorporated within the spectrin backbone (Supplementary Fig. S1A). On acceptor bleaching (AB), there was a marked decrease in the acceptor fluorescence (enhanced yellow fluorescent protein [eYFP]), which coincided with an increase in the donor fluorescence (enhanced cyan fluorescent protein [eCFP]) because of an inability of the acceptor to accept energy from the donor after bleaching (Supplementary Fig. S1B, ScpS FRET AB = 22.35%). Fluorescence recovery after photobleaching measurements indicated that the recovery rates for spectrin, microfilaments (MFs), and MTs were 45.43%, 71.35%, and 12.15%, respectively, after 500 s (Supplementary Fig. S1C). For their respective controls, Scp, Acp, and Tcp, the FRET index was very high compared with the tension probes (Supplementary Fig. S2). When expressed in pyroptotic cells, ScpS-generated FRET signals were conspicuously higher than those from control cells, as swells and blebs are sensitive to tension. This result implies that pyroptotic cells exhibited higher spectrin tension due to blebbing and swelling for tension regulation (Fig. 1B, D). Consistent with this, the spectrin tension in apoptotic cells was conspicuously higher than in controls and pyroptotic cells (Fig. 1D), with increased contractility at cytoskeleton polymerization sites, heightened spectrin tension, and significantly reduced cell surface area at the plasma membrane (Fig. 1C). These results indicate that cytoskeleton homeostasis is sensitive to spectrin tension during pyroptosis or apoptosis.
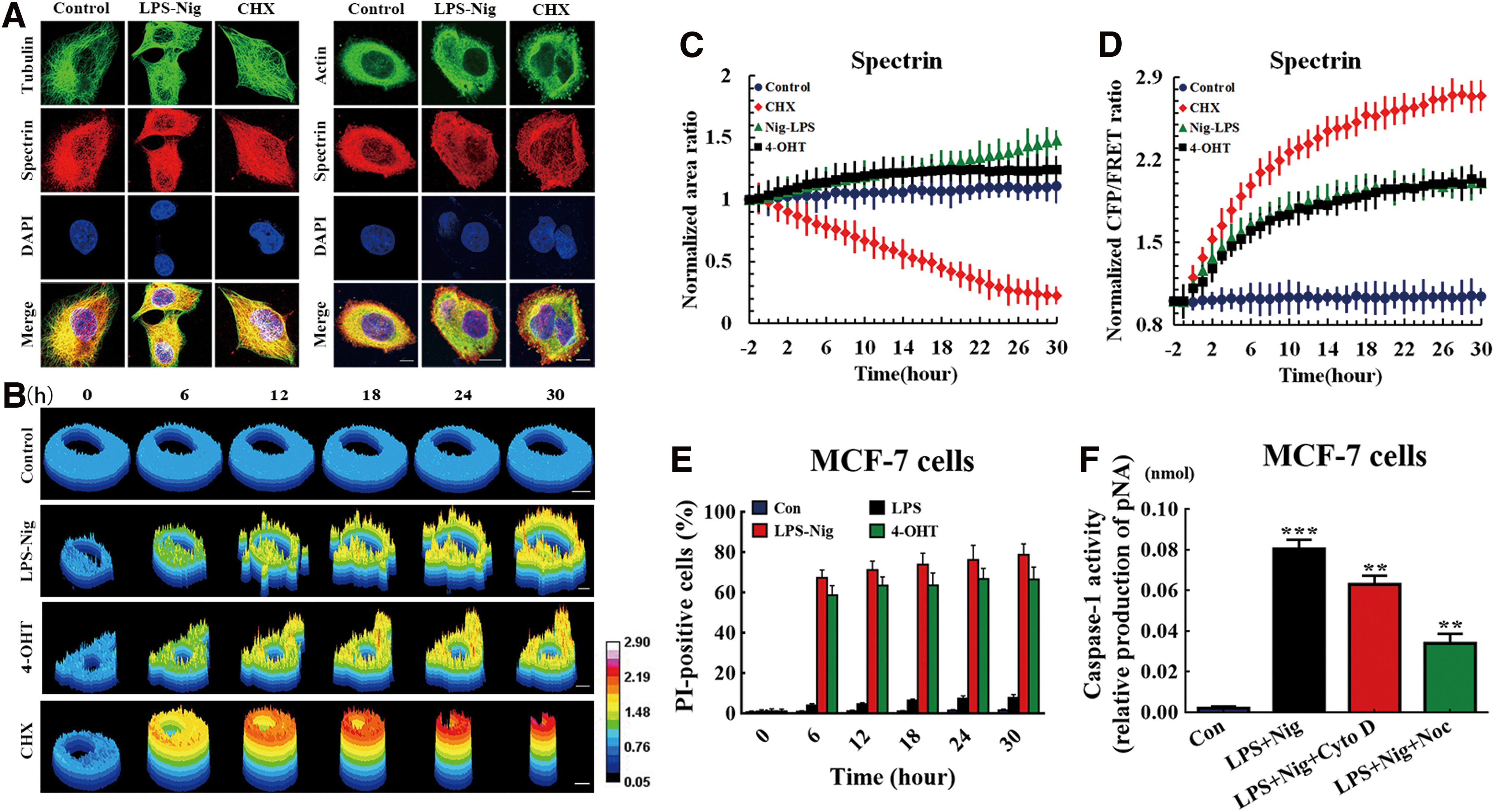
Spectrin tension participates in cell swelling and shape alterations when under extracellular stress
Cells undergo dramatic shape and volume changes and membrane skeleton interactions, which are closely related with membrane tension dynamics. With cytoskeletal structural tension and area in a feedback loop, high tensions favor recruitment of membrane to the surface in cell swelling situations. The hypoosmotic pressure that causes outward tension, through the pulling of cytoskeletal filaments, is associated with cellular swelling and enlarged volume. To measure the forces in spectrin during cell swelling, we exposed MCF-7 cells to hypoosmotic stress by switching the bath (isotonic HEPES buffer) from saline to hypoosmotic (250, 200, 100, and 0 mOsm/kg). It may be surprising to learn that many nucleated cells do not lyse under these conditions. We found that spectrin tension remained stable under isotonic conditions (300 mOsm/kg; Fig. 2A). The cells reacted to the hypoosmotic challenge by swelling followed by an actual decrease in cell volume, the latter process being known as regulatory volume decrease (RVD) (31, 46) (Fig. 2C). After the change in cellular volume was detected, an immediate homeostatic response followed with a sustained increase of spectrin tension observed (Fig. 2B). These results imply that increased spectrin tension is involved in cell swelling and even the RVD phenomenon. The spectrin tension might be inward and antagonize outward force under hypoosmotic pressure.

MF and MT—forces involved in the regulation of spectrin tension in response to osmotic pressure
Spectrin can combine with MFs and MTs and then transmit their tension to the membrane via adapter proteins ankyrin and 4.1. We explored the role of MF and MT tension in spectrin responses to inward or outward forces that originated from osmotic pressure. Figure 3A shows the spatiotemporal change of spectrin tension after hypoosmotic stimulation (15 min). After hypoosmotic stimulation, spectrin tension, including MF and MT forces, immediately increased as cells simultaneously swelled (Fig. 3A, B). To explore the role of MF or MT forces in cell swelling, depolymerizing agents of MF and MT, cytochalasin D (cyto D) and nocodazole (Noc), respectively, were used. The depolymerization of MFs by cyto D induced a relative decrease in spectrin and MT tensions, compared to the hypoosmotic treatment. MT depolymerization by Noc similarly lessened spectrin and MF tensions. Cotreatment of cyto D and Noc reversed the spectrin tension to the isotonic level (Fig. 3A). These data suggest that MF and MT forces can act synergistically in spectrin tension processes, by resisting the outward stress in hypoosmotic conditions. Then, in response to a high concentration of mannitol solution (555 mOsm/kg), we examined the hyperosmotic effect, which induced a membrane-inward force and manifested as decreases in spectrin, MT, and MF tensions and cell shrinking (Fig. 3C, D). The depolymerization of MFs by cyto D led to a reduction in spectrin tension but an increase in MT force, compared to the hyperosmotic treatment. MT depolymerization by Noc enhanced both spectrin tension and MF tension in hyperosmotic conditions (Fig. 3D). The results indicate that MF and MT forces acted antagonistically on spectrin tension under hyperosmotic stimuli. However, MTs but not MFs contributed to weakening spectrin tension, which resisted inward stress caused by hyperosmotic pressure. To summarize, spectrin tension could be adjusted synergistically or antagonistically by MF and MT tension, against inward or outward stresses caused by changes in ambient osmotic pressure.

Spectrin tension induced by MF and MT forces results in swelling and blebbing of pyroptotic cells
Cell shape can be altered by swelling and blebbing during pyroptosis. To explore the mechanical mechanisms that underpin pyroptosis, we constructed a pyroptosis model that could monitor the kinetics of cytoskeletal structural tension under different stimuli. MCF-7 cells underwent pyroptosis on lipopolysaccharides (LPS) + nigericin (LPS + Nig) or 4-hydroxytamoxifen (4-OHT) treatment, in accordance with previous reports (31, 46). Pyroptotic cells exhibited swelling and produced multiple bubble-like protrusions, verified by active caspase-1- and propidium iodide (PI) staining (Figs. 1E, F, and 4K) and increased cell volumes (Fig. 4J). Meanwhile, a temporal increase in spectrin tension, including MF and MT forces, was observed (Fig. 4A). On MF depolymerization, spectrin tension decreased to control group levels (Fig. 4A), with the obvious production of large blebs (Fig. 4A, J, and L). MT depolymerization resulted in a sustained rise in spectrin tension (Fig. 4A), which was accompanied by the disappearance of blebs (Fig. 4J, L), distinctly different from the effects induced by MF depolymerization. Interestingly, combined MF and MT depolymerization not only decreased the spectrin tension but also suppressed cell swelling and blebbing (Fig. 4A, J, and L). Combined, these results indicate that MT stress was the dominant force in cellular blebbing, which in turn indicates that stretching forces presenting on the cell surface are derived from MT tension. In the pyroptosis model, growing spectrin tension was accompanied by an increase in cellular volume, to the extent that the average volume increase was recorded as 1.32-fold the original volume. MF depolymerization was also accompanied by a transient increase in cell volume (Fig. 4J). However, no detectable swelling was observed on MT depolymerization or MF and MT combined depolymerization. Furthermore, MF (Fig. 4C) and MT (Fig. 4E) forces in the pyroptosis model were developed temporally. MF and MT depolymerization can lead to increases in MT and MF tension, respectively (Fig. 4C, E), suggesting that MT and MF tensions interact antagonistically in pyroptosis.
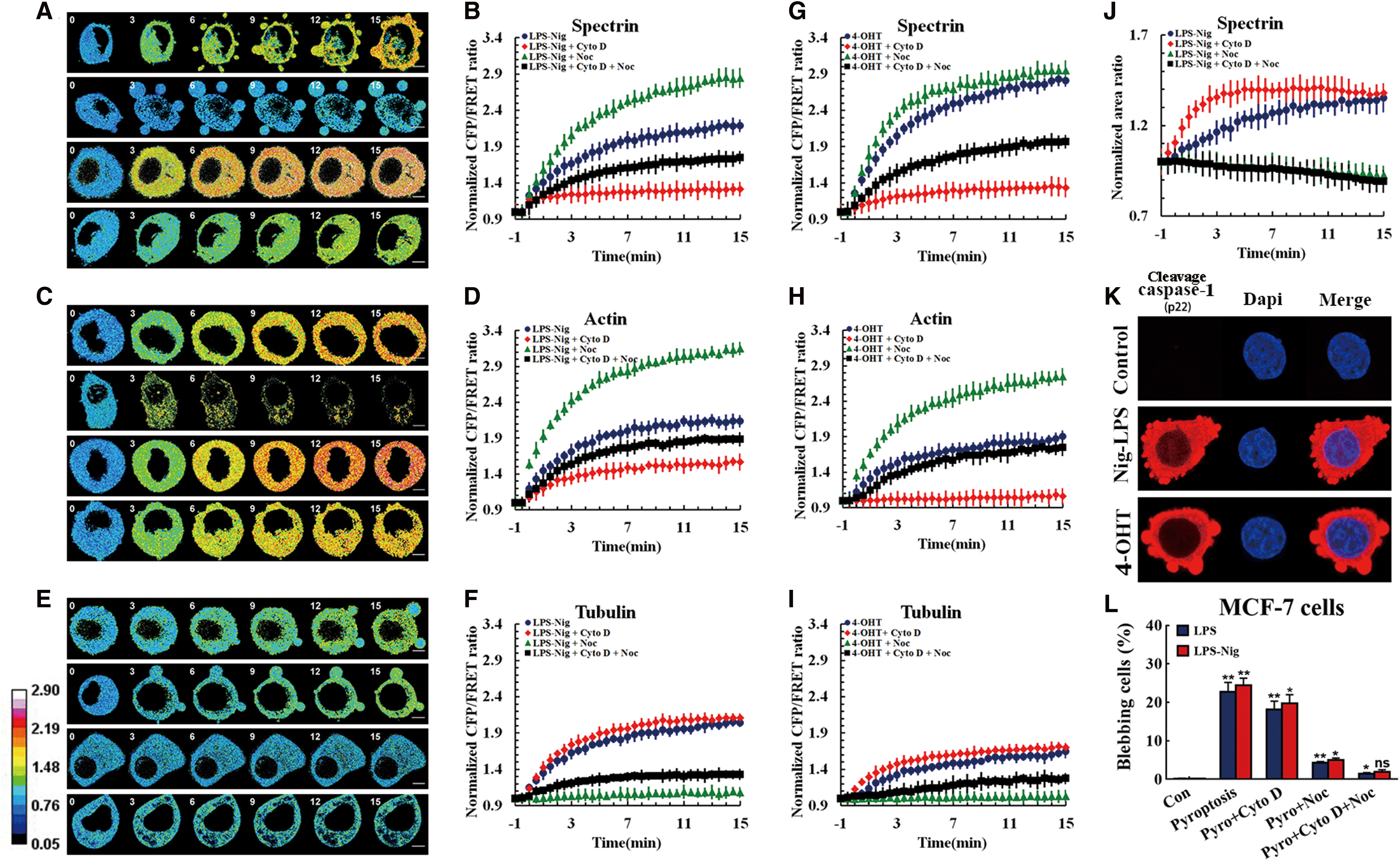
Meanwhile, the dynamics of spectrin, MF, and MT tensions and their interactions, as elicited by 4-OHT stimuli, are similar to those elicited by LPS + Nig treatment (Fig. 4G–I), suggesting that both treatments share similar tension mechanisms. These results imply that MT forces, but not MF forces, induce the swelling and blebbing phenomena observed in pyroptotic cells, and spectrin tensions elicited by MF forces resist changes to cell shape as a stress response.
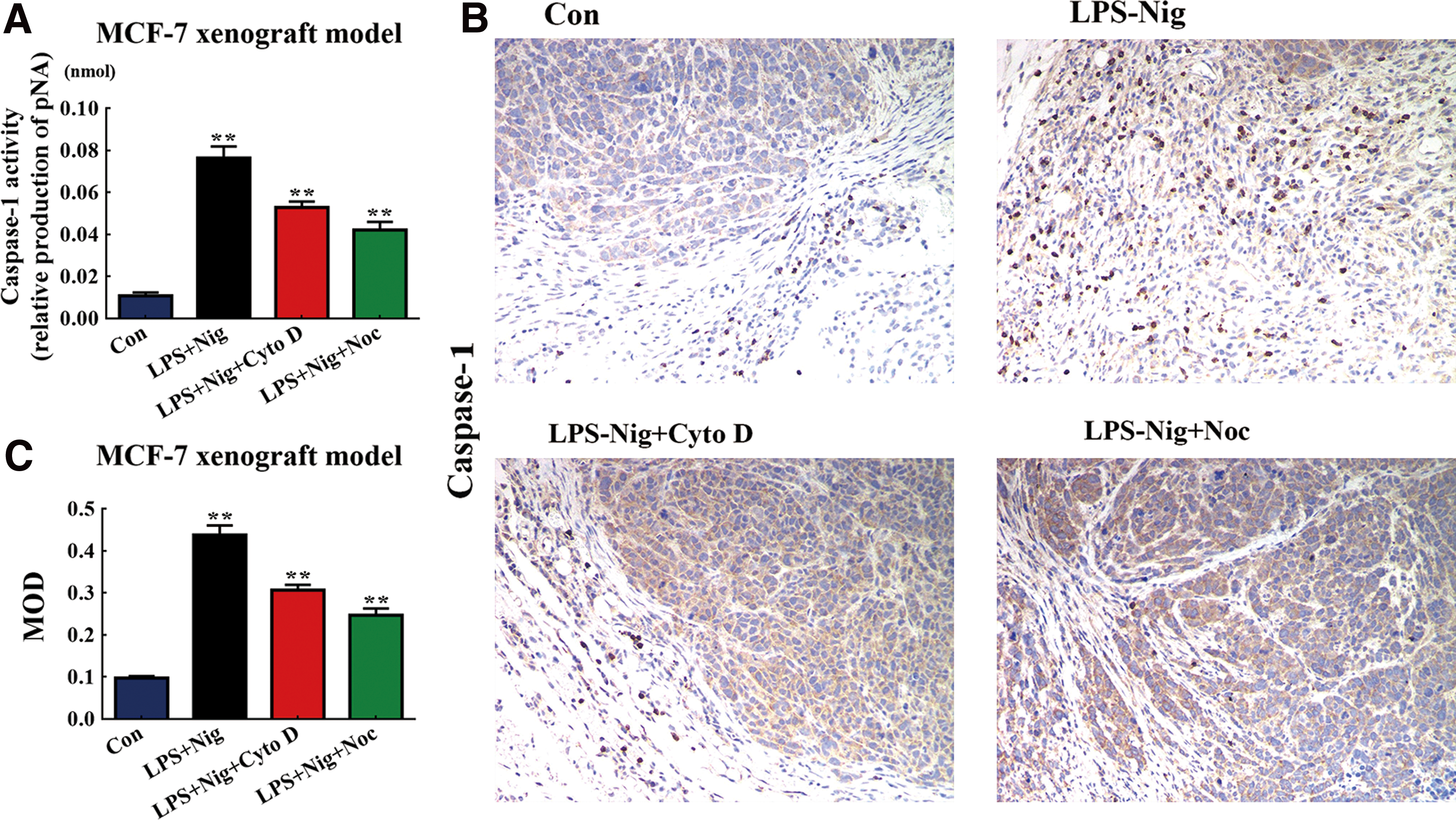
Depolymerization of MF or MT contributed to varying degrees of pyroptosis program inhibition in vivo
Cytoskeleton depolymerization was found to influence the regulation of blebs and swells during pyroptosis. We examined the role of MFs and MTs in a pyroptosis model. LPS-Nig increases the caspase-1 activity in MCF-7 cells, indicating a significant increase in pyroptosis rates. The depolymerization of MFs caused pyroptosis rates to decrease, while the depolymerization of MTs lowered the rate of pyroptosis (Fig. 5A, B). To understand the cytoskeleton function in pyroptosis, we explored the influence of MF and MT depolymerization in vivo pyroptosis experiments through immunohistochemical (IHC) staining. With intraperitoneal injection of LPS-Nig, significant activation of caspase-1 was detected by IHC in the pyroptosis mammary glands (Supplementary Fig. S3A) and the MCF-7 xenograft model (Fig. 5A). Interestingly, MF and MT depolymerization resulted in a meaningful reduction in caspase-1 activity compared with the in vivo pyroptosis model in mammary glands (Supplementary Fig. S3B, C) and in the MCF-7 xenograft model (Fig. 5B, C), and the role of MT was superior to that of MF (Fig. 5B, C). These results suggest that MF and MT depolymerization reduces pyroptosis rates and that depolymerization of MT is more evident than MF's.
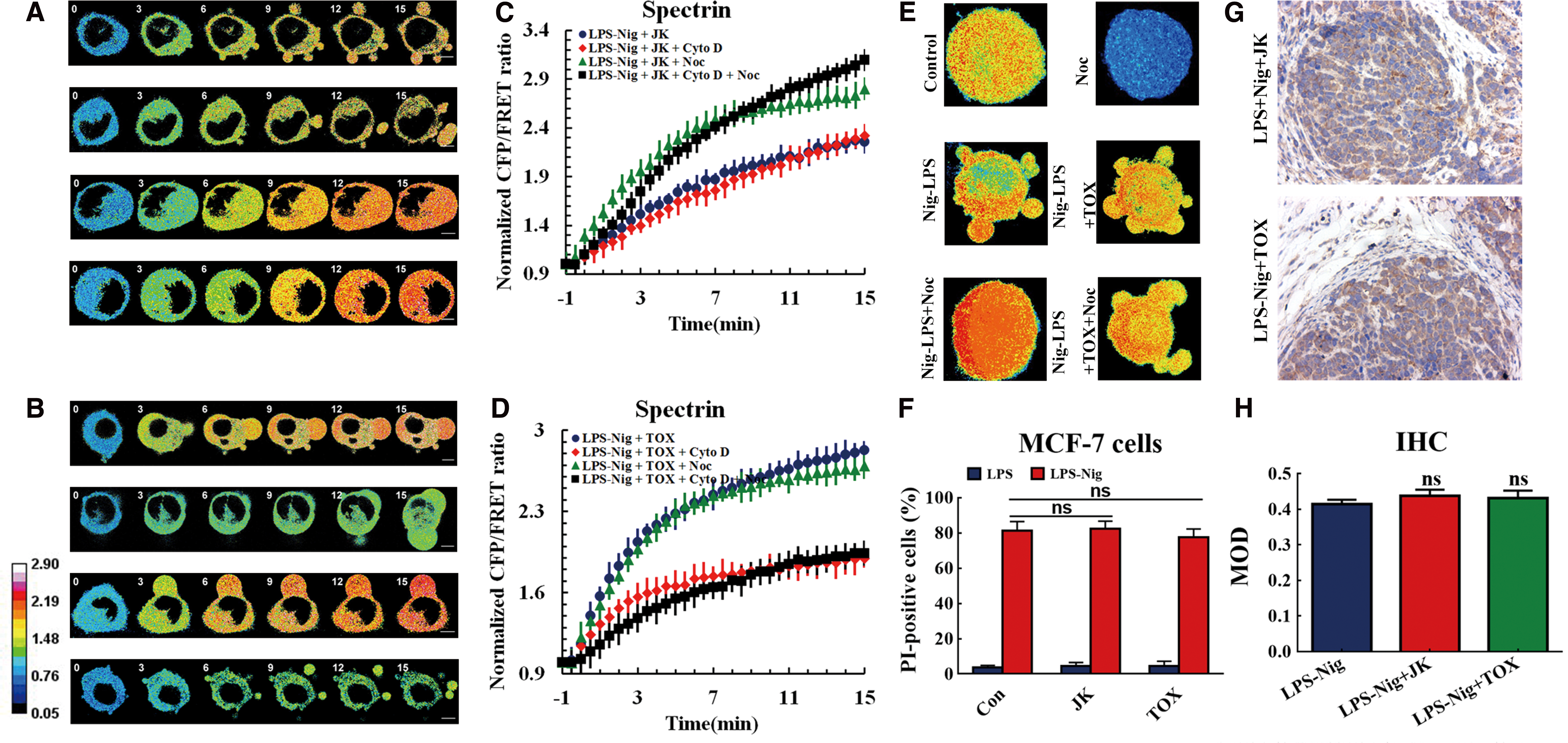
Cytoskeleton structure has minimal influence on intracellular cytoskeletal structural tensions in pyroptosis
Changes in cell shape are thought to be closely associated with cytoskeleton rearrangement. Cytoskeleton depolymerization was found to regulate blebs and swells in pyroptosis processes (10). To better understand the role of the cytoskeleton in pyroptosis, we assessed the effect of MF and MT structure stabilization on intracellular tensions in a pyroptosis model. MF stabilizer jasplakinolide (JK) and MT stabilizer taxol (paclitaxel [TOX]) were used in the mechanical study (45, 59). Neither JK nor TOX treatment had any effect on spectrin tension or cell shape in the pyroptosis model (Fig. 6A, B, and E). Cyto D or Noc treatments also did not alter the spectrin force or cell shape when applied in response to JK or TOX administration. As expected, cyto D and Noc could modify spectrin tension in a pyroptosis model with the treatment of MF and MT stabilizers, respectively (Fig. 6C, D). The pyroptosis rate after JK or TOX treatment was not significantly different to the pyroptosis model (Fig. 6E, F). Moreover, IHC staining was also used to confirm no significant difference between control and JK or TOX treatment in the in vivo MCF-7 xenograft model (Fig. 6G, H). These results indicate that intracellular cytoskeletal structural tension plays a more vital role than cytoskeleton structure in cell swelling and blebbing in pyroptosis. However, intracellular mechanical activities are dependent strictly on the integrity of the cytoskeleton structure.
The essential role of calcium channels in intracellular tension during pyroptosis
The enhancement of spectrin tension, combined with MF and MT forces, regulates swelling and blebbing in pyroptotic cells. Ion signals also play a role in pyroptosis due to imbalances in cellular ions on both sides of the plasma membrane. To further elucidate the influence of calcium channel force/voltage transfer, we examined the influence of ion channels on spectrin tension in our pyroptosis model. The calcium channel activator pregnenolone sulfate (PREGS) can aggravate spectrin tension, including MF and MT forces in pyroptosis models (Fig. 7A). The depolymerization of MFs led to a decrease in spectrin tension, but MT depolymerization resulted in an increase in spectrin tension. After PREGS treatment, MF and MT forces induced a relative increase in spectrin tension antagonistically (Fig. 7C). Moreover, the activator increased MF and MT forces and was associated with a higher pyroptosis rate than the pyroptosis model without activation (Fig. 7E). These results imply that calcium channels may be strongly involved in the regulation of cytoskeleton tension and cell pyroptosis, and that increases in intracellular forces are closely associated with the development of pyroptosis.
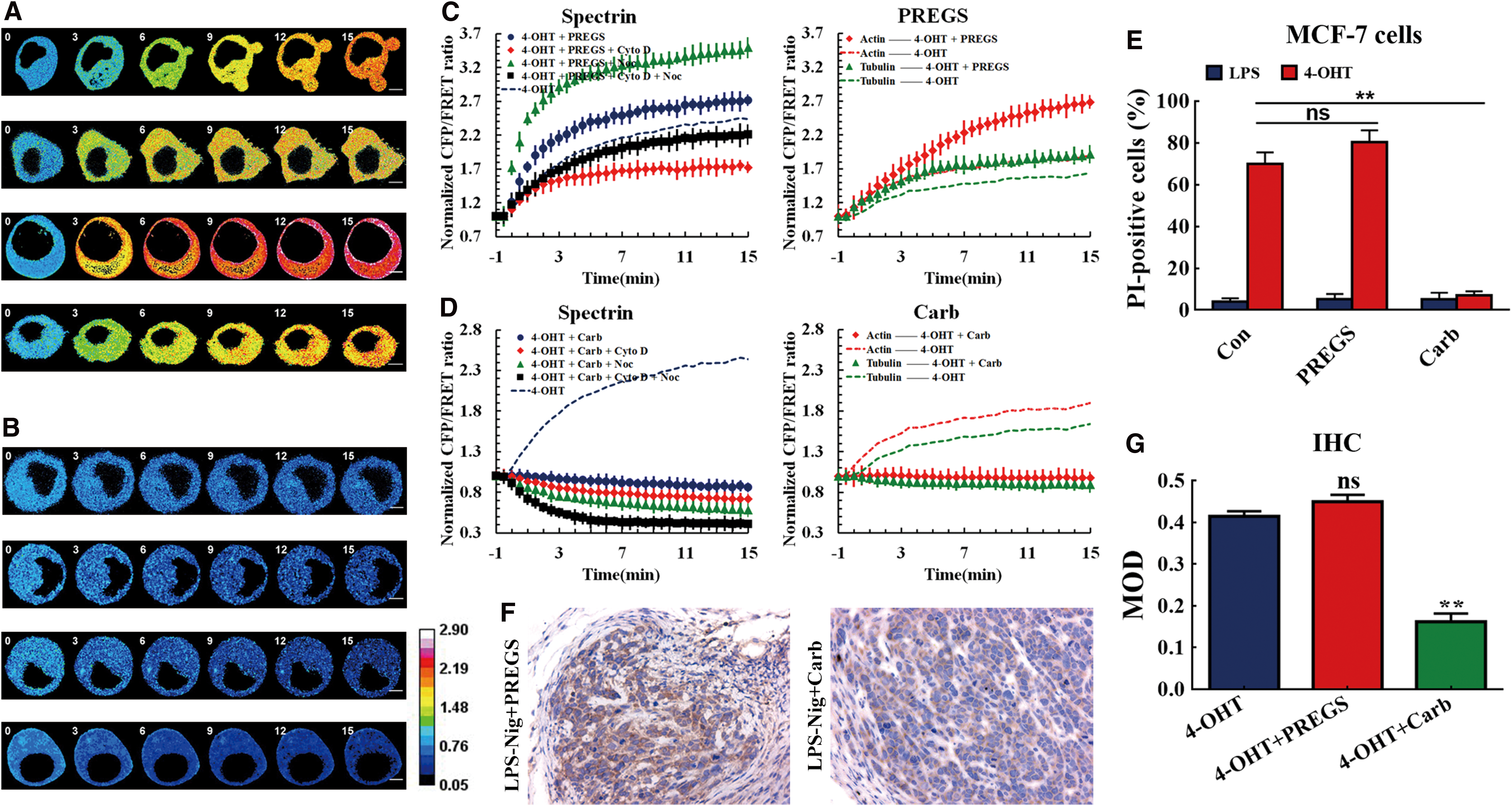
Next, we attempted to investigate the role of calcium channel inhibitor carboplatin (Carb) in cytoskeletal structural tension and pyroptosis. Carb triggered a sustained decrease in spectrin tension to below the control level, and suppressed cell swelling and blebbing (Fig. 7B). The depolymerization of MFs and MTs induced a relative decrease in the spectrin tension and pyroptosis rate compared to the pyroptosis model. Calcium depletion eliminated MF and MT forces in pyroptotic cells (Fig. 7D, E). IHC staining results also suggest that Carb but not PREGS treatment induced a significant reduction in caspase-1 activity in the in vivo MCF-7 xenograft model (Fig. 7F, G). The data suggest that calcium channels and their related calcium signals are general regulators of intracellular tension, coordinating cell shape during pyroptosis via structure tension and crosstalk with mechanical signals, under physiological and pathological circumstances.
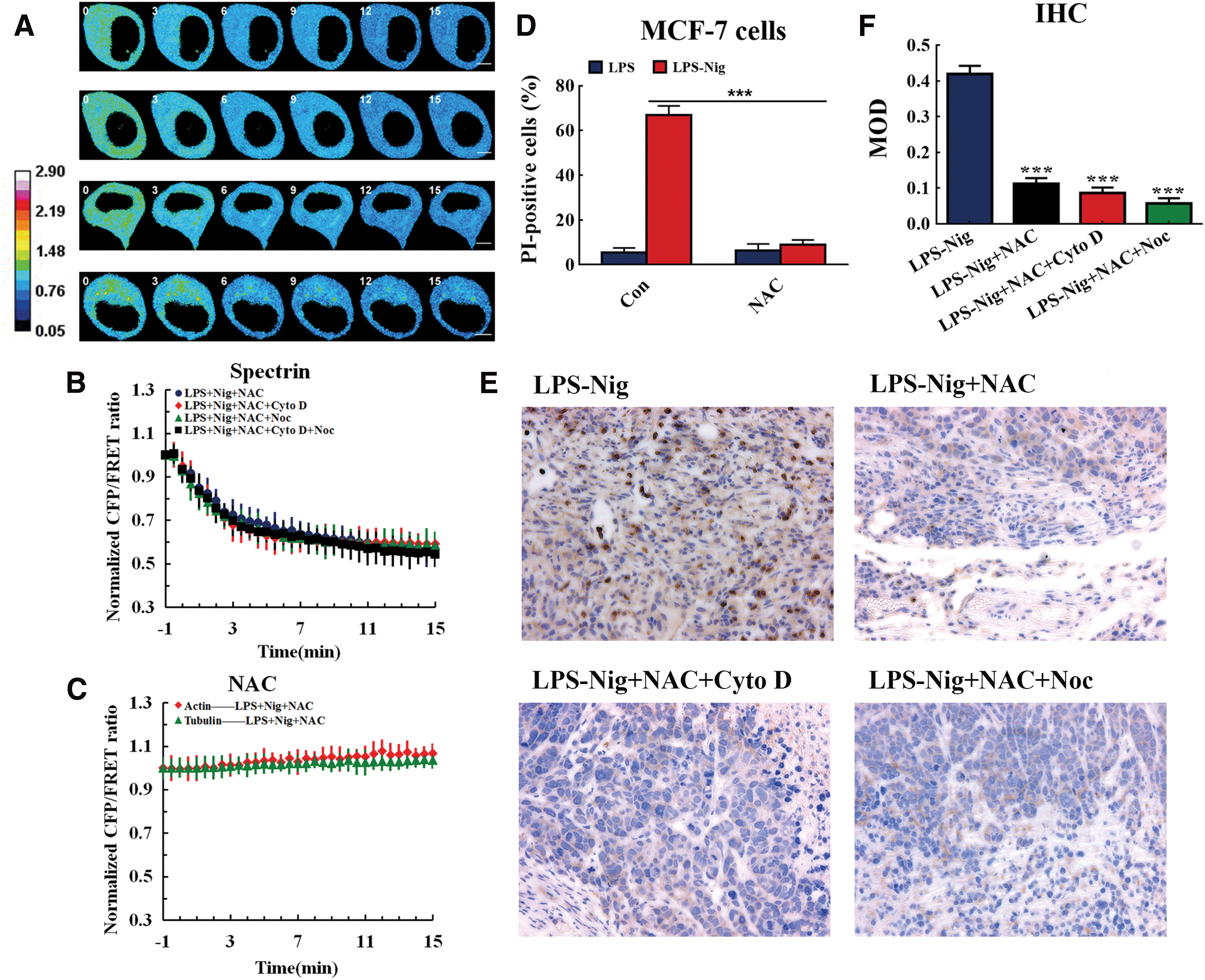
ROS elimination by antioxidants can inhibit cytoskeleton tension in pyroptosis
Antioxidants can restrain the damage of excessive ROS through maintaining intracellular glutathione redox balance and improving the redox buffering properties of cells. To explore the role of ROS elimination in pyroptosis, we treated the MCF-7 cells with N-acetyl-
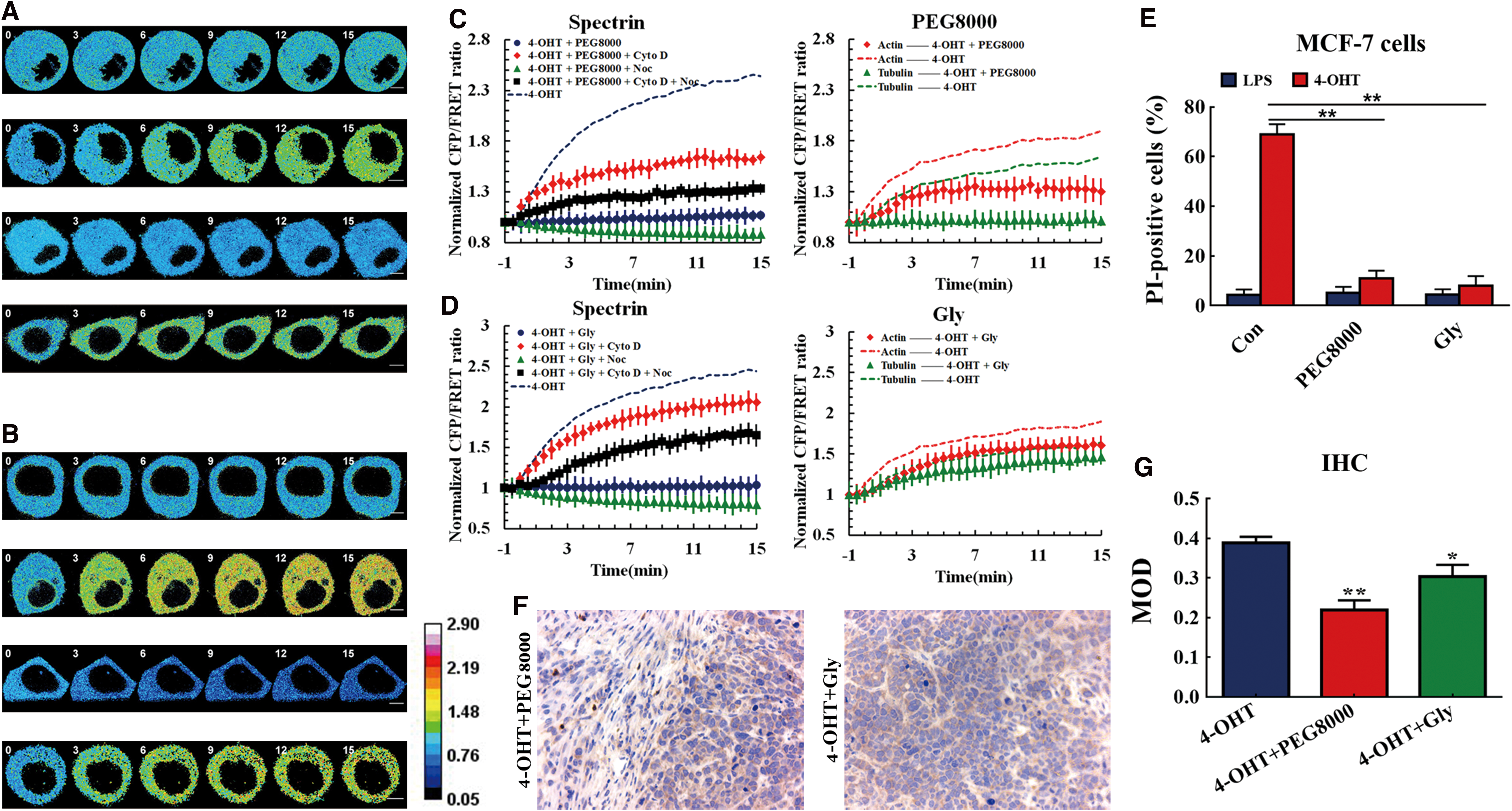
Formation of membrane pores and nonselective ion influx modify spectrin tension via MF and MT forces but not osmotic pressure
Membrane pores can form during pyroptosis, facilitating the uncontrollable flow of ions, including Ca2+, K+, and Cl− (11), which can influence structure tensions in the spectrin, MF, and MT. Membrane pore blocker polyethylene glycol 8000 (PEG8000) was used to block the pyroptosis-induced membrane pores. In comparison with the pyroptosis model, cellular swelling and blebbing were prevented, and spectrin tension remained stable, with MF and MT forces maintained at lower levels (Fig. 9A, C). These findings are consistent with published data that confirmed that large-sized PEG could block membrane pores (54). Protection conferred by PEG8000 also suppressed cellular pyroptosis (Fig. 9E). The depolymerization of MFs induced a relative increase in spectrin, while MT depolymerization reduced spectrin tension. The data suggest that MF and MT forces acted antagonistically in spectrin tension (Fig. 9C, D). A similar phenomenon was observed when the ion influx inhibitor glycine (47) was used (Fig. 9B, D). Rein in ion liquidity in pyroptosis progression improved a partly antagonistic role of MF and MT forces in spectrin tension. The impact of PEG8000 and glycine is consistent with inhibition of pyroptosis (Fig. 9E) (38). Similarily, PEG8000 and glycine (Gly) were heavily involved in the suppression of caspase-1 activity in the in vivo MCF-7 xenograft model (Fig. 9F, G). Collectively, these data indicate that the formation of membrane pores serves to control intracellular cytoskeletal structural tension during pyroptosis, and that spectrin tension modified via MF and MT forces might be not associated with osmotic pressure.
Depletion of caspase-1 triggers pyroptosis inhibition and alters cytoskeletal structural tension
Pyroptosis is programmed cell death via a caspase-1-dependent pathway, which involves cytoskeleton rearrangement and membrane pore formation. To explore the role of caspase-1 activation in pyroptosis, small interfering RNA of caspase-1 was transferred to stable transfection cells with tension sensors. The impairment of p20 production in caspase-1−/− cells directly correlated with caspase-1 inactivation and pyroptosis inhibition (Fig. 10B, E, and Supplementary Fig. S4). Restrained caspase-1 activity was accompanied by a reduction in spectrin tension (Fig. 10A, C). MF and MT depolymerization resulted in a relative decrease and increase in spectrin tension, respectively, indicating that MF and MT forces act antagonistically in spectrin tension (Fig. 10A, C). Caspase-1 inactivation resulted in the reduction of MF and MT tension (Fig. 10D), suggesting that the low spectrin tension was elicited by the reduction of MF and MT forces in caspase-1−/− cells. Together, these data show that intracellular structural strains can be modified by chemical caspase-1 signals, and the signal pathway can regulate changes in cell shape and volume via the interaction of intracellular strains or crosstalk between chemical and tension signals during pyroptosis.

Discussion
Along with cytoskeleton remodeling, subcellular tension in the cytoskeleton also leads to shifts in cellular shape in the form of swelling and blebbing during pyroptosis. Spectrin is anchored to the cell membrane and controls plasma membrane protrusions and shrinkage in pyroptosis. During pyroptosis, spectrin, as a force sensor, supports membrane stabilization and acts as an interchange adapter for cytoskeleton remodeling and tension transmission. Using FRET-based tension sensors, dependent on the angular orientation of the donor and acceptor (56), we measured the spectrin tension in living cells. Variation in constitutive spectrin tension originates from interactions of MF and MT strains. Remodeling of the MF and MT forces also drives spectrin tension changes and cellular deformation, such as pyroptotic swells and blebs.
Three forces, osmotic pressure, MF, and MT forces in live cells, serve to control intracellular tensions (24, 37, 56). Spectrin tension can be modified via their interaction. These steerable tensions can be activated in reaction to cell stress. Upregulation or downregulation of osmotic pressure can lead to alterations in MF and MT forces. The intracellular cytoskeletal structural tension can compete with outward or inward osmotic pressure, and is involved in the stabilization of cellular shape and volume, even RVD phenomena (6). The presented study discerned that interactions between MF and MT tensions can be synergistic or antagonistic in response to osmotic pressure. Apart from intracellular force magnitudes, their vectors are crucial factors in subcellular tension activities. The vector of intracellular tension can be analyzed by the comparison between these mechanical forces and the elimination of certain cytoskeletal structural tensions induced by their depolymerizing agents, providing an efficient way to decipher the vector puzzle (Table 1).
Interaction Between Osmotic Pressure, Microfilament, and Microtubule-Dependent Tensions
↑ Indicates the increment; ↓ indicates the reduction; = indicates the stabilization; → shows the inward direction; ← presents the outward direction.
MF, microfilament; MT, microtubule; RVD, regulatory volume decrease.
The occurrence of pyroptosis is accompanied by membrane swelling and blebbing, and the enhanced tension within spectrin relies mostly on MF and MT forces. Spectrin tension depends on MF strain, which can also be reversed by MT strain during pyroptosis. The fluctuating tension along MF and MT can be transduced antagonistically across the plasma membrane to enhance spectrin tension and induce pushing or pulling tension. Reduction of inward MF force can stimulate more blebbing, while vanishing outward MT strain led to reduced bleb formation and enhanced spectrin tension in pyroptosis. Bleb formation might be associated indirectly with spectrin tension during pyroptosis. In this case, spectrin tension, derived from the inward contractile tension of MF, contributes significantly to cell volume shrinkage and inhibition of cell deformation, resulting in protection during pyroptosis.
Cytoskeletal structural tension depends on intracellular cytoskeleton architecture (37, 56). Depolymerization of the cytoskeleton structure can result in cellular deformation, which indicates that cytoskeleton structure is crucial to alterations in cell shape (9). However, testing of MF and MT stabilizers, blocking filament elongation via capping the growing barbed end of F-actin and microtubulin, provides additional information. Inhibition of MF and MT rearrangement does not affect the occurrence of pyroptosis. Stabilization of MF and MT structures exerts little effect on intracellular structure tension and deformation of pyroptotic cells. Notably, depolymerization of MT led to inhibition of pyroptosis and contributed to the relief of subcellular strain (Fig. 1F), which was further supported by in vivo experiments (Fig. 5A, C). It should be admitted that rearrangement of MF and MT can contribute but is not a necessary condition for the alteration of intracellular tension (21). The present study further clarifies that structural forces, which are not cytoskeletal, control alterations in cell conformation and shape that are relevant to pyroptosis. The data also provide new insights into the role of pyroptosis inducers that are closely associated with intracellular mechanical forces, which is interesting in terms of targeting decreased breast tumor burden in cancer biology. This study suggests a potential functional basis for further cytoskeleton tension study with a view to developing new targets for antitumor drugs, leading to changes in the paradigms of anticancer drug discovery toward intracellular tension-targeted therapeutics.
Caspase-1 activation, pore formation, and nonselective ion flux are features of pyroptosis (18), and surface tension was found to depend on these properties during its progression. Activation of voltage-activated calcium channels aggravates structural spectrin, MF, and MT tensions. Higher cytoskeletal structural tension is associated with aggressive swelling and blebbing in pyroptosis. Meanwhile, the elimination of structure strain by calcium channel inhibitors is linked to depression in swelling and blebbing. The data suggest that calcium ion influx controls adjustments in intracellular cytoskeletal structural tension and the progression of pyroptosis. Meanwhile, ROS accumulation also plays a vital role in cytoskeletal tension in pyroptotic cells. ROS production is thought to be associated with caspase-1 activation and cytoskeleton alteration (36, 50). Not surprisingly, ROS accumulation may result in cell deformation through cytoskeleton force activity, including spectrin tension. Furthermore, pore formation could induce nonselective ion flux and regulate ion distribution on both sides of the plasma membrane in pyroptotic cells. According to our results, ion signals, rather than a loss of cellular ionic equilibrium, contribute to augment structural strains and the progress of pyroptosis. Likewise, the loss of caspase-1-dependent lysis by caspase-1 knockdown resulted in a decrease in the steerable tension forces during pyroptosis. Caspase-1 activation is a vital effector in pyroptosis and controls structural stress activities and adjustments in pyroptosis.
Most likely, the crosstalk between chemical and tension signals makes a “decision” for the progression of pyroptosis. Furthermore, pyroptosis can be attenuated by disrupting MT and its dependent force (Fig. 1F). The evidence that depolymerization of microtubulin reversed pyroptosis in the in vivo MCF-7 xenograft model further supported our data (Figs. 1F, 5A, and C). Unexpectedly, the results indicated that MF tension plays a much more important role in pyroptosis inhibition in tissues than in cultured cells, probably due to contact inhibition and adhesion restriction in in vivo tissues (35), outer tension of MT in tissues might be restricted or limited (Figs. 1F, 5A, and C). The data presented illustrate that relieving cytoskeletal structural tension can result in pyroptosis remission. Therefore, structural strain may also be characteristic of pyroptosis progression, which is closely associated with the magnitude and direction of subcellular forces.
In conclusion, the present study demonstrates that spectrin tension can be produced and modulated by MF and MT tension, the vectors of which are also vital variables. The MF and MT forces act in a push and pull-type manner to modulate the plasma membrane via motor molecule activities. MT forces are the primary tension contributors to bleb formation, while spectrin tension elicited by MF strain modifies the membrane swell in pyroptosis. Pyroptosis-based cytoskeletal tension may be an important and clinically relevant anticancer consideration. The formation of membrane pores or nonselective ion flux in pyroptosis can control the cytoskeletal structural tension via ion signals, but not osmotic pressure. Chemical signals, ROS generation, or enzyme activity can also regulate structure tension in pyroptosis. The results further indicate that subcellular structural strains act as a vector and can interact with chemical signals via their magnitude and direction, driving cellular deformation. Similar to chemical and electrical cellular activity, intracellular cytoskeletal structural tensions as signals or effectors in the live cell are essential components of metabolic life. This in-depth study should enrich medical research and shed more light on the mysteries of living cells.
Materials and Methods
Reagents and antibodies
Nigericin (Nig) was purchased from Santa Cruz (CA). Ultrapure LPS, 4-OHT, cycloheximide, and JK were purchased from Aladdin (Shanghai, China). Cyto D, TOX, PREGS, Noc, and NAC were provided by Sigma-Aldrich (St. Louis, MO). Carb was acquired from Merck Millipore (Darmstadt, Germany). Antibodies were purchased from commercial sources: rabbit anti-spectrin-β antibody (20856R; Bioss, Woburn), rabbit anti-Caspase-1 antibody (BM4291; Boster, Wuhan, China), rabbit anti-β-actin antibody (4970P; Cell Signaling, Temecula), and mouse anti-α-tubulin antibody (BM1452; Boster, Wuhan, China).
Animals
A total of 40 C57BL/6 female mice (20–22 g, six weeks old) and 80 female nude mice (16–18 g, six weeks old) were obtained from the Model Animal Research Center of Nanjing University of Chinese Medicine and housed in pathogen-free mouse colonies. Principles of laboratory animal care were followed, and all procedures were conducted according to the guidelines established by the National Institutes of Health. This study was approved by the Research Animal Care Committee of Nanjing University of Chinese Medicine (Nanjing, China).
Tumor inoculation in nude mice
To establish the human breast cancer (MCF-7) xenograft model, 1 × 107 MCF-7 cells suspended in 100 μL phosphate-buffered saline (PBS) were implanted into the subcutaneous tissues in the right armpits of nude mice. Four weeks after tumor inoculation, the mice were intravenously coinjected with LPS plus vehicle (200 μL of saline) and six mice each were randomly assigned to the following treatment groups: (i) untreated control, (ii) LPS-Nig, (iii) LPS-Nig + Cyto D, (iv) LPS-Nig + Noc, (v) LPS-Nig + JK, (vi) LPS-Nig + TOX, (vii) 4-OHT + PREGS, (viii) 4-OHT + Carb, (ix) 4-OHT + PEG8000, (x) 4-OHT + Gly, (xi) LPS-Nig + NAC, (xii) LPS-Nig + NAC + Cyto D, and (xiii) LPS-Nig + NAC + Noc. Two hours later the mice were euthanized and the primary tumors were excised.
Control, LPS-Nig, LPS-Nig + Cyto D, and LPS-Nig + Noc treatments of mice
LPS-Nig, LPS-Nig + Cyto D, and LPS-Nig + Noc were intraperitoneally coinjected with LPS plus vehicle (200 μL of saline) into C57BL/6 mice. Two hours later, the control and LPS-Nig, LPS-Nig + Cyto D, and LPS-Nig + Noc-treated mice were sacrificed, and the number four inguinal mammary glands of mice were collected.
IHC staining and analysis
Number four inguinal mammary glands and 13 MCF-7 xenograft models of mice were fixed in 10% formalin overnight at 4°C. The glands were then dehydrated in a graded ethanol series followed by xylene and embedded in paraffin. Four-micrometer sections were prepared and stained for caspase-1 expression (Leica CM3050 S; Leica Biosystems). For caspase-1 quantification, 10 images were captured per slide at 40 × magnification using a Leica DMi8 microscope. Semiquantitative analysis of the IHC images was conducted using a Image-Pro Plus Version 6.0 (Media Cybermetics, MD), from which integral optical density (IOD) and area data were collected. Then, the average optical density was calculated as IOD/area, which represented the staining intensity.
Design of tension sensors
The sensors were structured using the NovoRec PCR Seamless Cloning Kit and restriction enzyme cloning techniques, in accordance with previous reports (16). We constructed fluorescent sensors with circularly permutated cpVenus and cpCerulean (cpVenus–7aa–cpCerulean [cpstFRET]). The circularly permutated sstFRET (cpstFRET) sensors were created by substituting Venus and Cerulean with cpVenus and cpCerulean, separately. We cloned α-tubulin and β-tubulin and incorporated them at each side of cpstFRET to create pCMV-tubulinα-cpstFRET-tubulinβ (TcpT). In addition, we created pCMV-tubulinα-cpstFRET (cpT) as the control. pCMV-spectrin-cpstFRET (ScpS, the cpstFRET was inserted at amino acid position 1201 in nonerythroid spectrin), pCMV-spectrin-cpstFRET (cpS), pCMV-actin-cpstFRET-actin (AcpA), and pCMV-actin-cpstFRET (cpA) were fabricated according to a similar protocol (23).
Cell culture and transfection with tension sensors
MCF-7 (human breast metastatic adenocarcinoma) cells (HTB 22; ATCC, Manassas) were cultured on fibronectin-coated coverslips in Dulbecco's modified Eagle's medium (319-005-CL; Wisent, Montreal, Canada) containing 10% fetal bovine serum (086-150; Wisent) and a 1% mixture of penicillin and streptomycin (Gibco, Invitrogen). First, we examined the morphology and growth characteristics of the cells. Then, they were routinely tested for mycoplasma contamination using the Mycoplasma Plus PCR Primer Set (Agilent, Santa Clara, CA) and were found to be negative.
We used the E.Z.N.A Endo-free plasmid DNA Mini Kit II (OMEGA, Tarzana) to extract single colony plasmids, according to the manufacturer's instructions. Cells were transfected for 24 h before the experiments with tension sensor plasmids using FuGENE 6 Transfection Reagent (Roche Diagnostics, Basel, Switzerland), following the manufacturer's specifications. The transfection efficiency was ∼60%. The transfected MCF-7 cells were sorted using a Moflo XDP cell sorter with Summit 5.3 software (Beckma Coulter). Samples were analyzed by the excitation lasers and emission filters: cyan-458-nm laser and 447/60 bandpass filter; yellow-514-nm laser and 580/23 bandpass filter. The cells were sorted when the cyan and yellow emission wavelengths were detected at the same time. The sorted MCF-7 cells were diluted into single cells and then seeded on 96-well plates. In all experiments, the sorted single-cell lines were trypsinized, and then, the cells were incubated in cell culture media for 24–36 h.
CpstFRET analysis
The dipole angle between donor/CFP and acceptor/YFP determined the effectiveness of FRET (23). Cells were imaged on a Leica confocal microscope SP5 equipped with a × 63 oil-immersion objective. The donor and acceptor were tested by 458 and 514 nm argon lasers, respectively. CFP/FRET ratios were calculated using the equation 1/R = Cerulean donor/Venus acceptor.
Western blot
The extracted total proteins were separated via sodium dodecyl sulfate/polyacrylamide gel electrophoresis. Then, we transferred the protein band to the nitrocellulose membrane via wet electrotransfer. The primary antibodies were antibodies against caspase-1, actin-β, or tubulin-α. After blotting, the protein side of the membrane was exposed to X-ray film.
Determination of MCF-7 pyroptosis
Pyroptotic cells were assessed by the double-positive staining of activated caspase-1 and PI in MCF-7. The activated caspase-1 was detected by a caspase-1 activity assay kit (Beyotime Biotechnology; C1101), while PI was detected using a cell cycle and apoptosis analysis kit (Beyotime Biotechnology; C1052), according to the manufacturer's instructions. Stained cells were then analyzed by flow cytometry (BD FACSCalibur; Becton, Dickinson, and Company).
Bleb cell counts and measure of cell surface area
The average percentage of blebbing cells was calculated by comparing the number of blebbing cells to the total number of cells. The FRET three-dimensional images were analyzed using ImageJ software (U.S. National Institutes of Health) for quantification of the cell surface area. The length (longest, a), width (shortest, b), and altitude (height, c) radii of each cell were measured by fitting the cell outline to an optimized ellipse shape. The cell surface area was calculated using the equation (4/3) × π × (ab + bc + ca). For quantification, three images were taken and analyzed, and the calculated results were averaged.
Immunofluorescence analyses
In immunocytochemical studies, MCF7 cells were washed twice and fixed in newly prepared 4% paraformaldehyde solution for 20 min. The membrane was permeabilized in 0.1% Triton X-100 at 4°C and blocked with 3% bovine serum albumin/PBS for 30 min. For fluorescence labeling, cells were incubated with the primary antibody. After incubation overnight at 4°C, the cells were washed and incubated with the secondary antibodies in the dark. 4′,6-Diamidino-2-phenylindole was used to stain the nuclei. Labeled samples were examined by immunofluorescence using a confocal microscope (TCS SP5; Leica).
Statistical analysis
The ratio of YFP/CFP was calculated using the ImageJ software package. The FRET ratio in each subcellular region was measured for each cell and then averaged over multiple cells. The images were pseudocolored using ImageJ's 16-color map. All data are expressed as mean ± standard error of the mean. We applied one-way analysis of variance for single-factor sample comparisons and a least significant difference test for comparisons between any two means. Each experiment was repeated at least three times, >10 cells were imaged, and each condition was analyzed. The R of MCF-7 cells is presented as the pixel count distribution from >10 MCF-7 cells.
Footnotes
Acknowledgments
We thank Frederick Sachs and Fanjie Meng (University at Buffalo) for providing the plasmids encoding spectrin-cpstFRET (ScpS) and spectrin-cpstFRET (cpS). This work was supported by grants from the National Natural Science Foundation of China (81573409), the Natural Science Foundation of Jiangsu Province (BK20161574), and the Project of Priority Academic Program Development of Jiangsu Higher Education Institutions (Integration of Traditional Chinese and Western Medicine).
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.